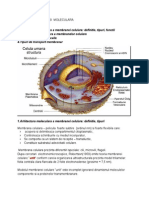
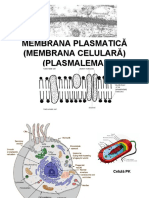
S-ar putea să vă placă și
- Definiţia Şi Funcţiile Membranelor BiologiceDocument23 paginiDefiniţia Şi Funcţiile Membranelor BiologiceDanila ElenaÎncă nu există evaluări
- Subiecte BiocelDocument26 paginiSubiecte BiocelDenisa AndreeaÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Structura Şi Ultrastructura CeluleiDocument34 paginiStructura Şi Ultrastructura CeluleiNadeaÎncă nu există evaluări
- Biocel Cursuri WordDocument38 paginiBiocel Cursuri Wordbubu_lina_ruza100% (2)
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Membrana PlasmaticăDocument56 paginiMembrana PlasmaticăAndrada Raicu100% (1)
- CelulaDocument10 paginiCelulaValentina Gina MuaÎncă nu există evaluări
- Curs 1 - Istoric, Cel Procariota, Eucariota, Membrana CelularaDocument40 paginiCurs 1 - Istoric, Cel Procariota, Eucariota, Membrana CelularaAdrian UrsuÎncă nu există evaluări
- Celula Eucariota Structura Si UltrastructuraDocument19 paginiCelula Eucariota Structura Si UltrastructuraFlorentina Claudia Calusaru100% (2)
- Membrana PlasmaticăDocument62 paginiMembrana PlasmaticăAndra-Amalia ArmulescuÎncă nu există evaluări
- Membrana CelularaDocument4 paginiMembrana CelularaLaura IonitaÎncă nu există evaluări
- Structura Membranei CelulareDocument11 paginiStructura Membranei CelulareOana Palade100% (2)
- Curs MembranaDocument17 paginiCurs MembranaMorgan RosewoodÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Dosar BioDocument17 paginiDosar BioTeodor MoroianuÎncă nu există evaluări
- Curs Biologie Celulara Si MolecularăDocument43 paginiCurs Biologie Celulara Si Molecularăionutbranza19Încă nu există evaluări
- Proteinele Si Lipidele MembranareDocument13 paginiProteinele Si Lipidele MembranareFlorentina Claudia CalusaruÎncă nu există evaluări
- Modelul Mozaicului Fluid Al Membranei CelulareDocument21 paginiModelul Mozaicului Fluid Al Membranei CelulareZgherea MihaiÎncă nu există evaluări
- Celula Animala vs. Celula VegetalaDocument13 paginiCelula Animala vs. Celula VegetalaCristina GabrielaÎncă nu există evaluări
- Celula Animala vs. VegetalaDocument14 paginiCelula Animala vs. VegetalaCristina Gabriela100% (1)
- Celula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Document1 paginăCelula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Gabor IzabellaÎncă nu există evaluări
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
- Celula EucariotaDocument5 paginiCelula EucariotaStefan GhergheÎncă nu există evaluări
- T5,6,7. CelulaDocument16 paginiT5,6,7. CelulaIuliana IrinaÎncă nu există evaluări
- CURS 2 Organizarea StructuralaDocument11 paginiCURS 2 Organizarea StructuralaEugen IuracÎncă nu există evaluări
- Celula Și Fiziologia CelularăDocument26 paginiCelula Și Fiziologia CelularăMARIA-CASANDRA VOICUÎncă nu există evaluări
- Proiect BiologieDocument13 paginiProiect BiologieFlorinAndrabuleaÎncă nu există evaluări
- Curs II CelulaDocument44 paginiCurs II CelulaMaria-Luminita ChiritaÎncă nu există evaluări
- HistologieDocument51 paginiHistologieRoxana CiofalcaÎncă nu există evaluări
- Membrane 2019Document38 paginiMembrane 2019Urecheanu DorinÎncă nu există evaluări
- CELULADocument29 paginiCELULAMadalina AndreeaÎncă nu există evaluări
- Celula Și Fiziologia CelularăDocument16 paginiCelula Și Fiziologia Celularăzilahivanessa799Încă nu există evaluări
- Curs BiocelDocument75 paginiCurs BiocelLavinia BragaÎncă nu există evaluări
- Fenomene de TransportDocument116 paginiFenomene de TransportNoțingher RaduÎncă nu există evaluări
- Cap 3Document8 paginiCap 3Marfescu CristianÎncă nu există evaluări
- CelulaDocument5 paginiCelulaProfu' de biologieÎncă nu există evaluări
- Biocel Sub ExamDocument12 paginiBiocel Sub ExamAlexandra ConstantinescuÎncă nu există evaluări
- Biologia Plantelor - RO - Manual - Celula Vegetala 1 - Particularitati - OrganiteDocument11 paginiBiologia Plantelor - RO - Manual - Celula Vegetala 1 - Particularitati - OrganiteFlorinaBurduceaÎncă nu există evaluări
- Biocel - Subiecte RezolvateDocument94 paginiBiocel - Subiecte RezolvateVictorVasile95% (20)
- Biocel Sub Rezolvate 2009Document25 paginiBiocel Sub Rezolvate 2009ariadna89100% (2)
- Curs 2 BiocelDocument39 paginiCurs 2 BiocelNicoleta NikoÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- Subiecte Nota 6-1Document12 paginiSubiecte Nota 6-1Jowan YassinÎncă nu există evaluări
- Membrana PlasmaticäDocument53 paginiMembrana PlasmaticäFantasyoflove VallyÎncă nu există evaluări
- 2.1. Celula: Celula Reprezintă Unitatea Morfo-Funcțională Și Genetică A OrganizăriiDocument6 pagini2.1. Celula: Celula Reprezintă Unitatea Morfo-Funcțională Și Genetică A OrganizăriiMedeea GabrielaÎncă nu există evaluări
- Celula Miracolul VTZ Evolutia Celulei EucarioteeeeeeDocument19 paginiCelula Miracolul VTZ Evolutia Celulei EucarioteeeeeeGeorgiana NițulescuÎncă nu există evaluări
- Referat Horticultură ID Fiziologia PlantelorDocument13 paginiReferat Horticultură ID Fiziologia Plantelornimeni8Încă nu există evaluări
- TOT CE TREBUIE SĂ ȘTII CA SĂ FII AS LA BIOLOGIE ÎNTR-UN DITAMAI CAIETULDe la EverandTOT CE TREBUIE SĂ ȘTII CA SĂ FII AS LA BIOLOGIE ÎNTR-UN DITAMAI CAIETULEvaluare: 5 din 5 stele5/5 (3)
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- NEUROPSIHOLOGIE: elementele de bază ale problemeiDe la EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiEvaluare: 5 din 5 stele5/5 (1)
- O zi din viața creierului: O perspectivă a neuroștiințeiDe la EverandO zi din viața creierului: O perspectivă a neuroștiințeiÎncă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 3.5 din 5 stele3.5/5 (9)