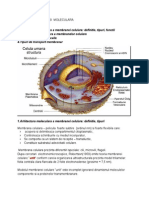
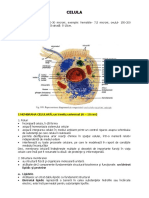
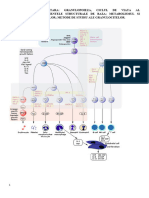
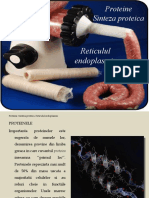
S-ar putea să vă placă și
- CitoscheletDocument7 paginiCitoscheletAndi ParascaÎncă nu există evaluări
- MICROTUBULII BiocelDocument2 paginiMICROTUBULII BiocelLuana-Maria Gherasie0% (1)
- Cito8 CitoscheletDocument42 paginiCito8 Citoscheletadelinamaria654Încă nu există evaluări
- Citoscheletul PT NurseDocument8 paginiCitoscheletul PT NurseElisabeta IonițăÎncă nu există evaluări
- New Microsoft Office Word DocumentDocument9 paginiNew Microsoft Office Word DocumentClaudiu BudÎncă nu există evaluări
- CitoscheletulDocument6 paginiCitoscheletulJessica Smith100% (1)
- C07 CitoscheletulDocument28 paginiC07 Citoscheletulpanterablo100% (1)
- Curs 5Document19 paginiCurs 5Larisa FoteaÎncă nu există evaluări
- Histologie CitoscheletDocument46 paginiHistologie CitoscheletAnamaria ConstantinÎncă nu există evaluări
- Subiecte Biologie CelularaDocument18 paginiSubiecte Biologie CelularaSzobolai MihaelaÎncă nu există evaluări
- Subiecte Biologie CelularaDocument18 paginiSubiecte Biologie CelularaGeorgiana UngureanuÎncă nu există evaluări
- Celula AnimalaDocument1 paginăCelula AnimalaconsumerstefanÎncă nu există evaluări
- Curs 5 - HistologieDocument63 paginiCurs 5 - HistologieAlexandra Florina BrujbanÎncă nu există evaluări
- BioCel Curs 10,11motilitatea CelularaDocument74 paginiBioCel Curs 10,11motilitatea CelularaMarin TheodorÎncă nu există evaluări
- C 3 - CitoscheletulDocument45 paginiC 3 - CitoscheletulNoțingher RaduÎncă nu există evaluări
- Organite Delimitate de EndomembranaDocument9 paginiOrganite Delimitate de EndomembranaStanciu MarinÎncă nu există evaluări
- BIOCEL - MitocondriaDocument5 paginiBIOCEL - MitocondriaVoineta GheorghiuÎncă nu există evaluări
- Rolul Microtubulilor in Mitoză Ref1 BiocelDocument7 paginiRolul Microtubulilor in Mitoză Ref1 BiocelRebeca FlorinaÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- Matricea Citoplasmatica 2016-1Document15 paginiMatricea Citoplasmatica 2016-1Alex CasyÎncă nu există evaluări
- Biologie Celulara SubiecteDocument12 paginiBiologie Celulara SubiecteAndreea AdumitroaieÎncă nu există evaluări
- Mitocondria Procese Și Structuri Implicate În Sinteza Energiei CelulareDocument19 paginiMitocondria Procese Și Structuri Implicate În Sinteza Energiei CelulareGeorgiana SimionÎncă nu există evaluări
- Subiectul 92-110Document30 paginiSubiectul 92-110Andrus MihaelaÎncă nu există evaluări
- HISTOLOGIE 1 - Curs 3 - 2022Document16 paginiHISTOLOGIE 1 - Curs 3 - 2022Andreea IlincaÎncă nu există evaluări
- Celula Si SCH Prin MBDocument10 paginiCelula Si SCH Prin MBTeodora VasileÎncă nu există evaluări
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- CC4 - Relatie celula-MEC PDFDocument24 paginiCC4 - Relatie celula-MEC PDFFlorina GrumezescuÎncă nu există evaluări
- LP10 OrganiteDocument21 paginiLP10 OrganiteAilin Cherim100% (1)
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara SubiecteIoana ManoleÎncă nu există evaluări
- Celula Vegetala Si Tesuturi VegetaleDocument8 paginiCelula Vegetala Si Tesuturi VegetaleBURCEA MARIANAÎncă nu există evaluări
- CitoscheletulDocument24 paginiCitoscheletulnicoletaionelaÎncă nu există evaluări
- Curs 5Document13 paginiCurs 5Denisa NeaguÎncă nu există evaluări
- Biologie CelularaDocument7 paginiBiologie CelularaBogdan CirjanÎncă nu există evaluări
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara Subiectedorofteim100% (1)
- Jonctiuni PDFDocument24 paginiJonctiuni PDFmateiadrian37yahoocomÎncă nu există evaluări
- ROLUL MICROTUBULILOR ÎN MITOZĂ Ref1 BiocelDocument5 paginiROLUL MICROTUBULILOR ÎN MITOZĂ Ref1 BiocelOana KoppandiÎncă nu există evaluări
- CITOSCHELETULDocument35 paginiCITOSCHELETULIoana ScarlatÎncă nu există evaluări
- SeriaDocument11 paginiSeriacretuolgutza91Încă nu există evaluări
- Srbiectul Exemple Proteine Membranare: Mobilitate: DescriereDocument11 paginiSrbiectul Exemple Proteine Membranare: Mobilitate: DescriereMirela OncioiuÎncă nu există evaluări
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- Citoplasma CelularăDocument4 paginiCitoplasma CelularăAndrei PopaÎncă nu există evaluări
- CELULADocument21 paginiCELULACaroll VilsanÎncă nu există evaluări
- AMG Curs 3Document44 paginiAMG Curs 3LorenaÎncă nu există evaluări
- Elemente de Biologie Celulară Şi MolecularăDocument196 paginiElemente de Biologie Celulară Şi MolecularăvalysaÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- Histologie ExamenDocument25 paginiHistologie ExamenMaria Teodora Catană100% (1)
- Curs Nr. 8'Document5 paginiCurs Nr. 8'Co BbÎncă nu există evaluări
- CELULADocument8 paginiCELULAAndreeaÎncă nu există evaluări
- Curs2 BotanicaDocument59 paginiCurs2 Botanicaanm.s2Încă nu există evaluări
- Subiecte BiocelDocument20 paginiSubiecte BiocelPishatiPishatiÎncă nu există evaluări
- CAMIN U5 Locuri Eliberabile La 30 Septembrie 2020Document1 paginăCAMIN U5 Locuri Eliberabile La 30 Septembrie 2020adelinamaria654Încă nu există evaluări
- Seria 6Document5 paginiSeria 6adelinamaria654Încă nu există evaluări
- AnalizatoriDocument124 paginiAnalizatoriadelinamaria654Încă nu există evaluări
- Cito6-Golgi, Lizosomi, CitozaDocument31 paginiCito6-Golgi, Lizosomi, Citozaadelinamaria654Încă nu există evaluări
- Cito5-Proteine, REDocument28 paginiCito5-Proteine, REadelinamaria654Încă nu există evaluări
- Cito3 MembraneDocument26 paginiCito3 Membraneadelinamaria654Încă nu există evaluări
- Clasa MiriapodeDocument8 paginiClasa Miriapodeadelinamaria654Încă nu există evaluări
- Psihologic. Karl Albrecht, Antreprenor de Succes, Realizează o Clasificare Tetrapartită ADocument1 paginăPsihologic. Karl Albrecht, Antreprenor de Succes, Realizează o Clasificare Tetrapartită Aadelinamaria654Încă nu există evaluări
- 2 ProviolentaDocument1 pagină2 Proviolentaadelinamaria654Încă nu există evaluări
- Rez1 PDFDocument7 paginiRez1 PDFadelinamaria654Încă nu există evaluări