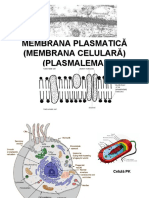
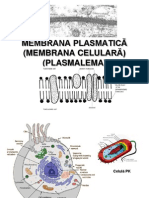
S-ar putea să vă placă și
- Membrane Biologice - BunDocument21 paginiMembrane Biologice - BunMarius Alexa67% (3)
- Biocel - Subiecte RezolvateDocument94 paginiBiocel - Subiecte RezolvateVictorVasile95% (20)
- Invelisul CelularDocument6 paginiInvelisul CelularCutes CameliaÎncă nu există evaluări
- Subiecte Biocel 2013-2014 NEW (Incompleta)Document133 paginiSubiecte Biocel 2013-2014 NEW (Incompleta)Maria ParisÎncă nu există evaluări
- BiocelDocument9 paginiBiocelClaudia SmaduÎncă nu există evaluări
- Biocel Sub Rezolvate 2009Document25 paginiBiocel Sub Rezolvate 2009ariadna89100% (2)
- Cap 4Document32 paginiCap 4cnajesrnasiferfÎncă nu există evaluări
- 1-110 BiocelDocument141 pagini1-110 BiocelBeatrice BondreaÎncă nu există evaluări
- BIOCEL Sinteza-1-1Document45 paginiBIOCEL Sinteza-1-1Adelin FrumusheluÎncă nu există evaluări
- Proteinele Si Lipidele MembranareDocument13 paginiProteinele Si Lipidele MembranareFlorentina Claudia CalusaruÎncă nu există evaluări
- Structura Membranei LipoproteiceDocument10 paginiStructura Membranei LipoproteiceBuliga ClaudiaÎncă nu există evaluări
- Elemente de Transport Prin Membrana CelularaDocument29 paginiElemente de Transport Prin Membrana CelularaIoana CojocaruÎncă nu există evaluări
- Membrana PlasmaticäDocument53 paginiMembrana PlasmaticäFantasyoflove VallyÎncă nu există evaluări
- Arhitectura Membranelor CelulareDocument1 paginăArhitectura Membranelor CelulareSimina MihaiÎncă nu există evaluări
- Membrana CelularaDocument4 paginiMembrana CelularaLaura IonitaÎncă nu există evaluări
- C 4 - Membranele CelulareDocument56 paginiC 4 - Membranele CelulareNoțingher RaduÎncă nu există evaluări
- Curs 2 Membrana CelularaDocument12 paginiCurs 2 Membrana CelularaStanciu Sebastian AndreiÎncă nu există evaluări
- Fenomene de Transport MembranarDocument23 paginiFenomene de Transport MembranarBfkt ProÎncă nu există evaluări
- Curs 9Document17 paginiCurs 9aleahim_aciroivÎncă nu există evaluări
- Membrana PlasmaticăDocument56 paginiMembrana PlasmaticăAndrada Raicu100% (1)
- Membrana Celulara PDFDocument40 paginiMembrana Celulara PDFOzana CopyShop100% (1)
- Reticulul Endoplasmatic - Full CursDocument16 paginiReticulul Endoplasmatic - Full CursStan AdrianÎncă nu există evaluări
- Curs 4-5 PrezDocument13 paginiCurs 4-5 PrezNastasya BalaniciÎncă nu există evaluări
- BiocelDocument26 paginiBiocelAdelin FrumusheluÎncă nu există evaluări
- 4 Transport Membranar MGDocument19 pagini4 Transport Membranar MGmarkopavloski100% (1)
- Biologie Celulara SubiecteDocument12 paginiBiologie Celulara SubiecteAndreea AdumitroaieÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Cito FinalDocument59 paginiCito FinalAndreea ElenaÎncă nu există evaluări
- HISTOLOGIE 1 - Curs 3 - 2022Document16 paginiHISTOLOGIE 1 - Curs 3 - 2022Andreea IlincaÎncă nu există evaluări
- Biocel 1Document56 paginiBiocel 1Corina IonescuÎncă nu există evaluări
- BiocelDocument50 paginiBiocelMariana Sorică100% (1)
- Biologie Celulara - SubiecteDocument50 paginiBiologie Celulara - Subiecteflorinneld100% (2)
- Curs3b 1Document11 paginiCurs3b 1Andrei CarlanÎncă nu există evaluări
- Fenomene de Transport MembranarDocument19 paginiFenomene de Transport MembranarChiriac SimonaÎncă nu există evaluări
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara SubiecteIoana ManoleÎncă nu există evaluări
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara Subiectedorofteim100% (1)
- Metab Lipidelor TotDocument419 paginiMetab Lipidelor TotPetrov MihaelaÎncă nu există evaluări
- TAM4Document37 paginiTAM4Adriana Loredana SecrieriuÎncă nu există evaluări
- Subiecte Biocel 2015Document43 paginiSubiecte Biocel 2015Daniel Mihai GalbenusÎncă nu există evaluări
- Biologie Clasa A IXDocument9 paginiBiologie Clasa A IXLeonte StefanÎncă nu există evaluări
- C 3 - Membranele CelulareDocument56 paginiC 3 - Membranele CelulareSergiuR94Încă nu există evaluări
- BiocelDocument6 paginiBiocelMada KisanoviciÎncă nu există evaluări