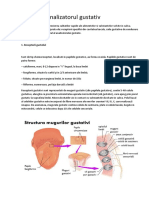
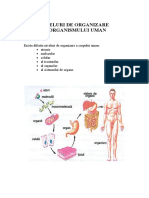
S-ar putea să vă placă și
- Curs 1 - ReceptoriDocument5 paginiCurs 1 - ReceptoriAura ConstantinÎncă nu există evaluări
- Temperamentul CA Latura Expresiva Si DinamicoDocument6 paginiTemperamentul CA Latura Expresiva Si DinamicoiuliancrÎncă nu există evaluări
- AnalizatoriiDocument11 paginiAnalizatoriiDiana LunguÎncă nu există evaluări
- Metabolismul NeuronuluiDocument5 paginiMetabolismul NeuronuluiMiliuhin ConstantinÎncă nu există evaluări
- Reflexe OsteotendinoaseDocument2 paginiReflexe OsteotendinoaseMadalina Marin100% (2)
- Transmiterea SinapticaDocument5 paginiTransmiterea SinapticaLavinia Carpen100% (2)
- Tesutul SecretorDocument21 paginiTesutul SecretorandreeaÎncă nu există evaluări
- Digestia Si AbsorbtiaDocument32 paginiDigestia Si AbsorbtiaCarmen AmuzaÎncă nu există evaluări
- Sistemul MuscularDocument9 paginiSistemul MuscularDiana AndreeaÎncă nu există evaluări
- 5 Explorarea Analizatorilor - Analizatorul VizualDocument71 pagini5 Explorarea Analizatorilor - Analizatorul VizualGeorgiana TrifanÎncă nu există evaluări
- Functiile MaduveiDocument15 paginiFunctiile MaduveiBogdan DraghiciÎncă nu există evaluări
- Morfologia Muschilor Striati 1Document10 paginiMorfologia Muschilor Striati 1AlexColibaÎncă nu există evaluări
- Arsurile ElectriceDocument3 paginiArsurile ElectriceAndreea ManescuÎncă nu există evaluări
- Analizatorul AuditivDocument5 paginiAnalizatorul AuditivCristina LoveÎncă nu există evaluări
- Analizatorul GustativDocument6 paginiAnalizatorul GustativCristina LoveÎncă nu există evaluări
- Activitatea Nervoasă SuperioaraDocument2 paginiActivitatea Nervoasă SuperioaraAgnes BogheanÎncă nu există evaluări
- Examinarea ReflexelorDocument11 paginiExaminarea ReflexelorAlexOO7Încă nu există evaluări
- Transmiterea SinapticaDocument2 paginiTransmiterea SinapticaGheorghe Deliu100% (1)
- Functia de Conducere A Maduvei SpinariiDocument10 paginiFunctia de Conducere A Maduvei SpinariiGeta RobuÎncă nu există evaluări
- C8 - SomesteziaDocument26 paginiC8 - Somesteziablacklovex100% (1)
- Receptor IDocument5 paginiReceptor IAncuta VijanÎncă nu există evaluări
- Ce Este Un Receptor SenzorialDocument2 paginiCe Este Un Receptor SenzorialVadim StavilaÎncă nu există evaluări
- Reflexul RotulianDocument9 paginiReflexul RotulianAlina33% (3)
- 8 Circuite Neuronale 2Document1 pagină8 Circuite Neuronale 2Alex Cu TridentÎncă nu există evaluări
- Nervii CranieniDocument9 paginiNervii CranieniPetrescu AuraÎncă nu există evaluări
- Biologie (2) ReferatDocument2 paginiBiologie (2) ReferatLuka LukichÎncă nu există evaluări
- Rebus DigestieDocument1 paginăRebus DigestieBianca PetrariuÎncă nu există evaluări
- Curs 13 AnalizatoriiDocument58 paginiCurs 13 AnalizatoriiRamona MihaelaÎncă nu există evaluări
- CELULADocument21 paginiCELULACaroll VilsanÎncă nu există evaluări
- Fiziologia SinapseiDocument10 paginiFiziologia SinapseicristÎncă nu există evaluări
- Astrologie Karma FamilialaDocument2 paginiAstrologie Karma FamilialachrishelenÎncă nu există evaluări
- Analizatorul Olfactiv Si GustativDocument4 paginiAnalizatorul Olfactiv Si GustativDiana CismaruÎncă nu există evaluări
- Subiecte Examen Anatomie Proba Scrisă Semestrul IiDocument10 paginiSubiecte Examen Anatomie Proba Scrisă Semestrul IiCristina FrățilescuÎncă nu există evaluări
- Analizatorul VizualDocument6 paginiAnalizatorul VizualIrina ASÎncă nu există evaluări
- Functiile Maduvei, Functiile NeuronuluiDocument1 paginăFunctiile Maduvei, Functiile NeuronuluilavybbyÎncă nu există evaluări
- Másuri de Protectie Privind Posibilele Efecte Ale SunetelorDocument4 paginiMásuri de Protectie Privind Posibilele Efecte Ale Sunetelorgiulia maria baltacÎncă nu există evaluări
- DVL1Document33 paginiDVL1Urecheanu DorinÎncă nu există evaluări
- Neuronii Si Celulele GlialeDocument14 paginiNeuronii Si Celulele GlialeRenata ProsutinskaiaÎncă nu există evaluări
- UrecheaDocument22 paginiUrecheaMiha MihaelaÎncă nu există evaluări
- Rolul EncefaluluiDocument1 paginăRolul EncefaluluiAlexandrucecÎncă nu există evaluări
- Tunica Vasculara O1901Document10 paginiTunica Vasculara O1901Cecan MihaelaÎncă nu există evaluări
- MEDICAMENTEDocument14 paginiMEDICAMENTECostel ChannelÎncă nu există evaluări
- Spectrul de AbsorbțieDocument13 paginiSpectrul de AbsorbțieVlada CovalciucÎncă nu există evaluări
- Neuro Sub IDocument9 paginiNeuro Sub ILoredana Elena UngureanuÎncă nu există evaluări
- Rolul CentrozomuluiDocument8 paginiRolul CentrozomuluiMariana SpiridonÎncă nu există evaluări
- Neuronul Si SinapsaDocument26 paginiNeuronul Si SinapsaEric PopaÎncă nu există evaluări
- Afazia TalamicăDocument1 paginăAfazia Talamicăalessia95xdÎncă nu există evaluări
- Curs 3 - Trunchiul CerebralDocument68 paginiCurs 3 - Trunchiul CerebralAlina Anca100% (1)
- Maduva SpinariiDocument8 paginiMaduva SpinariiKatty BrownÎncă nu există evaluări
- Cito8 CitoscheletDocument42 paginiCito8 Citoscheletadelinamaria654Încă nu există evaluări
- Curs 2 - Prezentarea OralaDocument6 paginiCurs 2 - Prezentarea Oralamadi14yÎncă nu există evaluări
- DiencefalDocument32 paginiDiencefalBiancaÎncă nu există evaluări
- Arcul Reflex ElementarDocument4 paginiArcul Reflex ElementarCalin CalinÎncă nu există evaluări
- C13-14 Arc Reflex Elementar Receptori Musculari Analizator Kinestezic Arcul Reflex Elementar Arcul Reflex ElementarDocument18 paginiC13-14 Arc Reflex Elementar Receptori Musculari Analizator Kinestezic Arcul Reflex Elementar Arcul Reflex ElementarMarius RusuÎncă nu există evaluări
- ReceptoriDocument2 paginiReceptoriLav IniiaÎncă nu există evaluări
- ANATOMIE - Sistemul NervosDocument39 paginiANATOMIE - Sistemul Nervoscchrisstin22Încă nu există evaluări
- RECEPTORIIDocument3 paginiRECEPTORIIAlexe DianaÎncă nu există evaluări
- Analizatorul KinestezicDocument3 paginiAnalizatorul KinestezicBianca BrlcÎncă nu există evaluări
- Cai de Conducere Si Sindroame NervoaseDocument20 paginiCai de Conducere Si Sindroame NervoaseBoglarka27Încă nu există evaluări
- Kinetologie 4Document28 paginiKinetologie 4Aland MediaÎncă nu există evaluări
- ExamenDocument85 paginiExamenAna DubceacÎncă nu există evaluări
- 01.recomandari Prevenire Infectii Virale PT Cabinete Stomatologie MedicinaDocument8 pagini01.recomandari Prevenire Infectii Virale PT Cabinete Stomatologie MedicinaRenata ProsutinskaiaÎncă nu există evaluări
- Preparatele Respirator BacinschiDocument53 paginiPreparatele Respirator BacinschiRenata ProsutinskaiaÎncă nu există evaluări
- Chirurgie Generala Si SemiologieDocument94 paginiChirurgie Generala Si SemiologieNatalia CaramanÎncă nu există evaluări
- Forme Livrare CARDIOTONICEDocument1 paginăForme Livrare CARDIOTONICEAndries TatianaÎncă nu există evaluări
- M1 Tot1 1422Document21 paginiM1 Tot1 1422Bistrea VictorÎncă nu există evaluări
- Patologia Inimii.Document146 paginiPatologia Inimii.Renata ProsutinskaiaÎncă nu există evaluări
- Tema 2infectia ChirurgicalaDocument46 paginiTema 2infectia ChirurgicalaGusan IrinaÎncă nu există evaluări
- Indicatii Ap RespDocument3 paginiIndicatii Ap RespPanfilAlinaÎncă nu există evaluări
- Fizpat TesteDocument29 paginiFizpat TesteRenata ProsutinskaiaÎncă nu există evaluări
- Totalizare III Indicatii MetodiceDocument3 paginiTotalizare III Indicatii MetodiceRenata ProsutinskaiaÎncă nu există evaluări
- Intrebari Atestarea I Test Medicina 1 ROMDocument4 paginiIntrebari Atestarea I Test Medicina 1 ROMRenata ProsutinskaiaÎncă nu există evaluări
- Respirator Indicatii MetodiceDocument3 paginiRespirator Indicatii MetodiceNatalia CaramanÎncă nu există evaluări
- Respirator Forme Med 2022 0Document1 paginăRespirator Forme Med 2022 0Renata ProsutinskaiaÎncă nu există evaluări
- Fizpat1 1Document65 paginiFizpat1 1Renata ProsutinskaiaÎncă nu există evaluări
- 1568490488504825 (2) (1)Document28 pagini1568490488504825 (2) (1)Renata ProsutinskaiaÎncă nu există evaluări
- Bacterie Examen Simu CompletDocument75 paginiBacterie Examen Simu CompletGabriel StratulatÎncă nu există evaluări
- Fizpat 1Document76 paginiFizpat 1Dyana GuaimaranÎncă nu există evaluări
- Cholera, Camp, HelicDocument7 paginiCholera, Camp, HelicRenata ProsutinskaiaÎncă nu există evaluări
- Cartea Final FARMDocument100 paginiCartea Final FARMTatiana Jalba100% (1)
- Atestarea 1 SIMU Stomatologie 2019 RomDocument6 paginiAtestarea 1 SIMU Stomatologie 2019 RomAndrei AndreeviciÎncă nu există evaluări
- Intrebari Pentru Evaluare Curenta Medicina IDocument4 paginiIntrebari Pentru Evaluare Curenta Medicina IVlada LozovanÎncă nu există evaluări
- Tot 2 FizpatDocument52 paginiTot 2 FizpatRenata ProsutinskaiaÎncă nu există evaluări
- Axiol Med 3-4 - 52945Document42 paginiAxiol Med 3-4 - 52945Renata ProsutinskaiaÎncă nu există evaluări
- EKG-Interpretare. HipertofiaDocument95 paginiEKG-Interpretare. HipertofiaVirgil Manole100% (1)
- Axiol. Med. 2.Document28 paginiAxiol. Med. 2.dvgÎncă nu există evaluări
- Atestarea II Medicina I SIMU 2019 Rom - 0Document7 paginiAtestarea II Medicina I SIMU 2019 Rom - 0DanielaÎncă nu există evaluări
- 2 1Document28 pagini2 1Renata ProsutinskaiaÎncă nu există evaluări
- 4Document1 pagină4Renata ProsutinskaiaÎncă nu există evaluări
- Axiol. Med. 1.Document25 paginiAxiol. Med. 1.dvgÎncă nu există evaluări