S-ar putea să vă placă și
- Biofizica Contracţiei MusculareDocument6 paginiBiofizica Contracţiei MuscularenoruletÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument4 paginiBiofizica Contracţiei Muscularechamps2009Încă nu există evaluări
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Biofizica Contractiei MusculareDocument6 paginiBiofizica Contractiei MusculareAndrone CameliaÎncă nu există evaluări
- Biofizica Contractiei Musculare Sistemul Muscular PDFDocument1 paginăBiofizica Contractiei Musculare Sistemul Muscular PDFandamuresan3Încă nu există evaluări
- Contracția Musculara BF FIMDocument16 paginiContracția Musculara BF FIMVlad ȘtefanÎncă nu există evaluări
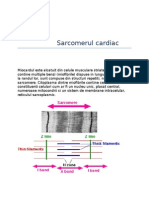
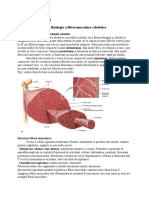
- Sarcomerul CardiacDocument17 paginiSarcomerul CardiacCiobotaru AlexandraÎncă nu există evaluări
- MuscularDocument9 paginiMuscularpufdepapadieÎncă nu există evaluări
- Fiziologia Contractiei MusculareDocument4 paginiFiziologia Contractiei MusculareIoana irimiaÎncă nu există evaluări
- Tesutul MuscularDocument4 paginiTesutul MuscularAriela Săvuţ100% (1)
- 06 Contractia Musculara 2017-12-04Document11 pagini06 Contractia Musculara 2017-12-04costica16100% (1)
- Morfologia Muschilor Striati 1Document10 paginiMorfologia Muschilor Striati 1AlexColibaÎncă nu există evaluări
- Structura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDocument2 paginiStructura Si Proprietetile Fiziologice Ale Fibrelor Muschilor StriatiDaniela IstratiÎncă nu există evaluări
- Fiziologia Muschilor StriatiDocument8 paginiFiziologia Muschilor Striatinigor1297Încă nu există evaluări
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- FIziologia Aparatului NeuromuscularDocument14 paginiFIziologia Aparatului NeuromuscularLoredana Boghez100% (1)
- Fiziologia MuşchilorDocument74 paginiFiziologia MuşchilorXiusa TurcanÎncă nu există evaluări
- Fiziologia Muschiului CardiacDocument14 paginiFiziologia Muschiului Cardiacthorscribd100% (7)
- Contractia MuscularaDocument12 paginiContractia MuscularaChriss ChristinaÎncă nu există evaluări
- Ţesut MuscularDocument12 paginiŢesut MuscularStan DianaÎncă nu există evaluări
- Contractia MuscularaDocument4 paginiContractia MuscularaTudor OlteanÎncă nu există evaluări
- Referat FiziollogieDocument7 paginiReferat FiziollogiedarlaloredanaÎncă nu există evaluări
- Curs 5 Muschiul Striat NetedDocument6 paginiCurs 5 Muschiul Striat Netedp3tru_mzqÎncă nu există evaluări
- ElectromiogramaDocument6 paginiElectromiogramaAna Maria100% (1)
- Biofizica CursuriDocument67 paginiBiofizica CursuriMihnea TurcuÎncă nu există evaluări
- Biochima MuschilorDocument7 paginiBiochima MuschilorstikssÎncă nu există evaluări
- Cursul 4 FizioDocument5 paginiCursul 4 Fiziodanyela0808Încă nu există evaluări
- Curs - Muschi StriatDocument40 paginiCurs - Muschi StriatDiana Didi Dia100% (1)
- Cursul IV, Note de Curs Neuro-MuscularDocument16 paginiCursul IV, Note de Curs Neuro-MuscularYusef Ceva100% (1)
- Fiziologia MușchilorDocument12 paginiFiziologia MușchilorVera AndreeaÎncă nu există evaluări
- Sistemul NeuromuscDocument67 paginiSistemul NeuromuscAlexandru PurcelÎncă nu există evaluări
- Biofizica Aparatului LocomotorDocument10 paginiBiofizica Aparatului LocomotorAurel BoleaÎncă nu există evaluări
- Teorii Si Modele Bio Chi Mice Ale Contractiei MusculareDocument4 paginiTeorii Si Modele Bio Chi Mice Ale Contractiei Muscularesakura.bomboÎncă nu există evaluări
- Fibra MuscularaDocument39 paginiFibra MuscularaIuliana Alexandra Valcu100% (3)
- Fiziologie LP NR 3 Anul IDocument13 paginiFiziologie LP NR 3 Anul IAnna MariaÎncă nu există evaluări
- 1.5. Fiziologia Fibrei MusculareDocument82 pagini1.5. Fiziologia Fibrei MusculareAlina Topor Psi-MedicalÎncă nu există evaluări
- Curs 2 InimaDocument20 paginiCurs 2 InimaAttila SzakacsÎncă nu există evaluări
- Mecanismul Biochimic Al Contractiei MusculareDocument15 paginiMecanismul Biochimic Al Contractiei Muscularetj_sweetgirlÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularAna-Maria TanasăÎncă nu există evaluări
- Fiziologia Fibrei MusculareDocument52 paginiFiziologia Fibrei MusculareDaniel HuntelarÎncă nu există evaluări
- Tesut Muscular Curs 2019 2020Document10 paginiTesut Muscular Curs 2019 2020rares gherghinaÎncă nu există evaluări
- Mecanismul de ContractieDocument1 paginăMecanismul de ContractieDiana ButacuÎncă nu există evaluări
- Bio ChimDocument22 paginiBio ChimEu VictorÎncă nu există evaluări
- Structura Tesutului MuscularDocument8 paginiStructura Tesutului MuscularVass GeosanuÎncă nu există evaluări
- Muschi NetedDocument6 paginiMuschi Netediuuu3Încă nu există evaluări
- Referat La FiziologieDocument11 paginiReferat La FiziologieFlorin Verdeata100% (1)
- CURS MUSCHI FIZIO PPTDocument46 paginiCURS MUSCHI FIZIO PPTIrina MureșanÎncă nu există evaluări
- Curs 16 AMB IIDocument55 paginiCurs 16 AMB IIDinca Lidia-ElenaÎncă nu există evaluări
- 31 Filamentele de ActinaDocument10 pagini31 Filamentele de ActinaAlina Calmuc100% (1)
- Curs Neuromuscular-Radu BragaDocument14 paginiCurs Neuromuscular-Radu BragaVicențiu Răzvan VornicuÎncă nu există evaluări
- Notite AnatomieDocument20 paginiNotite AnatomieStepByStepÎncă nu există evaluări
- Curs 5Document14 paginiCurs 5sham harbaÎncă nu există evaluări
- BiocelDocument41 paginiBiocelremus_stoica23Încă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Materia tuturor lucrurilor: 12 experimente care au schimbat lumeaDe la EverandMateria tuturor lucrurilor: 12 experimente care au schimbat lumeaÎncă nu există evaluări
- Miere SalcamDocument1 paginăMiere SalcamcurteanancaÎncă nu există evaluări
- Miere CremaDocument2 paginiMiere CremacurteanancaÎncă nu există evaluări
- Album Logopedic-Învățarea SunetelorDocument34 paginiAlbum Logopedic-Învățarea SunetelorcurteanancaÎncă nu există evaluări
- CapacealaDocument2 paginiCapacealacurteanancaÎncă nu există evaluări
- GHID DE TERAPIE ABA Partea A II ADocument239 paginiGHID DE TERAPIE ABA Partea A II AcurteanancaÎncă nu există evaluări
- Ivermectina Se Poate Folosi Si Ca Antibiotic, in Caz de Forta Majora - Medicina BucureştiDocument9 paginiIvermectina Se Poate Folosi Si Ca Antibiotic, in Caz de Forta Majora - Medicina BucureşticurteanancaÎncă nu există evaluări
- Test Rapid Virus Gripal A Si B Realy Tech Manual de Utilizare - 1Document2 paginiTest Rapid Virus Gripal A Si B Realy Tech Manual de Utilizare - 1curteanancaÎncă nu există evaluări
- Vazyme Manual de Utilizare Test Antigen Nazofaringian 1 BucDocument3 paginiVazyme Manual de Utilizare Test Antigen Nazofaringian 1 BuccurteanancaÎncă nu există evaluări
- INGRIJIREA BOLNAVULUI CU FRACTURA DIAFIZARA DE FEMUR referat LUCRARE DE DIPLOMA “INGRIJIREA BOLNAVULUI CU FRACTURA DIAFIZARA DE FEMUR” MOTO: “Ceea ce-l face pe om mai mare decat simpla lui viata, este dragostea pentru viata celorlalti” (L.F. Celine) CUPRINS 1. Partea teoretica 1.1. Notiuni de anatomie 1.2. Fractura, generalitati. Definitie. Clasificare. Simptomatologie Fracturile diafizare ale femurului. Definitie. Etiopatogenie. Simptomatologie. Diagnostic. Evolutie. Complicatii. 1.3.1. Tratament ortopedic. Tratament chirurgical. Tratament de recuperare. 2. Procesul de ingrijire – Notiuni teoretice. 3. Tehnici de nursing 3.1. Primul ajutor 3.2. Analize recoltate 3.3. Pregatire preoperatorie 3.4. Ingrijirea postoperatorie 4. Supravegherea a unui numar de trei bolnavi cu diagnostic de fractura diafizara de femur 4.1. Cazul nr. I 4.2. Cazul nr. 2 4.3. Cazul nr. 3 5. Bibliografie 1.Partea teoretica 1.1. Notiuni de anatomie Anatomia coapseDocument20 paginiINGRIJIREA BOLNAVULUI CU FRACTURA DIAFIZARA DE FEMUR referat LUCRARE DE DIPLOMA “INGRIJIREA BOLNAVULUI CU FRACTURA DIAFIZARA DE FEMUR” MOTO: “Ceea ce-l face pe om mai mare decat simpla lui viata, este dragostea pentru viata celorlalti” (L.F. Celine) CUPRINS 1. Partea teoretica 1.1. Notiuni de anatomie 1.2. Fractura, generalitati. Definitie. Clasificare. Simptomatologie Fracturile diafizare ale femurului. Definitie. Etiopatogenie. Simptomatologie. Diagnostic. Evolutie. Complicatii. 1.3.1. Tratament ortopedic. Tratament chirurgical. Tratament de recuperare. 2. Procesul de ingrijire – Notiuni teoretice. 3. Tehnici de nursing 3.1. Primul ajutor 3.2. Analize recoltate 3.3. Pregatire preoperatorie 3.4. Ingrijirea postoperatorie 4. Supravegherea a unui numar de trei bolnavi cu diagnostic de fractura diafizara de femur 4.1. Cazul nr. I 4.2. Cazul nr. 2 4.3. Cazul nr. 3 5. Bibliografie 1.Partea teoretica 1.1. Notiuni de anatomie Anatomia coapsecurteanancaÎncă nu există evaluări
- 216299Document13 pagini216299curteanancaÎncă nu există evaluări
- Efectuarea EkgDocument4 paginiEfectuarea Ekgcurteananca100% (1)
- Dep NevoiDocument3 paginiDep Nevoicurteananca100% (1)