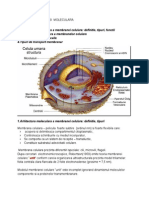
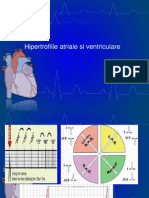
S-ar putea să vă placă și
- Biocel Sub ExamDocument12 paginiBiocel Sub ExamAlexandra ConstantinescuÎncă nu există evaluări
- Subiecte BiocelDocument20 paginiSubiecte BiocelPishatiPishatiÎncă nu există evaluări
- Biologie Celulara - SubiecteDocument50 paginiBiologie Celulara - Subiecteflorinneld100% (2)
- Subiecte Fizio LPDocument27 paginiSubiecte Fizio LPhubertsÎncă nu există evaluări
- Epitelii de Acoperire - Online - AziDocument108 paginiEpitelii de Acoperire - Online - AziFlorentina BogatuÎncă nu există evaluări
- Neuroanatomie SubDocument51 paginiNeuroanatomie SubDaDa1593Încă nu există evaluări
- Membrana CelularaDocument12 paginiMembrana CelularageorgepanciuÎncă nu există evaluări
- Meducatie - Biocel (Membrana, Fct. de Transport, Semnal. Cel, Citoscheletul)Document12 paginiMeducatie - Biocel (Membrana, Fct. de Transport, Semnal. Cel, Citoscheletul)Prunaru BogdanÎncă nu există evaluări
- Sinteza de ATP in MitocondriiDocument3 paginiSinteza de ATP in MitocondriiAndreea TartaÎncă nu există evaluări
- Limfocite BDocument2 paginiLimfocite BJcn KozmyÎncă nu există evaluări
- Curs 4 Tesutul ConjunctivDocument23 paginiCurs 4 Tesutul ConjunctivAadryana100% (1)
- Canale Ionice de Na+Document19 paginiCanale Ionice de Na+andrakris100% (2)
- Tema 4Document68 paginiTema 4Elena PavlicÎncă nu există evaluări
- Curs 1 - Org Sist ImunDocument44 paginiCurs 1 - Org Sist ImunDaniel Ciprian50% (2)
- Transcrierea AdnDocument21 paginiTranscrierea AdnPristavu Mircea-CosminÎncă nu există evaluări
- ScuDocument26 paginiScuMirela OncioiuÎncă nu există evaluări
- Subiecte Fizio LP - RezolvateDocument29 paginiSubiecte Fizio LP - RezolvateDaniela DulgheruÎncă nu există evaluări
- Membrana CelularaDocument6 paginiMembrana Celularaveronica.99Încă nu există evaluări
- Subiecte Fiziologie Practic Sem 2Document37 paginiSubiecte Fiziologie Practic Sem 2Aura ConstantinÎncă nu există evaluări
- Fiziologie SubiecteDocument22 paginiFiziologie SubiecteMareș Ion-AlexandruÎncă nu există evaluări
- Curs 3 Trunchiul CerebralDocument68 paginiCurs 3 Trunchiul CerebralPetrica Taranu100% (2)
- Curs 1 Tesutul EpitelialDocument11 paginiCurs 1 Tesutul EpitelialVerginia ShuganiÎncă nu există evaluări
- Lame BiocelDocument9 paginiLame BiocelMihnea ParaschivÎncă nu există evaluări
- Subiecte Exam - An II - Semi Rezolvate.Document80 paginiSubiecte Exam - An II - Semi Rezolvate.MonicaElenaÎncă nu există evaluări
- Tesutul Conjunctiv - BausicDocument70 paginiTesutul Conjunctiv - BausicRoxana PitaÎncă nu există evaluări
- Biofizica Contractiei MusculareDocument11 paginiBiofizica Contractiei MusculareMircea CiobîcăÎncă nu există evaluări
- Sistemul ComplementDocument25 paginiSistemul ComplementanairamÎncă nu există evaluări
- Subiecte Morfopat An III Sem IDocument61 paginiSubiecte Morfopat An III Sem IElena-Dana Oprea100% (1)
- C XI Partea 9 Controlul Local Si Umoral Al Fluxului Sanguin TisularDocument6 paginiC XI Partea 9 Controlul Local Si Umoral Al Fluxului Sanguin TisularGiugioiu AnamariaÎncă nu există evaluări
- Toleranta MedicamentoasaDocument5 paginiToleranta MedicamentoasaRoxana MargineanÎncă nu există evaluări
- 2015 - Fiziologia Tubului Digestiv Curs 1 PDFDocument36 pagini2015 - Fiziologia Tubului Digestiv Curs 1 PDFAndreea MxdÎncă nu există evaluări
- Curs 01 - 01 - Curs Tesut Epitelial1Document75 paginiCurs 01 - 01 - Curs Tesut Epitelial1Bubulac AngelaÎncă nu există evaluări
- HistologieDocument67 paginiHistologieThoma DanielÎncă nu există evaluări
- FIZIOLOGIE CARDIO-VASCULAR CURS md1Document100 paginiFIZIOLOGIE CARDIO-VASCULAR CURS md1Christina HurstÎncă nu există evaluări
- Examen BMDocument34 paginiExamen BMVirginia ChirutacÎncă nu există evaluări
- Receptor IDocument5 paginiReceptor IAncuta VijanÎncă nu există evaluări
- Comportamentul Uman - Un Complex Biopsihosocial Cu Implicaţii În Prezervarea Sau Alterarea SănătăţiiDocument427 paginiComportamentul Uman - Un Complex Biopsihosocial Cu Implicaţii În Prezervarea Sau Alterarea SănătăţiiMirela OncioiuÎncă nu există evaluări
- Histologie 1 Curs 12) Tesutul NervosDocument7 paginiHistologie 1 Curs 12) Tesutul NervosMariaMaryaÎncă nu există evaluări
- HISTOLOGIE - Lista SubiecteDocument5 paginiHISTOLOGIE - Lista SubiecteDiana Diaconescu100% (3)
- Formarea CelomuluiDocument22 paginiFormarea CelomuluiGeorgian BudaeÎncă nu există evaluări
- Lp4-Varietati de Tesut ConjunctivDocument3 paginiLp4-Varietati de Tesut ConjunctivCrinablue100% (1)
- SNC LP3Document13 paginiSNC LP3Sergiu ScafariuÎncă nu există evaluări
- Curs 1 GeneticaDocument48 paginiCurs 1 GeneticaSelenetyÎncă nu există evaluări
- Master 1Document63 paginiMaster 1Corina GrosuÎncă nu există evaluări
- 2017 MG Histologie LP StudentiDocument32 pagini2017 MG Histologie LP StudentiBianca VornicuÎncă nu există evaluări
- Notiunea de GenaDocument1 paginăNotiunea de GenaIonela MihăilăÎncă nu există evaluări
- Reticul EndoplasmaticDocument29 paginiReticul EndoplasmaticbeibyloveÎncă nu există evaluări
- Subiecte Anatomie Examen Anul 2Document42 paginiSubiecte Anatomie Examen Anul 2Elena Avram100% (2)
- Grila BiocelDocument3 paginiGrila BiocelJackyDanielsÎncă nu există evaluări
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara SubiecteIoana ManoleÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Biologie Celulara SubiecteDocument12 paginiBiologie Celulara SubiecteAndreea AdumitroaieÎncă nu există evaluări
- Biocel Sub Rezolvate 2009Document25 paginiBiocel Sub Rezolvate 2009ariadna89100% (2)
- CURS 2 Organizarea StructuralaDocument11 paginiCURS 2 Organizarea StructuralaEugen IuracÎncă nu există evaluări
- Curs 2 Membrana CelularaDocument12 paginiCurs 2 Membrana CelularaStanciu Sebastian AndreiÎncă nu există evaluări
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Subiecte BiocelDocument22 paginiSubiecte BiocelPishatiPishatiÎncă nu există evaluări
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- HipertrofiileDocument51 paginiHipertrofiileDaniela BocaÎncă nu există evaluări
- Cursuri Bacteriologie MedicalaDocument63 paginiCursuri Bacteriologie MedicalaIulian100% (5)
- HerpesviridaeDocument23 paginiHerpesviridaevladi_roÎncă nu există evaluări
- Intrebari Raspunsuri InfoDocument14 paginiIntrebari Raspunsuri InfoDoraÎncă nu există evaluări
- Curs 4Document45 paginiCurs 4Ana Maria NitaÎncă nu există evaluări
- Preturi Astra Classic 3 2011Document3 paginiPreturi Astra Classic 3 2011dorofteimÎncă nu există evaluări
- Mini Atlas de HistologieDocument6 paginiMini Atlas de HistologieLeonard D100% (19)
- 1 CelulaDocument36 pagini1 CeluladorofteimÎncă nu există evaluări