S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 3.5 din 5 stele3.5/5 (9)
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
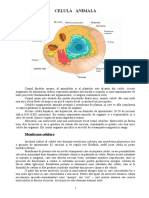
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Curs II CelulaDocument44 paginiCurs II CelulaMaria-Luminita ChiritaÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Celula AnimalaDocument10 paginiCelula AnimalaRaluka Siliște80% (5)
- WWW - Referat.ro-Tesuturi Si Celula30268e3fDocument8 paginiWWW - Referat.ro-Tesuturi Si Celula30268e3fAaron Wolfe100% (1)
- Tesuturile Si Celula AnimalaDocument8 paginiTesuturile Si Celula AnimalaAndreea Ștefura0% (1)
- Celula AnimalaDocument13 paginiCelula AnimalaMina YoungÎncă nu există evaluări
- Celula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Document1 paginăCelula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Gabor IzabellaÎncă nu există evaluări
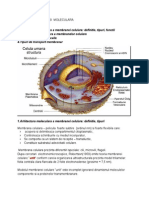
- Celula UmanaDocument14 paginiCelula Umanapaulv123100% (1)
- Scoala Postliceala Sanitara FundeniDocument4 paginiScoala Postliceala Sanitara FundeniAndra MihaelaÎncă nu există evaluări
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- REFERAT Celula UmanaDocument15 paginiREFERAT Celula UmanaIoana100% (2)
- Cap 3Document8 paginiCap 3Marfescu CristianÎncă nu există evaluări
- Celula AnimalaDocument4 paginiCelula AnimalaAlexandra ElaÎncă nu există evaluări
- Organizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternDocument4 paginiOrganizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternVata LuminitaÎncă nu există evaluări
- 1Document10 pagini1Simona StanciuÎncă nu există evaluări
- Celula Animala vs. VegetalaDocument14 paginiCelula Animala vs. VegetalaCristina Gabriela100% (1)
- Celula Eucariota Structura Si UltrastructuraDocument19 paginiCelula Eucariota Structura Si UltrastructuraFlorentina Claudia Calusaru100% (2)
- Anatomie Curs 1Document16 paginiAnatomie Curs 1Danut Ancuța MihaelaÎncă nu există evaluări
- Celula UmanaDocument13 paginiCelula Umana11224499Încă nu există evaluări
- Celula Și Fiziologia CelularăDocument16 paginiCelula Și Fiziologia Celularăzilahivanessa799Încă nu există evaluări
- Celula (Structură, Proprietăţi)Document37 paginiCelula (Structură, Proprietăţi)Sorina DobreÎncă nu există evaluări
- Anatomia Si Fiziologia Omului 191746Document3 paginiAnatomia Si Fiziologia Omului 191746murguÎncă nu există evaluări
- Biocel Cursuri WordDocument38 paginiBiocel Cursuri Wordbubu_lina_ruza100% (2)
- Celula Miracolul VTZ Evolutia Celulei EucarioteeeeeeDocument19 paginiCelula Miracolul VTZ Evolutia Celulei EucarioteeeeeeGeorgiana NițulescuÎncă nu există evaluări
- CelulaDocument10 paginiCelulaValentina Gina MuaÎncă nu există evaluări
- Celula Animala vs. Celula VegetalaDocument13 paginiCelula Animala vs. Celula VegetalaCristina GabrielaÎncă nu există evaluări
- Semiologie Referat Celula UmanaDocument9 paginiSemiologie Referat Celula UmanaCarmen Georgiana EnciuÎncă nu există evaluări
- Biocel Sub ExamDocument12 paginiBiocel Sub ExamAlexandra ConstantinescuÎncă nu există evaluări
- Organite Celulare ComuneDocument13 paginiOrganite Celulare ComuneDennisÎncă nu există evaluări
- Fenomene de TransportDocument116 paginiFenomene de TransportNoțingher RaduÎncă nu există evaluări
- Ce Este CelulaDocument4 paginiCe Este CelularalucaÎncă nu există evaluări
- Celula Și Fiziologia CelularăDocument26 paginiCelula Și Fiziologia CelularăMARIA-CASANDRA VOICUÎncă nu există evaluări
- Celula EucariotaDocument5 paginiCelula EucariotaStefan GhergheÎncă nu există evaluări
- Dosar BioDocument17 paginiDosar BioTeodor MoroianuÎncă nu există evaluări
- 5 Biofizica CelularăDocument70 pagini5 Biofizica CelularăBalint Ana MariaÎncă nu există evaluări
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara Subiectedorofteim100% (1)
- Sinteza Histo IDocument38 paginiSinteza Histo IAdriana ElenaÎncă nu există evaluări
- L 2 Celula Structura ProprietatiDocument10 paginiL 2 Celula Structura ProprietatirahatnikitaÎncă nu există evaluări
- Histo Partialu 1Document10 paginiHisto Partialu 1Evelina UngureanuÎncă nu există evaluări
- Anatomie - Curs1 - CelulaDocument27 paginiAnatomie - Curs1 - Celulaalfa5017Încă nu există evaluări
- NushDocument10 paginiNushcarlos2006conciÎncă nu există evaluări
- WWW - Referateok.ro: NeuronulDocument7 paginiWWW - Referateok.ro: NeuronulBuzatu MadalinaÎncă nu există evaluări
- 2.1. Celula: Celula Reprezintă Unitatea Morfo-Funcțională Și Genetică A OrganizăriiDocument6 pagini2.1. Celula: Celula Reprezintă Unitatea Morfo-Funcțională Și Genetică A OrganizăriiMedeea GabrielaÎncă nu există evaluări
- Organizare Celulare IDocument14 paginiOrganizare Celulare ISmadu AdrianaÎncă nu există evaluări
- Celula Și Organitele CelulareDocument10 paginiCelula Și Organitele CelulareTurlea FloareaÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- Tesutul Conjunctiv Exemplar de Lucru - 2 - 2014Document55 paginiTesutul Conjunctiv Exemplar de Lucru - 2 - 2014AdaBlack23Încă nu există evaluări
- Celula UmanaDocument12 paginiCelula UmanaUchiha AlexandraÎncă nu există evaluări
- Subiecte BiocelDocument26 paginiSubiecte BiocelDenisa AndreeaÎncă nu există evaluări
- Proiect BiologieDocument13 paginiProiect BiologieFlorinAndrabuleaÎncă nu există evaluări
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Cum functioneaza lumea cu adevarat: Ghidul unui om de stiinta pentru trecutul, prezentul si viitorul omeniriiDe la EverandCum functioneaza lumea cu adevarat: Ghidul unui om de stiinta pentru trecutul, prezentul si viitorul omeniriiÎncă nu există evaluări
- Subiecte LP Balneo SabDocument37 paginiSubiecte LP Balneo SabEugen IuracÎncă nu există evaluări
- Onco CursuriDocument108 paginiOnco CursuriEugen IuracÎncă nu există evaluări
- Sexologie - Toate CursurileDocument77 paginiSexologie - Toate CursurileEugen IuracÎncă nu există evaluări
- Curs Complet BalneoDocument65 paginiCurs Complet BalneoEugen Iurac100% (1)
- Curs CompletDocument158 paginiCurs CompletEugen IuracÎncă nu există evaluări
- Cursuri ComfDocument140 paginiCursuri ComfEugen IuracÎncă nu există evaluări
- Curs 3 Spondilita AnchilozantaDocument76 paginiCurs 3 Spondilita AnchilozantaEugen IuracÎncă nu există evaluări
- 18.pericardul Fibros - Descriere, Raporturi, Mijloace de FixareDocument2 pagini18.pericardul Fibros - Descriere, Raporturi, Mijloace de FixareEugen IuracÎncă nu există evaluări
- Cuprins Orientativ Cursuri BalneoDocument5 paginiCuprins Orientativ Cursuri BalneoEugen IuracÎncă nu există evaluări
- Rinichii - Situatie-Sunt Situati Retroperitoneal, Intre Vertebrele T12-L3, Au o Lungime deDocument9 paginiRinichii - Situatie-Sunt Situati Retroperitoneal, Intre Vertebrele T12-L3, Au o Lungime deEugen IuracÎncă nu există evaluări