Documente Academic
Documente Profesional
Documente Cultură
Biologie Celulara Subiecte
Încărcat de
Ioana ManoleTitlu original
Drepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Biologie Celulara Subiecte
Încărcat de
Ioana ManoleDrepturi de autor:
Formate disponibile
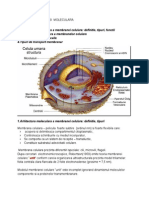
Biologie celulara subiecte: 1.
Conceptul actual despre organizarea membranelor celulare -membrana este o pelicula foarte subtire (cativa nm) si foarte flexibila, ce acopera si delimiteaza compartimentul citoplasmatic, controleaza schimburile cu mediul inconjurator, se comporta ca un sistem de receptie-transductie si constituie substratul adezivitatii celulare. Ea prezinta si diferentieri speciale cum sunt dispozitivele jonctinale, microvilii, cilii, flagelii, etc. -in esenta, continuumul bilipidic, asimetric si fluid , reprezinta axul intregului edificiu molecular; pe cele doua fete hidrofile, proteinele sunt distribuite asimetric si in aranjamente foarte caracteristice; -membrana celulara este alcatuita din trei parti componente principale: a)plasmalema, complexul lipoproteic, in grosime de cca ,! nm; b)glicolema, complexul glicoproteic, superficial in grosime de cca !" nm si c) citoscheletul membranei, reteaua de proteine asezat pe fata interna a plasmalemei in grosime de cativa nm. 2.Compozitia chimica si functiile membranei -in alcatuirea membranelor celulare participa apa cu aproximativ #"$ din masa umeda, substantele organice cu cca "$ si sarurile minerale in cantitati foarte mici. %"$ din masa uscata a membranelor este reprezentata de lipoproteine si ca glucidele sunt numai in cateva procente. &n majoritatea membranelor, proteinele sunt intre !! si '"$ iar lipidele intre #! si ("$. )embranele mielinice fac exceptie majoritatea sunt lipide. *otalitatea lipidelor din suprafata celulei sunt localizate in plasmalema, unde formeaza o foita continua si hidrofoba ce constituie axul in jurul caruia se organizeaza toate celelalte populatii de molecule din membrane. +lucidele sunt prezente exclusiv in glicolema numai sub forma de fragmente de molecule relativ scurte, fie conjugate cu proteine fie atasate la capetele hidrofile a unor lipide de pe fata externa a continumuului lipidic. ,roteinele sunt reprezentate in toate cele trei straturi ale suprafatei celulare - in plasmalema, in principal de o parte si de alta a continuumului lipidic; in glicolema alcatuind tesatura pe care sunt atasate glucidele; in citoscheletul membranei, unde realizeaza o retea foarte fina si foarte supla. )embrana permite patrunderea subst in celula... 3.Lipide membranare definitie si clasificare -toate lipidele membranei celulare sunt localizate in continuumul hidrofob al plasmalemei. .atorita acestui bistrat lipidic, diferitele tipuri de molecule strabat membranele cu viteze diferite.,rincipalele tipuri de lipide ce intra in alcatuirea membranelor sunt fosfolipidele, colesterolul si glicolipidele. 4.Fosfolipide distributie mobilitate -fosfolipidele formeaza marea majoritate a lipidelor din endomembrane si cca !$ din lipidele citolamei. /osfolipidele cele mai frecvent prezente in citomembrane sunt fosfatildicolina, sfingomielina, fosfatidietanolamina si fosfatidilserina. .eoarece fosfolipidele concentreaza la un capat radicalii hidrofili si la celalalt capat radicalii hidrofobi, sunt denumite molecule amfipate sau bimiodale. )oleculele bimodale au proprietatea de a se orienta in monostrat. 0tunci cand concentratia fosfolipidelor este mai mare decat cea necesara pentru a satura interfetele se formeaza micelii. !."ol lipide membranare -confera membranelor plasmatice selectivitate.
#.$roteine membranare generalitati si clasificare -o mare parte din proteinele membranei celulare sunt localizate la nivelul plasmalemei si ele se afla in relatii speciale cu bistratul lipidic. 1aracterizarea cea mai generala a proteinelor plasmalemale poate fi exprimata prin termenii de diversitate si mobilitate. ,roteinele reprezinta substratul specificitatii suprafetei celulare. ,roteinele membranare se pot clasifica in functie de pozitia lor fata de bistratul lipidic, care permite impartirea lor in doua categorii: a)proteine periferice sau extrinseci acele proteine care se afla atasate de o parte si de alta a bistratului lipidic, interactionand cu capetele hidrofile ale lipidelor; b) proteine integrale sau intrinseci acele proteine care sunt cufundate in bistratul lipidic, strabatandu-l complet sau partial. %.$roteine membranare e&emple si descriere -in functie de versantul bistratului lipidic caruia ii sunt asociate se clasifica in ectoproteine - proteine periferice asociate foitei externe a bistratului, asadar expuse la exteriorul membranei celulare sau pe fata luminala a membranelor organitelor; endoproteine - proteine periferice asociate foitei interne a bistratului, asadar expuse pe versantul citoplasmatic al membranelor si endomembranelor. '.(obilitatea proteinelor asociate membranei si rolul ei -mobilitatea proteinelor membranare consta in miscari de rotatie si iscari de translatie. )iscarea de rotatie a proteinelor membranare in jurul propriei axe, denumita si difuzie rotationala, este de cel putin 2"" de ori mai lenta decat a lipidelor. )iscarea de translatie, denumita si difuzie laterala, este si ea mult mai lenta decat a lipidelor, fiind in medie de 2microm3s. ).Citoscheletul asociat membranei descriere functii -este o retea de endoproteine cu ochiuri si noduri, solidara membranei prin atasarea la endodomeniul unor proteine transmembranare. +rosimea acestei structuri membranare este de !-2"nm. 1itoscheletul asociat membranei este solidar si cu citoscheletul citosolic, retea de proteine organizata sub forma de filamente sau microtubuli la nivelul citoplasmei, fiind responsabil de mentinerea si3sau modificarea formei celulelor, ca si geometria membranei. 1*."olul proteinelor membranare -ca si in cazul lipidelor membranare, proteinele au atat rol structural, cat si rol metabolic. 4olurile metabolice ale proteinelor membranare se manifesta in ceea ce priveste schimburile de informatii si substanta dintre celule, sau dintre acestea si mediu. 0stfel proteinele membranare pot si receptori (pt hormoni, factori de crestere, etc), transportatori prin membrana sau transportatori cu membrana. ,roteinele membranare mai pot functiona ca enzime sau ca proteine implicate in semnalizare (proteine +). 11.+licocali& definitie caractere generale -glicocalixul reprezinta invelisul glucidic al celulelor, constituit din structuri oligozaharidice inserate pe lipide ale foitei externe a bistratului, sau pe proteine. .in pct de vedere la structurii biochimice globale glicocalixul este alcatuit din trei componente: glicolipide, glicoproteinele si proteoglicanii. +rosimea medie a glicocalixului este in medie intre 5" si !" nm. &n structura glicocalixului nu participa toate monozaharidele existente in natura. 6umai sapte intra in alcatuirea lanturilor oligozaharidice ale glicolipidelor si glico proteinelor: glucoza, galactoza, manoza, fucoza, n-acetilglucozamina, n-acetil-galactozamina, acizii sialici.
12.Functiile glucidelor membranare -participa la sarina electrica negativa a suprafetei celulare; -necesare, dar nu neaparat suficiente functionarii receptorilor; -participa la fenomene de recunoastere celulara: purtatoarele antigenelor de grup sanguin; extravazarea leucocitelor; fertilizarea(functionarea spermatozoidului cu ovocitul). 13.Conceptul de microdomenii de membrana 14."eceptori intracelulari definitie caracteristici clasificare -receptorii sunt dispozitive moleculare specializate, asezate pe fata externa a membranei celulare, cu ajutorul carora se intercepteaza semnale sau mesaje venite pe cale nervoasa sau pe cale umorala. *ermenul generic de ligand defineste toate substantele care vehiculeaza pe cale umorala sau nervoasa, se leaga de receptori si moduleaza functia celulara. .in categoria liganzilor fac parte unele substante produse de celule care actioneaza specific asupra unui grup de celule 77tinta77 la nivelul carora induc modificari metabolice in concordanta cu necesitatile organismului. Ele sunt denumite mesageri extracelulari sau mesageri de ordinul &. Exemple de mesageri de ordinul & sunt neurotransmitatorii si hormonii. Exista doua mari categorii de receptori:receptori pt substante endogene si receptori pt substante exogene. 1!."eceptori pentru hormoni si neurotransmitatori clasificare -receptorii pt hormoni: dintre acestia mai bine caracterizati sunt receptorii pt insulina care au fost izolati din celulele adipoase si hepatocite. Ei au masa moleculara #""""" dal si pt fiecare celula exista cca 2"""" de receptori pt insulina. .esi receptorii pt ceilalti hormoni nu au fost inca izolati, se considera ca toate celulele 8tinta9 pentru hormoni prezinta receptori care intercepteaza si transmit mesajul hormonal in celula. -receptorii pentru neurotransmitatori sunt situati intotdeauna la nivelul fetei externe a membranei postsinaptice a celulelor nervoase, musculare sau efectoare receptorii influxului nervos. )ai bine cunoscuti sunt receptorii pt acetilcolina, noradrenalina, serotonina, histamina, dopamina, etc. 1#.,&plicati rolul adenilatciclazei membranare de catre receptorii cuplati cu proteine heterometrice 1%.(esageri secunzi definitie e&emple efectori intracelulari -0),-c este considerat mesager de ordinul && in citosol, intrucat determina efecte metabolice specifice induse de mesgerul de ordinul & prin intermediul receptorilor. &n modelul de mesager secundar, ligandul nu intra in celula si impactul cu celula are loc la suprafata membranei celulare. .eci efectele biologice ale ligandului sunt mediate in celula prin 0),-c si nu prin ligand. 1'.-ransport pasi. clasificare e&emple -realizeaza trecerea moleculelor mici prin membrana celulara in sensul gradientului de concentratie, iar in cazul unor ioni in sensul gradientului electrochimic. /olosind energia care rezulta din miscarea moleculelor dintr-o parte in alta in sensul gradientelor, transportul pasiv se face fara consum de energie; este deci independent de metabolismul celular. *ransportul pasiv are loc fie prin difuziune simpla, fie prin difuziune facilitata. 1).Calea de semnalizare /0123-02*.04uaporine semnificatie si implicatii medicale 21.proteine g heterometrice si proteine + mici asemanari si deosebiri 22.(odalitati genetice de reglare a acti.itatii canalelor ionice membranare
23.-ransport acti. clasificare si e&emple -este modalitatea de transport transmembranar caracteristica celulelor vii. El asigura schimburi foarte rapide prin membrana a unor molecule si ioni de importanta vitala pentru celule impotriva gradientelor de concentratie sau a celor electrochimice. &ntrucat transportul de molecule si ioni impotriva gradientelor se face cu consum de energie libera rezultata din metabolism, este metabolic-dependent. *ransportul activ prin pompe ionice.,t mentinerea gradientului ionic intracelular trecerea ionilor prin membrana celulara se face de cele mai multe ori in directia termodinamica nefavorabila, impotriva gradientului de concentratie sau a celui electrochimic.,ompele ionice pot transporta un singur ion (pompa de 1a:) sau concomitent doi ioni (pompa de 6a:) 24.(ecanisme de semnalizare transmembranara .ia receptori cu acti.itate tirozin 5inazica 2!.Care sunt destinatiile posibile ale materialului e&tracelular internalizat prin endocitoza de receptori6 2 e&emple 2#.,tapele fagocitozei -prin fagocitoza celulele transporta din mediul extracelular in citoplasma particule de natura foarte diferita, utilizand regiuni specializate din membrana celulara purtatoare de receptori. &ntr-o prima etapa complexul antigen-opsonina este recunoscut de receptorii de la suprafata fagocitelor si fixat de membrana celulara determinand activarea receptorilor. 0ctivarea receptorilor duce la agregarea proteinelor contractile bogate in actina de la niv citoscheletului si determina emiterea de pseudopode. Ele inconjoara strans particula si ca urmare apar interactiuni receptor-particula pe toata suprafata ei, care fixeaza particula de membrana pseudopodelor. ,seudopodele care inconjoara particula se unesc si la locul de unire fuzioneaza, favorizand formarea unei vezicule care cuprinde particula fara lichid din fluidul extracelular, denumita fagozom. &n citoplasma fagozomii se unesc cu lizozomii care cuprinzand enzime de tipul hidrolazelor acide degradeaza sau digera particulele introduse in fagozomi din mediul extracelular. 2%./onctiunea stransa -jonctiunile stranse sunt dispozitive de adezivitate cu o dispozitie 8in panglica9 impermeabile pentru mar;erii intercelulari separand net compartimentele tisulare cu compozitii chimice diferite. &n organizarea jonctiunilor stranse, membranele adiacente se apropie complet una de alta pentru a edifica structuri pentalaminate impiedicand astfel scurgerea moleculelor printre membranele angajate in organizarea dispozitivului jonctional. ,rincipalele proprietati ale jonctiunilor stranse sunt: a) dispozitive de adezivitate intercelulara flexibile si sigure; b) bariere chimice si fizice intercelulare; c) confera polaritate celulelor angajate in astfel de jonctiuni; d)apar timpuriu intre celulele embrionare de obicei sub forma de macule, pt ca apoi sa dispara sau sa se transforme treptat in zonule pe masura diferentierii celulelor respective. 2'./onctiunea de ancorare pe filamente de actina -jonctiunile focale sunt o varianta a joctiunilor stranse al caror rol preponderent in adezivitatea celulara. <a niv jonctiunilor focale membrana celulei se apropie de substratul sau de adezivitate pana la o distanta de 2"-2! nm. ,e frontul E (sau extracelular) se afla filamente glicoproteice de fibronectina. ,e frontul , (intracelular) al jonctiunii se aglomereaza fascicule de microfilamente matriciale reprezentate de actina si vinculina
care se asocieaza cu membrana plasmatica prin intermediul unor densificari submembranoase 2)./onctiunea de ancorare pe filamente intermediare 3*.Filamente de actina -actina se gaseste in cele mai multe celule in proportie de 2"$ din totalul proteinelor celulare. 1antitatea ei in celule difera dupa gradul de activitate al acestora. )oleculele de actina apar sub forma polimerizata filamentoasa (actina /) in microfilamente fie depolimerizata - monomerica (actina +) in citosol. /ilamentele de actina se asocieaza in parte cu endomembranele si se leaga de obicei de membrana celulara de suprafata. 31.Filamente de miozina -miozina se gaseste in cantitate redusa in celulele nemusculare, fiind de forma si marime diferita in aceeasi celula. <egarea microfilamentelor de membrana de suprafata se realizeaza prin diferite proteine (alfa-actinina si vilina). <egarea microfilamentelor este esentiala pt formarea citoscheletului precum si pt unele procese celulare de baza ca cito;inezia, miscarile filopodiale si microvilare, etc. 32.$roteine asociate actinei 33.Filamente intermediare -sunt prezente in celulele eucariotelor superioare, fiind diferite de microfilamentele de actina si microfilamentele de miozina. =unt numite intermediare pt ca diametrul lor de aproximativ 2" nm este situat intre cel al microfilamentelor mici de actina si cel al microfilamentelor mari de miozina 5!nm. .upa proteinele componente sunt de mai multe tipuri: microfilamente formate din precheratina - cheratina; microfilamente ce contin vimentina; microfilamente ce contin desmina sau scheletina. 34.(icro.ili stereocili cili flageli -microvilii reprezinta o forma de organizare stabila a microfilamentelor, fiind prezenti in diferite tipuri de celule nemusculare. )icrovilii sunt formatiuni ale membranei celulare de suprafata, prezenti de obicei la polul sau polii liberi, nejonctionali, ai unor celule foarte acive in transportul celular. )icrovilii se gasesc astfel bine reprezentati la enterocite sau la nefrocite. .e obicei microvilii au o forma de deget de manusa, au inaltimi si grosimi egale sau inegale. 1ilii apar de obicei sub forma de prelungiri celulare dispuse la unul din ppolii celulei, fiind alcatuiti din citoplasma si membrana celulara. 1ilii pot fi mobili si imobili(stereocili). /lagelii prezinta o structura asemanatoare cililor. <a om ei sunt reprezentati numai prin gametii ssexuali masculini (spermatozoizi) 3!.(icrotubuli -sunt prezenti sub forma de cilindri subtiri in citoplasma celulelor animale si vegetale. &n sectiune transversala au un diametru cuprins intre 5" si #" nm. 1a si microfilamentele, ei pot aparea grupati sau izolati in citoplasma. )icrotubulii sunt in general formatiuni labile datorita moleculelor de tubulina ce se agrega si se dezagrega incontinuu in citoplasma, in functie de concentratia ionilor de 1a: si a moleculelor de 0), ciclic din citosol. Ei au rol in formarea citoscheletului impreuna cu microfilamentele, contribuind la mentinerea formei celulei, participa deasemenea si la transportul axonal mai putand avea si functii speciale 3#.Centriolii centrul celular -centriolii sunt organite microtubulare stabile in citoplasma celulelor. 0vand un diametru de ",2! microni si o lungime pana la ",! microni. 1entrul celular poate contine unul sau frecvent doi centrioli ce sunt inconjurati de o zona citoplasmatica mai clara. 1entrul
celular este responsabil in celula de initierea si declansarea diviziunii celulare (mitoza sau meioza) formand fusul de diviziune. .e asemenea genereaza miscarea cililor si flagelilor prin faptul ca la baza acestora se gaseste intotdeauna prezent un centriol denumit corpuscul bazal. 3%.Centrul de organizare al microtubulilor 3'."ibozomi structura si organizare -sunt organite celulare intracitoplasmatice prezente in toate celulele cu exceptia eritrocitelor. 4ibozomii pot fi liberi in citoplasma (izolati sau grupati) si atasati de membranele reticulului endoplasmic. 4ibozomul este format din doua subunitati inegale ca dimensiuni si inegale in ceea ce priveste constanta de sedimentare: subunitatea mare (diam de #" nm) - o forma sferoidala cu o mica depresiune inspre subunitatea mica si o constanta de sedimentare egala cu '"s; subunitatea mica de forma alungita , convexconcava, cu o constanta de sedimentare de (" s. 4ibozomii contin 046, proteine, cantitati mici de apa diferiti ioni metalici. 3).Functiile ribozomilor -aceste organite celulare reprezinta in citoplasma locul de sinteza al proteinelor. <a nivelul poliribozomilor neatasati membranelor reticulului endoplasmic se sintetizeaza proteinele de structura iar la nivelul ribozomilor atasati membranelor reticulului endoplasmic se sintetizeaza proteinele de export (enzime hormoni anticorpi) 4*.3tructura proteazomilor. 7egradarea proteinelor mediata de ubi4uitina 41.8ncluziuni pigmentare aspect la microscop optic si microscop electronic 2pigmentii apar in mod normal sub forma de granule de melanina (pigment brun negru) in celulele pigmentare sau sub forma de lipofuscina (pigment galben) in celulele nervoase, in unele celule ale corticosuprarenalei sau pe masura imbatranirii organismului in celulele musculare cardiace si in neuroni. &n stari patologice pot aparea si alti pigmenti in citosol cum ar fi hemosiderina, pigmentii biliari, etc. 42.8ncluziuni pigmentare aspect la microscop optic si microscop electronic 43.8ncluziuni de glicogen aspect la microscop optic si microscop electronic 2glucidele apar sub forma de glicogen in celulele animale, in special in celulele hepatice si musculare. &n celulele vegetale, se depun sub forma de amidon. +lucidele se evidentieaza prin colorare cu carmin >est sau prin metoda ,0=, aparand sub forma de gramezi sau plaje. <a microscopul electronic glicogenul se prezinta fie sub forma de particule mici cu aspect de bastonase lungi de 5" - #" nm (particule beta), fie sub forma neregulata sau de rozeta cu diametru de peste 2!" nm (particule alfa). .e obicei, incluziunile glucidice apar in citosol, dar in unele boli de depozit sau in diabet pot fi si in nucleu. 44.Lizozomi biogeneza si functii 2sunt organite celulare de forma sferica sau ovoidala prezenti in citoplasma tuturor celulelor animale cu exceptia hematiilor adulte. <izozomi incep sa se formeze in reticulul endoplasmic rugos unde sunt sintetizate proteinele componente structurale si enzimatice. .e aici pot fi semnalati in unele celule in complexul +olgi si apoi eliberati in citosol. &n alte celule ei vor fi formati direct din reticulul endoplasmic prin sistemul +E4<, scurtcircuitand structurile golgiene. &n alte celule, lizozomii se pot forma prin ambele cai. /unctia lizozomilor: lizozomii sunt organite celulare responsabile de digestia intracelulara prin procesele de heterofagie si autofagie. ?eterofagia este procesul de digestie intracelulara cu ajutorul lizozomilor, a substantelor nutritive, a bacteriilor, a
virusurilor introduse in citoplasma prin endocitoza si fagocitoza. 0uto fagia reprezinta procesul de digestie intracitoplasmatica(cu ajutorul enzimelor lizozomale) a organitelor celulare din celula respectiva care si-au terminat activitatea si se afla in diferite grade de dezintegrare. 4!.$ero&izomi structura si ultrastructura 2sunt organite celulare prezente in citosolul tuturor celulelor animale si vegetale. ,eroxizomii se prezinta sub forma de vacuole sferice sau ovoide, uneori de forme neregulate cu prelungiri. Ei sunt delimitati de o endomembrana groasa de ',!-@ nm si contin o matrice in granulata uneori cu un cristaloid. 1ristaloidul poate aparea format din manunchiuri paralele de tubuli densi la fluxul de electroni care in sectiune transversala dau imaginea unui fagure de miere. 1itomembrana peroxizomala prezinta o compozitie asemanatoare cu a membranei reticulului endoplasmic dar si difera de aceasta prin unele polipeptide si enzime din structura sa. Aneori se poate observa continuitate intre membrana peroxizomala, in special a microperoxizomilor si reticulului endoplasmic neted, dar mai degraba continuitate peroxizom - peroxizom, formand un Breticulul peroxizomal9 , diferit de reticulul endoplasmic. 4#.$ero&izomi biogeneza si functii 2peroxizomii pot lua nastere mai rar direct din reticulul endoplasmic prin dilatarea si desprinderea anumitor parti terminale ale cisternelor acestuia care sunt pozitive pentru catalaza. )ai des peroxizomii se formeaza prin reticulul endoplasmic - complexul +olgi dar inca nu este precizat mecanismul molecular prin care proteinele peroxizomale patrund in organit. =e pare ca unele enzime sunt directionate in peroxizomi direct prin citosol din ribozomii liberi (catalaza, uricaza); altele din reticulul endoplasmic rugos. /unctii: peroxizomii intervin in metabolismul ?5C5 in doua etape: in prima etapa prin actiunea oxidazelor se produce ?5C5 din . si < - aminoacid, acid lactic si alte substante; in a doua etapa, catalaza desface hidrogenul peroxid care este toxic pentru celula, in oxigen si apa. ,eroxizomii pot activa si oxidarea acizilor grasi (palmitatul, laureatul). Ei intervin si in biosinteza glicerolipidelor prin dihidroacetonofosfatacil-transferaza, enzima prezenta in aceste organite. 4%."eticul endoplasmic definitie clasificare forme 2este un organit celular ce se gaseste in toate tipurile de celule cu exceptia hematiei adulte. Crganitul este mai bine reprezentat in celulele angajate in sinteze de proteine, de glucide, de lipide, in celule secretorii exo- si endocrine.organitul este mai dezvoltat in partea mijlocie si interna a citoplasmei si mai putin in partea externa. 4eticulul endoplasmic este de doua tipuri: reticulul endoplasmic rugos si reticulul endoplasmic neted. 4eticulul endoplasmic rugos prezinta pe suprafata externa a membranei ribozomi atasati, izolati sau in grupuri active poliribozomale. 4ibozomii sunt atasati de membranele reticulului prin subunitatea mare, strabatuta de un canal ce se deschide in lumenul reticulului, canal pe care il strabat lanturile polipeptidice ce se formeaza pe ribozomi. 4eticulul endoplasmic neted nu prezinta ribozomi pe membrana sa delimitanta. El comunica adeseori prin canale de legatura cu reticulul endoplasmic rugos. El este alcatuit mai mult din formatiuni tubulare cu un diametru de #" nm. Crganitul este prezent in citoplasma tuturor celulelor dar apare mai putin bine reprezentat decat reticulul endoplasmic rugos. 4'.(ecanisme prin care lantul polipeptidic e transferat din citosol in reticulul endoplasmic
4)."eticulul endoplasmic rol in biosinteza si prelucrarea proteinelor 2reticulul endoplasmic rugos este organitul la nivelul caruia se sintetizeaza proteinele de export. 0cestea se sintetizeaza in aproximativ 2 min la nivelul ribozomilor si poliribozomilor atasati membranelor reticulului, de unde in #-! min, prin canalul aflat in subunitatea mare ribozomala trec in cisternele reticulului. .rumul pe care il parcurg proteinele de la locul de sinteza pana in mediul extracelular realizeaza ciclul secretor celular care cuprinde urmatoarele etape obligatorii: sinteza pe ribozomi, traversarea cisternelor reticulului endoplasmic rugos, maturarea in complexul +olgi, formarea veziculelor secretorii. 1iclul secretor se incheie cu exocitarea produsului sintetizat, respectiv a continutului veziculelor secretorii in spatiul extracelular. !*."eticulul endoplasmic rol in biosinteza lipidelor membranare 2reticulul endoplasmic este angajat si in sinteza unor fosfolipide (lecitina) si a unor glicoproteine integrale din membrana. !1.0paratul +olgi ultrastructura -in organizarea complexului +olgi intra trei elemente componente: pachete de saci turiti sau cisterne asezate in stiva, microvezicule, macrovezicule. *oate cele trei formatiuni sunt delimitate de citomembrane groase de aproximativ '-@ nm, de aspect neted, fara ribozomi atasati pe suprafata lor. =acii sau cisternele golgiene, de aspect turtit sunt paralele unele fata de altele si apar separate prin spatii regulate de 5"-#" nm; sacii contin un material dens omogen, fin granular sau fin fibrilar. 1atre fata imatura sacii golgieni sunt mai mici, mai ingustati, ei devin mai lungi, mai largi si cu segmente mai dilatate catre fata de maturare. 6umarul sacilor golgieni este in general de #-25 pentru fiecare celula, iar in unele celule de tip particular secretor, pot ajunge pana la 5". )icroveziculele au un diametru de 5"-@" nm; sunt inconjurate de o citomembrana groasa de aproximativ ' nm si asezate de obicei spre fata imatura - cis a complexului +olgi. )acroveziculele au un diametru cuprins intre ".2 si ".! micrometri. Ele sunt inconjurate de o citomembrana groasa de @nm si au un continut amorf sau granular-heterogen cu material mai condensat si inegal la fluxul de electroni. )acroveziculele iau nastere din dilatatiile laterale ale sacilor golgieni, care se desprind si se individualizeaza. !2.0paratul +olgi functii -principala functie a aparatului golgi este legata de procesul de secretie intracelulara. 1omplexul golgi are rol in prelucrarea si transportul produsului secretat la nivelul 4.E. catre veziculele de secretie. El condenseaza produsul de secretie primit de la 4.E., il matureaza si il impacheteaza in vezicule secretorii. ,rodusul de secretie poate ajunge la formatiunile golgiene prin microveziculele ce se detaseaza din capetele reticulului endoplasmatic rugos. 0ceste microvezicule functioneaza ca o suveica cu frecvente deplasari dus-intors intre 4.E. si sacii golgieni.. !3.,&ocitoza definitie tipuri !4.3isteme generatoare de energie in celula generalitati si comparatie intre cele doua sisteme principale de sinteza 0-$. !!.(itocondria structura si ultrastructura 2mitocondriile sunt organite prezente in toate tipurile de celule, cu exceptia eritrocitului adult in care lipsesc. Ele se evidentieaza in celula proaspata, nefixata prin colorantul verde Danus, iar in celula fixata prin hematoxilina ferica 4egaud. <a microscopul luminiscent mitocondria aparea sub trei aspecte: granule izolate in citosol, granule
insirate ca margele pe o ata si filamente sau virgule. &ndiferent de cele trei forme optice, examenul la microscopul electronic a aratat ca ele prezinta aceiasi organizare ultrastructurala: o membrana externa, o membrana interna cu criste mitocondriale, un spatiu extern, cuprins intre cele doua membrane si un spatiu intern sau matrice mitocondriala dispusa in interiorul membranei interne si a cristelor. !#.(itocondria compozitia moleculara 2mitocondria este formata din '!- "$ proteine (structurale #"$ si enzime "$), 5!-5@$ lipide (fosfolipide cu predominanta de lecitine, dar si trigliceride, cardiolipin etc), ",!$ 046, o cantitate mica de 0.6, glucide, ioni in diferite concentratii, apa uneori si vitamina 1. intr-o mitocondrie toate proteinele, ca si toate lipidele, sunt inlocuite la aproximativ 5" de zile. !%.(itocondria biogeneza 2in celule, mitocondria se reinnoieste continuu, iar organitele iesite din functie sunt indepartate prin autofagie, cu ajutorul lizozomilor. 0lte noi mitocondrii apar prin condriodiereza, o mitocondrie functionala dand nastere la alte doua organite asemanatoare. !'.(itocondria rol in apoptoza !).9ucleul celular definitie aspect la microscop optic si microscop electronic 2nucleul este organitul caracteristic celulelor eucariote si indeplineste o functie dubla: cea de stocarea informatiei genetice si cea de centru de control al activitatii celulare. <a nucleul vizibil la microscopul electronic se remarca nucleoplasma care cuprinde matricea nucleara, heterocromatina, eucromatina si bucleolul; membrana nucleara cu cisterna nucleara este intrerupta de complexe por din loc in loc. #*.9ucleul celular numar forma localizare 2de regula: o celula - un nucleu. Exista insa numeroase exceptii. Elementul cel mai adesea citat il constituie hepatocitele care sunt binucleate. Exista si celule cu cativa nuclei sau cateva zeci de nuclei. &n conditii normale, cel mai tipic exemplu il reprezinta osteoclastele din tesutul osos. .e obicei forma nucleului urmareste forma celulei si de aceea nucleii apar sub o mare varietate de forme: rotund ovalare, alungit, turtit, lobat, inmugurit. .e cele mai multe ori nucleul ocupa o pozitie centrala in celula, pozitie ce poate fi socotita ca strategica din punct de vedere al rolului sau de coordonare a activitatii celulare. Exista insa relativ numeroase exceptii: nucleu excentric(adipocite); nucleu medio-bazal(in pancreasul exocrin, in parotida), nucleu bazal (celulele calciforme din epiteliul mucoasei intestinale sau traheale), #1.7ogma centrala a biologiei moleculare enunt si semnificatie biologica #2.+ene genom genotip fenotip definitii #3.:nitati genetice si ierarhia lor definitie #4.8n.elis nuclear definitie organizare ultrastructura #!.$orul nuclear transport si generalitati -din loc in loc cele doua membrane nucleare se afla in raporturi de continuitate, delimitand orificii circulare denumite pori sau porus nuclearis. 0cestia delimiteaza un material anular. ,orul impreuna cu materialul anular . porul impreuna cu materialul anular adiacent constituie asa-numitul complex por. 1omplexul por este un component al membranei nucleare prezent in toate tipurile de celule, cu exceptia probabila a celulelor aflate in ultimele faze ale spermiogenezei. 1omplexele por nu sunt specifice pentru membranele nucleare. /ormatiuni similare se intalnesc si in cisternele 4.E. in cisternele
golgiene, precum si in asa-numitele lamele inelate prezente in unele celule fie in citoplasma, fie in nucleu.. ##.9ucleol rol compozitie chimica e.identiere la microscop optic ultrastructura localizare 9;" 2nucleolul are rol in sinteza 046r si biogeneza ribozomilor, rol in transferul 046m si 046s in citoplasma, rol in pregatirea mitozei. ,rincipalele componente chimice ale nucleolului sunt 0.6, 046 si proteine, care se gasesc in functie de tipul celular si momentul functional, in proportie de aprox. #$, $ si %"$ din greutatea uscata. &n celulele vii, la microscopul cu contrast de faza, nucleolul este usor identificabil ca un corpuscul puternic refringent, cu un contur discret neregulat si cu un continut heterogen. 0sezarea nucleolilor in nucleu este de obicei centrala sau paracentrala, dar poate varia in functie de momentul functional. #%.(atricea nucleara ultrastructura si roluri 2matricea nucleara este denumirea atribuita scheletului de natura proteica care inglobeaza cromatina si nucleolii si care se sprijina la exterior pe membrana nucleara. Ea are rol esential in organizarea nucleului si joaca un rol proeminent in sinteza 0.6, sau 046, in medierea semnalelor hormonale si in alte functii nucleare. )atricea nucleara este compusa din doua parti: a) matricea nucleara propriu-zisa reprezentata de scheletul sau reteaua proteica ce se obiectivizeaza atat ultrastructural, cat si biochimic prin extragerea totala a cromatinei, precum si a celei mai mari parti din nucleol. 4eteaua proteica a matricei propriu-zise este alcatuita din proteine stabile de masa moleculara mare, care rezista prelucrarilor ultrastructurale si biochimice; b) fractiunea labila a matricei care este legata lax de reteaua proteica si care este alcatuita din proteina solubila de masa moleculara mica, molecule organice mici, substante anorganice si apa. )atricea nucleara contine si anumite enzime care efectueaza unele procese metabolice inclusiv sinteza de 0.6. si 046. 6ucleul este lipsit de enzimele metabolismului aerob. )atricea contine insa enzimele metabolismului anaerob, pe cele ale formarii legaturilor fosfatmacroergice, enzimele sintezei coenzimelor, precum si 0.6-polimeraza si 046polimeraza. #'.Cromatina definitie clasificare 2cromatina este materialul biologic intranuclear care se coloreaza cu colorantii bazici; cromatina este deci bazofila. .in punct de vedere al colorabilitatii cu coloranti bazici, cromatina se clasifica in doua categorii: a) eucromatina(cromatina Badevarata9) care se coloreaza putin intens cu coloranti bazici este cromatina necondensata - transcriptibila ; b) heterocromatina (Balta9 cromatina) care se coloreaza foarte intens cu coloranti bazici este cromatina condensata - netranscriptibila. #).Fibra de cromatina %*.<istone si proteine non2histone caractere generale si rol -histonele sunt proteine bazice existente numai in genomul eucariotelor. Etimologic, termenul histone sugereaza prezenta lor ubicvitara in tesuturi (nuclei).histonele sunt proteine cu masa moleculara mica; sunt puternic bazice, datorita continutului mare 2"5"$ de aminoacizi bazici, lizina si arginina, purtatori de sarcini pozitive, ceea ce explica legarea lor de 0.6 ; proteine cu o evolutie filogenetica moleculara definita. /unctia majora a histonelor consta in impachetarea 0.6-ului in nucleu, adica organizarea supramoleculara a 0.6-ului sub forma de nucleozomi. ,rotaminele sunt echivalentul
histonelor dar numai in cazul spermatoidului. ,rotaminele sunt proteine cu greutate moleculara foarte mica , cu o lungime medie de numai ## aminoacizi si foarte bazice, bogate in arginina, care reprezinta circa '"$ din reziduurile aminoacidice. &n cadrul secventei celulare care duce la formarea spermatozoidului (spermatogeneza), la trecerea din spermatida in spermatozoid, protaminele inlocuiesc histonele. .atorita protaminelor, cromatina din nucleul spermatozoidului este extrem de condensata si virtual inactiva. 1ompactarea cromatinei din nucleul spermatozoidului este un fenomen esential pentru Bacomodarea9 0.6-ului intr-un volum foarte mic, cum este cel al capului spermatozoidului. %1.,ucromatina -este purtatoare de gene structurale, este portiunea functional activa a cromatinei, este cromatina pe care se face transcriptie. Eucromatina se clasifica in: a)eucromatina activa fractiunea din eucromatina pe care se face continuu transcriptie si asigura viata normala (bazala) a celulei; b)eucromatina permisiva - fractiunea din eucromatina potential activa, ce devine imediat activa ca urmare a actiunii unor semnale specifice modulatoare (de exemplu: hormoni) %2.<eterocromatina. <eterocromatina gonozomala. -se disting doua tipuri de heterocromatina: constitutiva si facultativa. ?eterocromatina constitutiva este o cromatina constant condensata in interfaza. Ea este genetic inactiva, nu contine gene structurale si pe ea nu se face niciodata transcriptie. ?eterocromatina constitutiva contine asa-numitul 06. repetitiv sau satelit, care este un 0.6 in care scurte secvente nucleotidice sunt repetate de un numar foarte mare de ori. ?eterocromatina facultativa este o cromatina condensata care contine gene structurale ce sunt represate(inactive), adica nu se face transcriptie pe ele desi fie s-a facut(sau nu) transcriptie intr-o perioada anterioara, fie se va face (sau nu) transcriptie, daca se transforma temporar (sub influenta unui Bdepresor9) in eucromatina. ?eterocromatina facultativa este de doua feluri in functie de tipul de cromozomi caruia ii corespunde: autozomala si gonozomica (cel mai tipic exemplu, cromatina de sex sau corpusculul >arr). ?eterocromatina gonozomica (corpusculul >arr sau cromatina de sex sau cromatina E) este un corpuscul cromatidian prezent, in mod normal, numai in nucleii celulelor somatice ale persoanelor de sex feminin. El reprezinta in cariotipul feminin (((:EE) condensarea interfazica a unuia dintre cei 5 gonozomi E.
S-ar putea să vă placă și
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara Subiectedorofteim100% (1)
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Biocel Sub Rezolvate 2009Document25 paginiBiocel Sub Rezolvate 2009ariadna89100% (2)
- Biologie Celulara SubiecteDocument12 paginiBiologie Celulara SubiecteAndreea AdumitroaieÎncă nu există evaluări
- Subiecte BiocelDocument22 paginiSubiecte BiocelPishatiPishatiÎncă nu există evaluări
- Subiecte BiocelDocument20 paginiSubiecte BiocelPishatiPishatiÎncă nu există evaluări
- Membrana CelularaDocument4 paginiMembrana CelularaLaura IonitaÎncă nu există evaluări
- Subiecte BiocelDocument26 paginiSubiecte BiocelDenisa AndreeaÎncă nu există evaluări
- Organizarea Membranelor Celulare Si Functiile MembranareDocument25 paginiOrganizarea Membranelor Celulare Si Functiile MembranareCristian ParaianuÎncă nu există evaluări
- CelulaDocument10 paginiCelulaMonica AbbateÎncă nu există evaluări
- Curs 5 Ultrastructura Celulei Bacteriene. 10.03.2009Document5 paginiCurs 5 Ultrastructura Celulei Bacteriene. 10.03.2009Raluca BrumaÎncă nu există evaluări
- Structura Şi Ultrastructura CeluleiDocument34 paginiStructura Şi Ultrastructura CeluleiNadeaÎncă nu există evaluări
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Biologia Plantelor - RO - Manual - Celula Vegetala 1 - Particularitati - OrganiteDocument11 paginiBiologia Plantelor - RO - Manual - Celula Vegetala 1 - Particularitati - OrganiteFlorinaBurduceaÎncă nu există evaluări
- Celula UmanaDocument14 paginiCelula Umanapaulv123100% (1)
- C04 Membrana Celulara JonctiuniDocument46 paginiC04 Membrana Celulara JonctiunipanterabloÎncă nu există evaluări
- Curs 2 Membrana CelularaDocument12 paginiCurs 2 Membrana CelularaStanciu Sebastian AndreiÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- BiocelDocument6 paginiBiocelMada KisanoviciÎncă nu există evaluări
- Fiziologie GeneralaDocument6 paginiFiziologie GeneralaValentino PetreÎncă nu există evaluări
- CURS 2 Organizarea StructuralaDocument11 paginiCURS 2 Organizarea StructuralaEugen IuracÎncă nu există evaluări
- Invelisul CelularDocument4 paginiInvelisul CelularNicola CreangaÎncă nu există evaluări
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- Celula - Fiziologie CelularaDocument56 paginiCelula - Fiziologie CelularaCarmen Stoicescu100% (1)
- Curs 4 Fiziologie An I Ro Seriile BD 2020Document41 paginiCurs 4 Fiziologie An I Ro Seriile BD 2020MITAÎncă nu există evaluări
- REFERAT Celula UmanaDocument15 paginiREFERAT Celula UmanaIoana100% (2)
- Membrana PlasmaticăDocument56 paginiMembrana PlasmaticăAndrada Raicu100% (1)
- Biocel - Subiecte RezolvateDocument94 paginiBiocel - Subiecte RezolvateVictorVasile95% (20)
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Subiect eDocument162 paginiSubiect eMarina MaziluÎncă nu există evaluări
- Curs3b 1Document11 paginiCurs3b 1Andrei CarlanÎncă nu există evaluări
- 49-78 Biocel Bun.Document9 pagini49-78 Biocel Bun.Draga GeorgetaÎncă nu există evaluări
- Curs II CelulaDocument44 paginiCurs II CelulaMaria-Luminita ChiritaÎncă nu există evaluări
- Membrană Celulară2Document9 paginiMembrană Celulară2Cosmin MihoreanuÎncă nu există evaluări
- Curs Biologie Celulara Si MolecularăDocument43 paginiCurs Biologie Celulara Si Molecularăionutbranza19Încă nu există evaluări
- Celula UmanaDocument12 paginiCelula UmanaUchiha AlexandraÎncă nu există evaluări
- Ultrastructura BacteriilorDocument4 paginiUltrastructura BacteriilorDomnișoara AlinaÎncă nu există evaluări
- Celula UmanaDocument13 paginiCelula Umana11224499Încă nu există evaluări
- Biocel Sub ExamDocument12 paginiBiocel Sub ExamAlexandra ConstantinescuÎncă nu există evaluări
- Invelisul CelularDocument5 paginiInvelisul CelularD CatalinÎncă nu există evaluări
- Membrana CelularaDocument6 paginiMembrana Celularaveronica.99Încă nu există evaluări
- OrganiteDocument5 paginiOrganiteAnda Stoica100% (1)
- CELULADocument21 paginiCELULACaroll VilsanÎncă nu există evaluări
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
- BiocelDocument19 paginiBiocelSerdean MaraÎncă nu există evaluări
- Membrana Celulara PDFDocument40 paginiMembrana Celulara PDFOzana CopyShop100% (1)
- Micro CursDocument37 paginiMicro CursMaria JalbaÎncă nu există evaluări
- Matricea ExtracelularaDocument4 paginiMatricea ExtracelularaBizgan Mihaela100% (1)
- Curs 4 BiocelDocument4 paginiCurs 4 BiocelGeorgeMtreixÎncă nu există evaluări
- Biofizica 2003Document17 paginiBiofizica 2003Ioana ManoleÎncă nu există evaluări
- Lp2 Tromb Embol InfarctDocument44 paginiLp2 Tromb Embol InfarctIoana ManoleÎncă nu există evaluări
- IMUNO Sem 12Document42 paginiIMUNO Sem 12Ioana ManoleÎncă nu există evaluări
- Lp2 Tromb Embol InfarctDocument44 paginiLp2 Tromb Embol InfarctIoana ManoleÎncă nu există evaluări
- Carte KinetoterapieDocument258 paginiCarte Kinetoterapiealexandra_m100% (8)
- Teorie MedicoLegala (Radu Vieru)Document295 paginiTeorie MedicoLegala (Radu Vieru)wonder_childÎncă nu există evaluări
- Cerere Certificat DigitalDocument1 paginăCerere Certificat DigitalIoana ManoleÎncă nu există evaluări
- Medii de Cultura Preparate in Flacoane BIOLAB 2011 01 WWW - ClinilabDocument5 paginiMedii de Cultura Preparate in Flacoane BIOLAB 2011 01 WWW - ClinilabIoana ManoleÎncă nu există evaluări
- Scrisoare de Motivatie 1363261Document1 paginăScrisoare de Motivatie 1363261Ioana ManoleÎncă nu există evaluări
- LP 6Document7 paginiLP 6Ioana ManoleÎncă nu există evaluări
- 10 Performanta CardiacaDocument7 pagini10 Performanta CardiacaIoana ManoleÎncă nu există evaluări
- Relaţia MedicDocument2 paginiRelaţia MedicIoana ManoleÎncă nu există evaluări