S-ar putea să vă placă și
- CitoscheletulDocument6 paginiCitoscheletulJessica Smith100% (1)
- Functiile PlasmalemeiDocument17 paginiFunctiile PlasmalemeiDragos IdkÎncă nu există evaluări
- ATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialDocument8 paginiATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialPascu Alexandru0% (1)
- Degradarea Scheletului Atomilor de CarbonDocument11 paginiDegradarea Scheletului Atomilor de CarbonCatalin Apetrei100% (2)
- Formule BiofizicaDocument4 paginiFormule BiofizicaMihai HondruÎncă nu există evaluări
- Ligaturile Lui StaniusDocument2 paginiLigaturile Lui StaniusRareş Secară100% (1)
- Excretia 2Document10 paginiExcretia 2Alex_Burn_99130% (1)
- Subiecte Rezolvate ColocviuDocument18 paginiSubiecte Rezolvate ColocviuCristina IoanaÎncă nu există evaluări
- Adeziune CelularăDocument10 paginiAdeziune CelularăValentina MusatÎncă nu există evaluări
- Fiziologie 2Document11 paginiFiziologie 2Charlie McdonnellÎncă nu există evaluări
- Embriologie Curs 5 Saptamana 3Document71 paginiEmbriologie Curs 5 Saptamana 3LumyÎncă nu există evaluări
- Sinteza de ATP in MitocondriiDocument3 paginiSinteza de ATP in MitocondriiAndreea TartaÎncă nu există evaluări
- LP - OSMOZA Cu Calcul Pres OsmoticaDocument7 paginiLP - OSMOZA Cu Calcul Pres OsmoticaDarth Raz-WanÎncă nu există evaluări
- BIOCEL Sinteza-1-1Document45 paginiBIOCEL Sinteza-1-1Adelin FrumusheluÎncă nu există evaluări
- Matrice ExtracelularaDocument27 paginiMatrice ExtracelularaOana PatriciaÎncă nu există evaluări
- 1b Lanțul Transportor de ElectroniDocument82 pagini1b Lanțul Transportor de ElectroniLaura HutzelÎncă nu există evaluări
- C05 6 TransportDocument41 paginiC05 6 TransportpanterabloÎncă nu există evaluări
- Celula Procariotă, Eucariotă, Diviziunea CelularăDocument27 paginiCelula Procariotă, Eucariotă, Diviziunea Celularăkomamix252Încă nu există evaluări
- LP 1 AdnDocument18 paginiLP 1 AdnGabriela Victoria MartinescuÎncă nu există evaluări
- 6 Fosforilare Oxidativa, Sinteza Proteica, Trafic de MembraneDocument28 pagini6 Fosforilare Oxidativa, Sinteza Proteica, Trafic de MembraneGreta IulianaÎncă nu există evaluări
- Curs 8Document18 paginiCurs 8Georgiana BunghiuzÎncă nu există evaluări
- Tesutul Cartilaginos-2Document4 paginiTesutul Cartilaginos-2more_faithÎncă nu există evaluări
- Membrana CelularaDocument6 paginiMembrana Celularaveronica.99Încă nu există evaluări
- Fiziologie Si Patologie-ColocviuDocument25 paginiFiziologie Si Patologie-ColocviugencimoravaÎncă nu există evaluări
- Curs Biofizik - Canale IoniceDocument5 paginiCurs Biofizik - Canale IoniceFany25Încă nu există evaluări
- Aparatul GolgiDocument3 paginiAparatul Golgipisimiau070% (1)
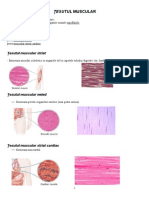
- Tesutul MuscularDocument8 paginiTesutul MuscularGabriel Ungureanu100% (1)
- Notiuni de Termodinamica BiologicaDocument24 paginiNotiuni de Termodinamica BiologicaCristina-Alexandra ConstantinÎncă nu există evaluări
- Biochimie - Subiecte MetabolismDocument84 paginiBiochimie - Subiecte Metabolismdcn0% (1)
- Fiziologia Tesuturilor ExcitabileDocument72 paginiFiziologia Tesuturilor ExcitabileNellu CebotariÎncă nu există evaluări
- Reticulul EndoplasmaticDocument4 paginiReticulul EndoplasmaticIgor HarbuzÎncă nu există evaluări
- Bazele Fizice Şi Biofizice Ale ElectrocardiografieiDocument13 paginiBazele Fizice Şi Biofizice Ale ElectrocardiografieiGabriela Bulgaru100% (1)
- Curs 7 Biochimie - Energetica BiochimicaDocument13 paginiCurs 7 Biochimie - Energetica BiochimicaIonutÎncă nu există evaluări
- BiocelDocument9 paginiBiocelmarian lupuÎncă nu există evaluări
- C 4 AN I - FiziologieDocument22 paginiC 4 AN I - FiziologieVladDanilaÎncă nu există evaluări
- 03 15 51 19partea A 3a - Polaritate Aciditate Proprietati Fizice PDFDocument11 pagini03 15 51 19partea A 3a - Polaritate Aciditate Proprietati Fizice PDFgh4 gh4Încă nu există evaluări
- Tunica Vasculara O1901Document10 paginiTunica Vasculara O1901Cecan MihaelaÎncă nu există evaluări
- Curs 1 Biologie Celulara Si MolecularaDocument10 paginiCurs 1 Biologie Celulara Si MolecularaatezorÎncă nu există evaluări
- CURS 2-Sisteme de Electroliti / Bioelectroliti: "Ce Intelegem Prin Electrolitii?"Document7 paginiCURS 2-Sisteme de Electroliti / Bioelectroliti: "Ce Intelegem Prin Electrolitii?"Leau Sorina-AlexandraÎncă nu există evaluări
- 2020-2021 Membrana CelularaDocument5 pagini2020-2021 Membrana CelularaRéd Ðrågøș100% (1)
- Membrana CelularaDocument25 paginiMembrana CelularaFlavius DobreÎncă nu există evaluări
- 5 Explorarea Analizatorilor - Analizatorul VizualDocument71 pagini5 Explorarea Analizatorilor - Analizatorul VizualGeorgiana TrifanÎncă nu există evaluări
- Curs 6 - MitocondriaDocument17 paginiCurs 6 - MitocondriaRebecca Sandu100% (1)
- PedigreeDocument101 paginiPedigreeElena Filip-MuraruÎncă nu există evaluări
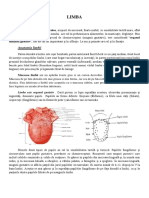
- LimbaDocument3 paginiLimbaSonia MariaÎncă nu există evaluări
- Membrana CelularaDocument9 paginiMembrana CelularaRadu MaziluÎncă nu există evaluări
- RibozomiiDocument22 paginiRibozomiiRaluca LeonteÎncă nu există evaluări
- T1Document12 paginiT1Vasilică SavaÎncă nu există evaluări
- Curs2 Epitelii GlandulareDocument13 paginiCurs2 Epitelii GlandulareAna SismanÎncă nu există evaluări
- Fiziologie TotDocument45 paginiFiziologie TotPasvantu DanielÎncă nu există evaluări
- Epiteliile GlandulareDocument7 paginiEpiteliile GlandulareTheodora Andra MariaÎncă nu există evaluări
- Acizi NucleiciDocument12 paginiAcizi NucleiciMihaela Gaidur100% (1)
- Arcul Reflex VegetativDocument1 paginăArcul Reflex Vegetativalexandra663Încă nu există evaluări
- Regiunea NucalaDocument4 paginiRegiunea NucalaFlavius Ion NicolaeÎncă nu există evaluări
- Organizarea Corpului UmanDocument4 paginiOrganizarea Corpului UmanIuliaNaTuFaruÎncă nu există evaluări
- Curs 1 Tesutul EpitelialDocument11 paginiCurs 1 Tesutul EpitelialVerginia ShuganiÎncă nu există evaluări
- CitoscheletDocument7 paginiCitoscheletAndi ParascaÎncă nu există evaluări
- Citoscheletul PT NurseDocument8 paginiCitoscheletul PT NurseElisabeta IonițăÎncă nu există evaluări
- Matricea Citoplasmatica 2016-1Document15 paginiMatricea Citoplasmatica 2016-1Alex CasyÎncă nu există evaluări
- CitoscheletulDocument24 paginiCitoscheletulnicoletaionelaÎncă nu există evaluări
- Anatomia Omului - Axe, Planuri, OsosDocument134 paginiAnatomia Omului - Axe, Planuri, Ososmiki.elaÎncă nu există evaluări
- EndocrinDocument69 paginiEndocrinAndreea LorenaÎncă nu există evaluări
- Preturi CC Afi - CotroceniDocument4 paginiPreturi CC Afi - CotroceniGheorghe NicoletaÎncă nu există evaluări
- AnalizatoriDocument124 paginiAnalizatoriadelinamaria654Încă nu există evaluări
- Aparatul CardiovascularDocument107 paginiAparatul Cardiovascularcriki_blonduÎncă nu există evaluări
- Digest IVDocument171 paginiDigest IVMaria BindeaÎncă nu există evaluări
- Candidati Admisi PDFDocument21 paginiCandidati Admisi PDFLarisa StanÎncă nu există evaluări
- Anunt Pentru Depunere Burse SocialeDocument5 paginiAnunt Pentru Depunere Burse SocialeValentina Alexandra TudorÎncă nu există evaluări
- Anunt Pentru Depunere Burse SocialeDocument5 paginiAnunt Pentru Depunere Burse SocialeValentina Alexandra TudorÎncă nu există evaluări
- CAMIN U5 Locuri Eliberabile La 30 Septembrie 2020Document1 paginăCAMIN U5 Locuri Eliberabile La 30 Septembrie 2020adelinamaria654Încă nu există evaluări
- CAMIN U5 Locuri Eliberabile La 30 Septembrie 2020Document1 paginăCAMIN U5 Locuri Eliberabile La 30 Septembrie 2020adelinamaria654Încă nu există evaluări
- Candidati Admisi PDFDocument21 paginiCandidati Admisi PDFLarisa StanÎncă nu există evaluări
- Seria 6Document5 paginiSeria 6adelinamaria654Încă nu există evaluări
- Seria 6Document5 paginiSeria 6adelinamaria654Încă nu există evaluări
- Cito6-Golgi, Lizosomi, CitozaDocument31 paginiCito6-Golgi, Lizosomi, Citozaadelinamaria654Încă nu există evaluări
- Preturi CC Afi - CotroceniDocument4 paginiPreturi CC Afi - CotroceniGheorghe NicoletaÎncă nu există evaluări
- Cito11-Comp Extracel, JonctiuniDocument11 paginiCito11-Comp Extracel, Jonctiuniadelinamaria654Încă nu există evaluări
- Cito10-Ciclul CelularDocument33 paginiCito10-Ciclul Celularadelinamaria654Încă nu există evaluări
- Cito3 MembraneDocument26 paginiCito3 Membraneadelinamaria654Încă nu există evaluări
- OrarDocument1 paginăOraradelinamaria654Încă nu există evaluări
- Cito9 MicrotubuliDocument37 paginiCito9 Microtubuliadelinamaria654Încă nu există evaluări
- Cito5-Proteine, REDocument28 paginiCito5-Proteine, REadelinamaria654Încă nu există evaluări
- Cito7-Compartimente EnergeticeDocument21 paginiCito7-Compartimente Energeticeadelinamaria654Încă nu există evaluări
- Cito05 PDFDocument30 paginiCito05 PDFAndreeaÎncă nu există evaluări
- Alarmă: RezistențăDocument1 paginăAlarmă: Rezistențăadelinamaria654Încă nu există evaluări
- Faza de RezistențăDocument1 paginăFaza de Rezistențăadelinamaria654Încă nu există evaluări
- Psihologic. Karl Albrecht, Antreprenor de Succes, Realizează o Clasificare Tetrapartită ADocument1 paginăPsihologic. Karl Albrecht, Antreprenor de Succes, Realizează o Clasificare Tetrapartită Aadelinamaria654Încă nu există evaluări
- Clasa MiriapodeDocument8 paginiClasa Miriapodeadelinamaria654Încă nu există evaluări
- Este Cea Ale Cărei Efecte Sunt Cele Mai Cunoscute Din ManifestărileDocument1 paginăEste Cea Ale Cărei Efecte Sunt Cele Mai Cunoscute Din Manifestărileadelinamaria654Încă nu există evaluări