S-ar putea să vă placă și
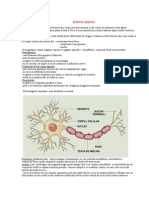
- Fiziologia NeuronuluiDocument14 paginiFiziologia NeuronuluiAdeline ZephyrusÎncă nu există evaluări
- NEUROPSIHOLOGIE: elementele de bază ale problemeiDe la EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiEvaluare: 5 din 5 stele5/5 (1)
- Neuroni Si Celule GlialeDocument20 paginiNeuroni Si Celule GlialeEmma Elena Iancu100% (1)
- Subiecte NeuroanatomieDocument42 paginiSubiecte NeuroanatomieMihai Radulescu100% (2)
- Curs 8 Sistem Nervos OrganizareDocument5 paginiCurs 8 Sistem Nervos OrganizareDelia CristeaÎncă nu există evaluări
- Neuro Tot SubDocument75 paginiNeuro Tot SubDiana CuleaÎncă nu există evaluări
- WWW - Referat.ro Neuronul Celulacheieinorganism 19188Document12 paginiWWW - Referat.ro Neuronul Celulacheieinorganism 19188Joanne Lee100% (1)
- NeuroniiDocument11 paginiNeuroniiAndreeaMariaÎncă nu există evaluări
- Celula Nervoasa Si NevrogliaDocument10 paginiCelula Nervoasa Si NevrogliaAlamentheÎncă nu există evaluări
- Tesut NervosDocument11 paginiTesut NervosSimion CristianÎncă nu există evaluări
- HISTO CURS 13 - Review PDFDocument13 paginiHISTO CURS 13 - Review PDFSerban ElenaÎncă nu există evaluări
- NeuronulDocument32 paginiNeuronulgorcosti7Încă nu există evaluări
- Curs 4 - Țesutul NervosDocument21 paginiCurs 4 - Țesutul NervosAlexandra SavinÎncă nu există evaluări
- L 3 - Sistemul Nervos (Introducere, Maduva Spinarii)Document10 paginiL 3 - Sistemul Nervos (Introducere, Maduva Spinarii)Svetlana Doibani100% (1)
- Cursul 2 - 03.03.2017Document6 paginiCursul 2 - 03.03.2017Ioana StanciuÎncă nu există evaluări
- Tesutul NervosDocument5 paginiTesutul NervosIacob Paula-Izabela100% (1)
- Tesutul NervosDocument10 paginiTesutul NervosToader SilviaÎncă nu există evaluări
- Subiecte Rezolvate SNCDocument5 paginiSubiecte Rezolvate SNCsirbumÎncă nu există evaluări
- NeuronulDocument6 paginiNeuronulFilip Filipelu100% (1)
- Sistemul NervosDocument22 paginiSistemul NervosCarleta Alexiana SîrbuÎncă nu există evaluări
- Sistemul Nervos SomaticDocument16 paginiSistemul Nervos SomaticAna Karisia67% (3)
- Elemente de Teorie NeuronalaDocument12 paginiElemente de Teorie Neuronalagrecu0Încă nu există evaluări
- Str. NeuronuluiDocument7 paginiStr. NeuronuluiLehaci Adelina100% (1)
- NeuropsihologieDocument35 paginiNeuropsihologieValentin Bogdan Vfb100% (1)
- Creierul Si Maduva Spinarii Formeaza Unitatea Centrala Care Prelucreaza ImpulsurileDocument8 paginiCreierul Si Maduva Spinarii Formeaza Unitatea Centrala Care Prelucreaza ImpulsurileOctavian PopoviciÎncă nu există evaluări
- Sistemul Nervos CentralDocument23 paginiSistemul Nervos CentralEduard Gangan100% (2)
- NeuronulDocument4 paginiNeuronulIonela-Georgiana UrseÎncă nu există evaluări
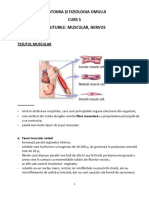
- Anatomia Si Fiziologia Omului. Curs 5 - ŢESUTURILE MUSCULAR, NERVOSDocument7 paginiAnatomia Si Fiziologia Omului. Curs 5 - ŢESUTURILE MUSCULAR, NERVOSSimona GanuÎncă nu există evaluări
- Structura Morfologică A NeuronuluiDocument4 paginiStructura Morfologică A NeuronuluiDiana Florentina AdumitrăceseiÎncă nu există evaluări
- NeuronDocument6 paginiNeuronMaria RalucaÎncă nu există evaluări
- Fiziologia Neuronului I A SinapseiDocument3 paginiFiziologia Neuronului I A Sinapseiemilia2282Încă nu există evaluări
- Neuro Subiecte OralDocument76 paginiNeuro Subiecte OralPuiu AlexandruÎncă nu există evaluări
- NEURONUL Reprezinta Unitatea Structural A Si Functional A A Tesutului NervosDocument13 paginiNEURONUL Reprezinta Unitatea Structural A Si Functional A A Tesutului NervoscaltunlilianaÎncă nu există evaluări
- Note Curs NervosDocument91 paginiNote Curs NervosAlina IrinaÎncă nu există evaluări
- Sistemul Nervos Teorie - OdtDocument6 paginiSistemul Nervos Teorie - OdtBianca PascuÎncă nu există evaluări
- Tesutul NervosDocument17 paginiTesutul NervosLina SorescuÎncă nu există evaluări
- Tesutul Nervos (Suport de Curs Histologie)Document23 paginiTesutul Nervos (Suport de Curs Histologie)Mercea AlexÎncă nu există evaluări
- Neuron UlDocument6 paginiNeuron UlMariana BudnicovÎncă nu există evaluări
- Ţesutul NervosDocument9 paginiŢesutul NervosAndreea Ionela CiobotarÎncă nu există evaluări
- NervulDocument11 paginiNervulviorelÎncă nu există evaluări
- Celulele GlialeDocument3 paginiCelulele GlialeDiana RuicanÎncă nu există evaluări
- Studiul SNC, SNP, MS, Nervi SpinaliDocument9 paginiStudiul SNC, SNP, MS, Nervi SpinaliAdriana FlorentinaÎncă nu există evaluări
- Teoria Eficacității Neuronale, Esența EiDocument10 paginiTeoria Eficacității Neuronale, Esența EiIrina StratilaÎncă nu există evaluări
- NeuronulDocument3 paginiNeuronulsabovlad100% (1)
- NeuronulDocument9 paginiNeuronulBadilas EmiÎncă nu există evaluări
- Anatomie Sistemul NervosDocument25 paginiAnatomie Sistemul NervosRadu RoșianuÎncă nu există evaluări
- Tesutul NervosDocument2 paginiTesutul Nervoszanica2010Încă nu există evaluări
- Țesutul Nervos Este Alcătuit Din Două Tipuri de CeluleDocument15 paginiȚesutul Nervos Este Alcătuit Din Două Tipuri de Celulealessia95xdÎncă nu există evaluări
- Curs 2Document11 paginiCurs 2sham harbaÎncă nu există evaluări
- SN, Maduva SpinariiDocument36 paginiSN, Maduva Spinariilucateodoralin100% (2)
- Slide Uri Transcrise Psihoneurofiziologie PDFDocument163 paginiSlide Uri Transcrise Psihoneurofiziologie PDFIordachi Iulia100% (1)
- NEURONULDocument14 paginiNEURONULDaniela VladÎncă nu există evaluări
- Corpul OmenescDocument45 paginiCorpul Omenescclaudiafrunza207830Încă nu există evaluări
- Tesutul NervosDocument4 paginiTesutul NervosteoctistcostacheÎncă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- O zi din viața creierului: O perspectivă a neuroștiințeiDe la EverandO zi din viața creierului: O perspectivă a neuroștiințeiÎncă nu există evaluări
- IaurtulDocument5 paginiIaurtulManea Elena100% (1)
- Franceza CompuneriDocument5 paginiFranceza CompuneriManea ElenaÎncă nu există evaluări
- AdnDocument18 paginiAdnManea ElenaÎncă nu există evaluări
- 1679Document202 pagini1679Manea ElenaÎncă nu există evaluări
- Franceza CompuneriDocument5 paginiFranceza CompuneriManea ElenaÎncă nu există evaluări
- Ecos 22Document24 paginiEcos 22Manea ElenaÎncă nu există evaluări
- Test de TemperamentDocument3 paginiTest de TemperamentManea ElenaÎncă nu există evaluări
- Proiect de Lunga DurataDocument4 paginiProiect de Lunga DurataManea ElenaÎncă nu există evaluări