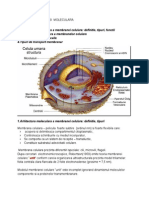
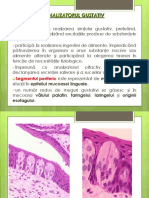
S-ar putea să vă placă și
- Citoscheletul PDFDocument41 paginiCitoscheletul PDFOzana CopyShopÎncă nu există evaluări
- Transportul MembranarDocument8 paginiTransportul MembranarCojocariu ClaudiuÎncă nu există evaluări
- Capitolul I CELULA SI TESUTURIDocument14 paginiCapitolul I CELULA SI TESUTURIVioleta DiaconuÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- C07 CitoscheletulDocument28 paginiC07 Citoscheletulpanterablo100% (1)
- Mecanismele Genetice Ale Diversitatii ImunoglobulinelorDocument7 paginiMecanismele Genetice Ale Diversitatii ImunoglobulinelorGumus MedelaineÎncă nu există evaluări
- Biocel Sub Rezolvate 2009Document25 paginiBiocel Sub Rezolvate 2009ariadna89100% (2)
- Biocel 1Document6 paginiBiocel 1Elena IscuÎncă nu există evaluări
- Membrana CelularaDocument6 paginiMembrana Celularaveronica.99Încă nu există evaluări
- CitoscheletulDocument6 paginiCitoscheletulJessica Smith100% (1)
- Examen Biologie CelularaDocument8 paginiExamen Biologie CelularaElena IscuÎncă nu există evaluări
- Biochimie Teste ClalculatorDocument15 paginiBiochimie Teste ClalculatorVirgiliuMateiÎncă nu există evaluări
- Histologie 1 Curs 3) Tesuturi ConjunctiveDocument7 paginiHistologie 1 Curs 3) Tesuturi ConjunctiveMariaMarya100% (2)
- Citoscheletul MembranarDocument1 paginăCitoscheletul MembranarCristina PopÎncă nu există evaluări
- Imunitatea InnascutaDocument30 paginiImunitatea InnascutaKfa Dkh100% (1)
- Biofizică MedicalăDocument18 paginiBiofizică MedicalăGeorgiana HarjaÎncă nu există evaluări
- LP - OSMOZA Cu Calcul Pres OsmoticaDocument7 paginiLP - OSMOZA Cu Calcul Pres OsmoticaDarth Raz-WanÎncă nu există evaluări
- Tesut EpitelialDocument16 paginiTesut EpitelialLorelai_20Încă nu există evaluări
- Curs 3 Aparatul GolgiDocument4 paginiCurs 3 Aparatul GolgiOana Koppandi100% (1)
- MitozaDocument3 paginiMitozaAlexandra MiocÎncă nu există evaluări
- Biofizica PT EleviDocument91 paginiBiofizica PT EleviCojocaru ConstantinÎncă nu există evaluări
- Matrice ExtracelularaDocument27 paginiMatrice ExtracelularaOana PatriciaÎncă nu există evaluări
- Fiziologia Membranei CelulareDocument29 paginiFiziologia Membranei CelulareCondrea100% (1)
- Curs Proteine Structura ProteomicaDocument96 paginiCurs Proteine Structura ProteomicaIoana BandilaÎncă nu există evaluări
- ANALIZATORII. 2pptDocument47 paginiANALIZATORII. 2pptCasandra TudoracheÎncă nu există evaluări
- Rolul Ficatului În Biochimia LipidelorDocument6 paginiRolul Ficatului În Biochimia Lipidelorlaurisca937660Încă nu există evaluări
- Fiziologie Generala C.cristescuDocument20 paginiFiziologie Generala C.cristescudamaskin9480% (5)
- Lp4-Varietati de Tesut ConjunctivDocument3 paginiLp4-Varietati de Tesut ConjunctivCrinablue100% (1)
- CitoscheletulDocument1 paginăCitoscheletulAna Maria Cristina MateiÎncă nu există evaluări
- Cursuri Fizio 1Document416 paginiCursuri Fizio 1Florea RaduÎncă nu există evaluări
- Biochimia Senescentei Stresul OxidativDocument42 paginiBiochimia Senescentei Stresul OxidativGabriela TomaÎncă nu există evaluări
- Ţesutul Epitelial Glandular (Secretor)Document36 paginiŢesutul Epitelial Glandular (Secretor)Andra AgheorghieseiÎncă nu există evaluări
- Structura Primara A Acizilor NucleiciDocument6 paginiStructura Primara A Acizilor NucleiciValérie IordatiÎncă nu există evaluări
- Teste Grile Fiziologie ColorateDocument283 paginiTeste Grile Fiziologie Coloratealinusha_7100% (1)
- AQUAPORINEDocument26 paginiAQUAPORINEProca IonÎncă nu există evaluări
- 3.fiziologia LeucocitelorDocument28 pagini3.fiziologia LeucocitelorSchuster Stephan100% (2)
- Referat docxVIOLETADocument10 paginiReferat docxVIOLETAtuturici liviuÎncă nu există evaluări
- Manual HistologieDocument94 paginiManual HistologieGabrielaÎncă nu există evaluări
- MicrotubuliiDocument2 paginiMicrotubuliiDavidRaduÎncă nu există evaluări
- C04 Membrana Celulara JonctiuniDocument46 paginiC04 Membrana Celulara JonctiunipanterabloÎncă nu există evaluări
- ATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialDocument8 paginiATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialPascu Alexandru0% (1)
- Morfologia - Anatomia Plantelor - AH 2 - Tesuturile PDFDocument61 paginiMorfologia - Anatomia Plantelor - AH 2 - Tesuturile PDFMinodora Isar100% (2)
- Curs VizualDocument31 paginiCurs VizualLarisa P. OanceaÎncă nu există evaluări
- 6 Fosforilare Oxidativa, Sinteza Proteica, Trafic de MembraneDocument28 pagini6 Fosforilare Oxidativa, Sinteza Proteica, Trafic de MembraneGreta IulianaÎncă nu există evaluări
- Boli Genetice MitocondrialeDocument21 paginiBoli Genetice Mitocondrialeiulica mybsÎncă nu există evaluări
- Ciclul CelularDocument19 paginiCiclul CelularElena-Catalina Popa100% (1)
- Structura Osoasa Si OsteogenezaDocument58 paginiStructura Osoasa Si OsteogenezaV1ctor1aÎncă nu există evaluări
- Carte LP 2007 Biologie Celulara-1Document85 paginiCarte LP 2007 Biologie Celulara-1Alina Madalina50% (2)
- Fiziologie - Curs 5 Neuronul + MiocardulDocument8 paginiFiziologie - Curs 5 Neuronul + Miocardulionut_lupul100% (1)
- BiocelDocument40 paginiBiocelAdelina MusaÎncă nu există evaluări
- BiocelDocument41 paginiBiocelremus_stoica23Încă nu există evaluări
- Curs 4 - CitoscheletDocument46 paginiCurs 4 - CitoscheletAdrian UrsuÎncă nu există evaluări
- Curs Nr. 8'Document5 paginiCurs Nr. 8'Co BbÎncă nu există evaluări
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- Curs 6 - Matricea Citoplasmatica (Citoschelet)Document14 paginiCurs 6 - Matricea Citoplasmatica (Citoschelet)Alexandra MincaÎncă nu există evaluări
- Ciclu CardiacDocument17 paginiCiclu CardiacCiprian JurjeÎncă nu există evaluări
- Biologie CelularaDocument7 paginiBiologie CelularaBogdan CirjanÎncă nu există evaluări
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Tesutul MuscularDocument4 paginiTesutul MuscularAriela Săvuţ100% (1)
- Muschi NetedDocument6 paginiMuschi Netediuuu3Încă nu există evaluări
- Grile - Diabet ZaharatDocument26 paginiGrile - Diabet ZaharatAlina CalmucÎncă nu există evaluări
- Rezumat Iordache AlinDocument29 paginiRezumat Iordache AlinComan IoanaÎncă nu există evaluări
- Starea de Bine A FatuluiDocument49 paginiStarea de Bine A FatuluiAlina CalmucÎncă nu există evaluări
- Precizari StagiuDocument1 paginăPrecizari Stagiur_zxcvbnmÎncă nu există evaluări
- Violeta B - Imunologie Curs NouDocument80 paginiVioleta B - Imunologie Curs NouAlina CalmucÎncă nu există evaluări
- Obezitate + Diabet (10 Nov-Prof. Fica)Document48 paginiObezitate + Diabet (10 Nov-Prof. Fica)Alina CalmucÎncă nu există evaluări
- CozonacDocument4 paginiCozonacAlina CalmucÎncă nu există evaluări
- Mutatii GeneticeDocument24 paginiMutatii GeneticeTeo BăjeuÎncă nu există evaluări
- Precizari StagiuDocument1 paginăPrecizari Stagiur_zxcvbnmÎncă nu există evaluări
- Grile GinecologieDocument9 paginiGrile GinecologieCURT FLORIANÎncă nu există evaluări
- Prezentiatia Pelviana, Transversa Si Prezentatiile Deflectate - Prezentatii, Pozitie, Varietati de PozitieDocument22 paginiPrezentiatia Pelviana, Transversa Si Prezentatiile Deflectate - Prezentatii, Pozitie, Varietati de PozitiePal OareÎncă nu există evaluări
- CozonacDocument4 paginiCozonacAlina CalmucÎncă nu există evaluări
- Semiologie Grile TipDocument39 paginiSemiologie Grile TipKaybukan Constanta100% (3)
- Rolul FormatoruluiDocument79 paginiRolul FormatoruluimateimaraÎncă nu există evaluări
- Carte PaiDocument144 paginiCarte PaiAlina CalmucÎncă nu există evaluări
- Grile Antropologie UMF Carol Davila 2012Document16 paginiGrile Antropologie UMF Carol Davila 2012Rares Deculescu100% (1)
- Rezumat ToldisanDocument33 paginiRezumat ToldisanAlina CalmucÎncă nu există evaluări
- Albert Einstein - Despre Dumnezeu Si StiintaDocument4 paginiAlbert Einstein - Despre Dumnezeu Si StiintacalinsatÎncă nu există evaluări
- 121 Fisa de LucruDocument1 pagină121 Fisa de LucruAlina CalmucÎncă nu există evaluări
- Cap4 - Tehnici de Procesare A Imaginilor Bio Medic AleDocument32 paginiCap4 - Tehnici de Procesare A Imaginilor Bio Medic AleMagor Kozma-vigÎncă nu există evaluări
- Sindromul Wolf - HirschhornDocument2 paginiSindromul Wolf - HirschhornPetru OnicăÎncă nu există evaluări
- Mecanisme de CopingDocument8 paginiMecanisme de CopingCristian Vasile BotaÎncă nu există evaluări
- Allan Carr - in Sfarsit NefumatorDocument183 paginiAllan Carr - in Sfarsit NefumatorDragosÎncă nu există evaluări
- Subiecte Examen Oral FiziologieDocument2 paginiSubiecte Examen Oral FiziologieCătălin NechitaÎncă nu există evaluări
- Fisa de Caracterizare PsihopedagogicaDocument2 paginiFisa de Caracterizare PsihopedagogicaAlina Calmuc100% (1)
- Cordul CursDocument11 paginiCordul CursAlex AliÎncă nu există evaluări
- ExcitabilitateaDocument27 paginiExcitabilitateastrengara100% (2)
- Jester Si Mos CraciunDocument16 paginiJester Si Mos CraciunLidia Lorena NituÎncă nu există evaluări
- Subiecteanato LazarDocument5 paginiSubiecteanato LazarAlina CalmucÎncă nu există evaluări