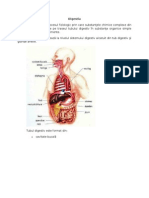
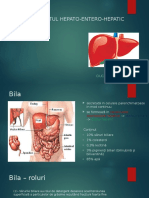
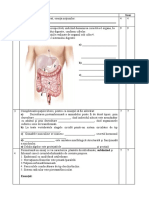
S-ar putea să vă placă și
- Superintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateDe la EverandSuperintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateÎncă nu există evaluări
- Fiziologie - Curs 9Document5 paginiFiziologie - Curs 9ElaEllyÎncă nu există evaluări
- Rolul Si Structura BileiDocument2 paginiRolul Si Structura BileiCosmin LemnaruÎncă nu există evaluări
- Rețete vegetariene din toată lumea. Mâncăruri sănătoase și gustoaseDe la EverandRețete vegetariene din toată lumea. Mâncăruri sănătoase și gustoaseEvaluare: 4 din 5 stele4/5 (8)
- Curs Fiziologia Aparatului Digestiv 3Document35 paginiCurs Fiziologia Aparatului Digestiv 3Andra Maria DinuÎncă nu există evaluări
- Bila-15 03 2022Document3 paginiBila-15 03 2022Ion Florentin100% (1)
- Dieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivDe la EverandDieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivEvaluare: 5 din 5 stele5/5 (1)
- Sistemul Digestiv 2Document40 paginiSistemul Digestiv 2anitasoceanuÎncă nu există evaluări
- Fiziologie ReferatDocument6 paginiFiziologie ReferatReia DrigneiÎncă nu există evaluări
- Anatomia TDDocument13 paginiAnatomia TDTanasa-LucaClaudiaIulianaÎncă nu există evaluări
- Curs Fizio 15Document4 paginiCurs Fizio 15Catalina GabrielaÎncă nu există evaluări
- CD 2Document4 paginiCD 2Mateescu CristianÎncă nu există evaluări
- Biochimia DigestieiDocument6 paginiBiochimia DigestieiReka OrbanÎncă nu există evaluări
- Absorbtia IntestinalaDocument2 paginiAbsorbtia IntestinalaFlorentina TerghesÎncă nu există evaluări
- Digestia IntestinalăDocument28 paginiDigestia IntestinalăitsmeirinaÎncă nu există evaluări
- Secretia Pancreatica ExocrinaDocument21 paginiSecretia Pancreatica ExocrinaNarcisel Patratel100% (2)
- Sistemul Digestiv - Schema Biologie Vegetala Si Animala BacDocument6 paginiSistemul Digestiv - Schema Biologie Vegetala Si Animala BacIvona Mihaescu100% (3)
- 8.rolul Enzimelor in Etapele DigestieiDocument6 pagini8.rolul Enzimelor in Etapele DigestieiPrivateryan91Încă nu există evaluări
- Functia de NutritieDocument29 paginiFunctia de NutritieManea Lucian100% (1)
- Sistemul DigestivDocument6 paginiSistemul DigestivionutÎncă nu există evaluări
- Proprietatile BileiDocument2 paginiProprietatile BileiEmilian RusuÎncă nu există evaluări
- Secretia Pancreatica+bilaDocument42 paginiSecretia Pancreatica+bilaAndrone CameliaÎncă nu există evaluări
- Alcătuirea Sistemului Digestiv: Secreția PancreaticăDocument4 paginiAlcătuirea Sistemului Digestiv: Secreția PancreaticădariaÎncă nu există evaluări
- Proiect La BiologieDocument22 paginiProiect La BiologieTrandafir RoseÎncă nu există evaluări
- AP. DigestivDocument7 paginiAP. Digestivchris_ty_an7256Încă nu există evaluări
- Aparatul DigestivDocument17 paginiAparatul DigestivEla Dany100% (5)
- Biochimia DigestieiDocument6 paginiBiochimia DigestieiLea ElenaÎncă nu există evaluări
- Tema 4.4 Fiziologie STOM ROMDocument4 paginiTema 4.4 Fiziologie STOM ROMstudentÎncă nu există evaluări
- Andrici Marius CosminDocument14 paginiAndrici Marius CosminLacramioara AndriciÎncă nu există evaluări
- Medicația Aparatului DigestivDocument19 paginiMedicația Aparatului DigestiviulianaÎncă nu există evaluări
- Curs 10 - Pancreas, BilaDocument7 paginiCurs 10 - Pancreas, BilaToader Isabela100% (1)
- DigestiaDocument6 paginiDigestiaMeglia GabrielÎncă nu există evaluări
- Digestia Şi Absorbţia SintezeDocument14 paginiDigestia Şi Absorbţia SintezeManuela ConcioiuÎncă nu există evaluări
- Curs 8 Sucuri Digestive.Document6 paginiCurs 8 Sucuri Digestive.Ramona Elena100% (1)
- Secretia PancreaticaDocument5 paginiSecretia Pancreaticacristina_zabavaÎncă nu există evaluări
- New Microsoft Word DocumentDocument5 paginiNew Microsoft Word DocumentHopeÎncă nu există evaluări
- Procesul Digestiei Şi AbsorbţieiDocument23 paginiProcesul Digestiei Şi AbsorbţieiMaricica RausÎncă nu există evaluări
- Proiect - Principii de Nutriție Și Toxicologie AlimentarăDocument17 paginiProiect - Principii de Nutriție Și Toxicologie Alimentarălupu andreiaÎncă nu există evaluări
- Secretie BiliaraDocument6 paginiSecretie BiliaraRoxana Manuela FericeanÎncă nu există evaluări
- Fiziologia Sistemului DigestivDocument5 paginiFiziologia Sistemului DigestivlauraÎncă nu există evaluări
- Circuitul Hepato Entero HepaticDocument10 paginiCircuitul Hepato Entero HepaticMihaela456100% (1)
- Referat Despre Bila Si FicatDocument5 paginiReferat Despre Bila Si FicatAnonymous MhAQS1jJF100% (1)
- Digestia Și AbsorbţiaDocument7 paginiDigestia Și AbsorbţiaIonica Bondei100% (1)
- Metabolismul LipidelorDocument12 paginiMetabolismul LipidelorAndreea Iacob Ioana75% (4)
- 30 Fiziologia Sistemului DigestivDocument6 pagini30 Fiziologia Sistemului Digestivcosmin pesteÎncă nu există evaluări
- Cap 3 Secretia & Motilitatea BiliaraDocument3 paginiCap 3 Secretia & Motilitatea BiliaraAnca FlorescuÎncă nu există evaluări
- Absorbtia IntestinalaDocument2 paginiAbsorbtia Intestinalaİrina Fortu100% (4)
- Curs 12Document3 paginiCurs 12Toader IsabelaÎncă nu există evaluări
- Biologie 11 ADocument6 paginiBiologie 11 Aandrei scorneaÎncă nu există evaluări
- Compoziţia Sucului GastricDocument9 paginiCompoziţia Sucului GastricDaiana Ciobanu100% (3)
- Sistemul DigestivDocument13 paginiSistemul DigestivAna Maria ToporascuÎncă nu există evaluări
- Curs 2Document89 paginiCurs 2입은입은Încă nu există evaluări
- 17 Digestie 2Document14 pagini17 Digestie 2Dana Iulia MoraruÎncă nu există evaluări
- Functiile FicatuluiDocument4 paginiFunctiile FicatuluiGabriela Bulgaru100% (1)
- Bolile Aparatului DigestivDocument55 paginiBolile Aparatului Digestivciobii100% (2)
- Medic in ADocument56 paginiMedic in ACosma CostelÎncă nu există evaluări
- Digestia IntestinalaDocument5 paginiDigestia IntestinalaCosmin Calancia0% (1)
- Componentele Bilei Si Rolurile AceteiaDocument3 paginiComponentele Bilei Si Rolurile Aceteiaagapi ionutÎncă nu există evaluări
- Curs 4Document31 paginiCurs 4Marian Dan IngridÎncă nu există evaluări
- Prim AjutorDocument49 paginiPrim AjutorMarian Dan Ingrid100% (1)
- Constantele Eritrocitului NormalDocument37 paginiConstantele Eritrocitului NormalXiusa TurcanÎncă nu există evaluări
- Partea IIIDocument35 paginiPartea IIIAlina TeodosieÎncă nu există evaluări
- Fiziologia MuşchilorDocument74 paginiFiziologia MuşchilorXiusa TurcanÎncă nu există evaluări
- Prim AjutorDocument36 paginiPrim AjutorMarian Dan IngridÎncă nu există evaluări
- Curs 6, 7, 8Document64 paginiCurs 6, 7, 8Marian Dan IngridÎncă nu există evaluări
- Curs 2Document29 paginiCurs 2Marian Dan IngridÎncă nu există evaluări
- Limba Română Și Artă FotograficăDocument6 paginiLimba Română Și Artă FotograficăXiusa TurcanÎncă nu există evaluări
- NeuronDocument70 paginiNeuronXiusa TurcanÎncă nu există evaluări
- Mark Finley 10 ZileDocument45 paginiMark Finley 10 ZileXiusa Turcan100% (1)
- Test Biologia UmanistDocument4 paginiTest Biologia UmanistXiusa TurcanÎncă nu există evaluări
- Bio TestDocument7 paginiBio TestXiusa TurcanÎncă nu există evaluări
- Bio Test Uman RoDocument6 paginiBio Test Uman RoXiusa TurcanÎncă nu există evaluări
- EnergiaDocument7 paginiEnergiaXiusa TurcanÎncă nu există evaluări
- 1 Decembrie, Ziua Naţională A RomânieiDocument4 pagini1 Decembrie, Ziua Naţională A RomânieiXiusa TurcanÎncă nu există evaluări
- PestisiamfibieniDocument8 paginiPestisiamfibieniOlga RotariÎncă nu există evaluări
- Evaluare Economicul Si Socialul MedievalDocument3 paginiEvaluare Economicul Si Socialul MedievalXiusa TurcanÎncă nu există evaluări
- Ideologii În Epoca ModernăDocument10 paginiIdeologii În Epoca ModernăXiusa Turcan100% (1)
- EnergiaDocument7 paginiEnergiaXiusa TurcanÎncă nu există evaluări
- Numere Intregi in Viata MeaDocument4 paginiNumere Intregi in Viata MeaXiusa Turcan0% (1)
- Crearea Unui SiteDocument1 paginăCrearea Unui SiteXiusa TurcanÎncă nu există evaluări
- Pădurile TropicaleDocument7 paginiPădurile TropicaleAdrian TodericaÎncă nu există evaluări
- Marile Descoperiri GeograficeDocument31 paginiMarile Descoperiri GeograficeXiusa Turcan100% (1)
- Padurile EcuatorialeDocument4 paginiPadurile EcuatorialeElena Botezatu92% (12)
- Evaluare La Lupta Antiotoman (Document3 paginiEvaluare La Lupta Antiotoman (Xiusa TurcanÎncă nu există evaluări
- Romanii in Lupta AntiotomanaDocument45 paginiRomanii in Lupta AntiotomanaXiusa Turcan100% (1)
- ReptileDocument34 paginiReptileOlga RotariÎncă nu există evaluări