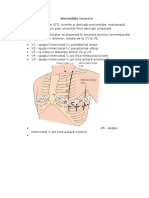
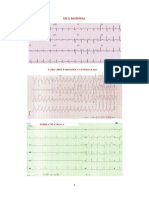
S-ar putea să vă placă și
- 07 Vizual 2017 12 08Document11 pagini07 Vizual 2017 12 08Codrin FodorÎncă nu există evaluări
- Receptia AuditivaDocument10 paginiReceptia AuditivaRadu BulboacaÎncă nu există evaluări
- Metode de Microscopie S.A.Document30 paginiMetode de Microscopie S.A.Mirela BojogaÎncă nu există evaluări
- Nervul Vestibulo-CohlearDocument28 paginiNervul Vestibulo-CohlearGeorge Catalin100% (1)
- Biofizica OpticaDocument7 paginiBiofizica OpticaRotaru Cristian ElizaÎncă nu există evaluări
- Ortopedie RXDocument88 paginiOrtopedie RXAmina GorganÎncă nu există evaluări
- Biopotentiale Celulare Si TisulareDocument19 paginiBiopotentiale Celulare Si TisulareCONSTANTIN CARPENÎncă nu există evaluări
- Canismele de Producere A Traumatismelor CranioDocument19 paginiCanismele de Producere A Traumatismelor CranioAnca Carmen NiculaiasaÎncă nu există evaluări
- Biofizica Receptiei AuditiveDocument14 paginiBiofizica Receptiei AuditiveAndrone Camelia100% (1)
- Tehnologie Si Aparatura MedicalaDocument4 paginiTehnologie Si Aparatura MedicalaOana CretuÎncă nu există evaluări
- Biomecatronica În Medicină - DociuDocument12 paginiBiomecatronica În Medicină - DociuLuminita CojocaruÎncă nu există evaluări
- Implicatii Ale Dezvoltarii Tehnologiei in MedicinaDocument10 paginiImplicatii Ale Dezvoltarii Tehnologiei in MedicinaMaria CristinaÎncă nu există evaluări
- ARTROLOGIADocument9 paginiARTROLOGIAAndraGabrielaOanceaÎncă nu există evaluări
- Bariera Hematoencefalică 2Document12 paginiBariera Hematoencefalică 2Mardari ClaudiuÎncă nu există evaluări
- PlagiDocument31 paginiPlagiDelka AdelÎncă nu există evaluări
- CUL. de Teste. Aparat Locomotor CompletpdfDocument456 paginiCUL. de Teste. Aparat Locomotor CompletpdfBistrea VictorÎncă nu există evaluări
- Fiziologia OchiDocument6 paginiFiziologia OchiDorobantu MarianaÎncă nu există evaluări
- Biofizica CursuriDocument191 paginiBiofizica CursuriTeodor Buse100% (1)
- DializaDocument3 paginiDializaHummel HummelsÎncă nu există evaluări
- Aparate OpticeDocument5 paginiAparate OpticeRobsson RobertÎncă nu există evaluări
- Corectat Referat Audiometrie - CristinaDocument12 paginiCorectat Referat Audiometrie - CristinapaurrasbrÎncă nu există evaluări
- Tesuturile EpitelialeDocument8 paginiTesuturile EpitelialeAlex Seicean100% (1)
- Referat Sanatatea Mediului IDocument4 paginiReferat Sanatatea Mediului IAna Maria Cristina MateiÎncă nu există evaluări
- Biofizica Recepţiei AuditiveDocument22 paginiBiofizica Recepţiei AuditiveValentino PetreÎncă nu există evaluări
- Glandele EndocrineDocument9 paginiGlandele EndocrineMadalina AndreeaÎncă nu există evaluări
- Proiect FinalDocument22 paginiProiect FinalAntonela-Georgiana MateiÎncă nu există evaluări
- Analizatorul VizualDocument2 paginiAnalizatorul VizualAdadinca-26Încă nu există evaluări
- Anatomia, Fiziologia AuditivaDocument8 paginiAnatomia, Fiziologia AuditivaAntonia NarcisaÎncă nu există evaluări
- Analiza Gazelor RespiratoriiDocument6 paginiAnaliza Gazelor RespiratoriiMihaela Cernacovschi100% (2)
- Difuzia PDFDocument2 paginiDifuzia PDFCarolinaMoisaÎncă nu există evaluări
- Imag Ptr. An I MVDocument31 paginiImag Ptr. An I MVbloosomeÎncă nu există evaluări
- Derivatele Laminelor EmbrionareDocument6 paginiDerivatele Laminelor EmbrionareDiana NeofetÎncă nu există evaluări
- Farmacologie Curs 4Document9 paginiFarmacologie Curs 4ayamechanÎncă nu există evaluări
- AmpicilinaDocument2 paginiAmpicilinavalentinanlÎncă nu există evaluări
- Schelet Si Muschi Cap Si GatDocument11 paginiSchelet Si Muschi Cap Si GatIrina CatrinescuÎncă nu există evaluări
- NEUROPSIHOLOGIEDocument3 paginiNEUROPSIHOLOGIEMaria Mădălina100% (1)
- Sub Farmaco Sem1 1Document11 paginiSub Farmaco Sem1 1Simona NeculaÎncă nu există evaluări
- 2 - HumerusDocument13 pagini2 - HumerusJim Boy100% (1)
- LP 1 - Microscopul OpticDocument1 paginăLP 1 - Microscopul OpticEugen IuracÎncă nu există evaluări
- Organe de Simt GrigorasDocument16 paginiOrgane de Simt GrigorasScolobiuc Emma ElenaÎncă nu există evaluări
- Subiecte Exam - An II - Semi Rezolvate.Document80 paginiSubiecte Exam - An II - Semi Rezolvate.MonicaElenaÎncă nu există evaluări
- PERICARDULDocument2 paginiPERICARDULAdriana IuliaÎncă nu există evaluări
- Nervii Cranieni OculomotoriDocument11 paginiNervii Cranieni OculomotoriBianca DenisaÎncă nu există evaluări
- ExteroceptoriiDocument8 paginiExteroceptoriiGiurgiuveanu NicoletaÎncă nu există evaluări
- Curs 5-2 OrlDocument7 paginiCurs 5-2 OrlAdela PașcÎncă nu există evaluări
- Clasa Flagelata 4Document8 paginiClasa Flagelata 4Ienasel AlexandraÎncă nu există evaluări
- Spermatogeneza OvogenezaDocument4 paginiSpermatogeneza OvogenezaM91tinaDrokiaÎncă nu există evaluări
- Receptor IDocument5 paginiReceptor IAncuta VijanÎncă nu există evaluări
- Nervul TrigemenDocument5 paginiNervul TrigemenDanbog BogdanÎncă nu există evaluări
- Radiobiologie GeneralaDocument130 paginiRadiobiologie GeneralaMiruna MafteiÎncă nu există evaluări
- Jonctiuni Celulare NurseDocument5 paginiJonctiuni Celulare NurseasafteioanaÎncă nu există evaluări
- REZUMATE BiocelDocument75 paginiREZUMATE BiocelCristian MargineanÎncă nu există evaluări
- Mecanica RespiratieiDocument6 paginiMecanica RespiratieiRoxana SimonaÎncă nu există evaluări
- CORTICOSUPRARENALADocument27 paginiCORTICOSUPRARENALAAlexandra LatcuÎncă nu există evaluări
- Examenbiochimie PDFDocument78 paginiExamenbiochimie PDFCristi PopoviciÎncă nu există evaluări
- Subiecte BIOCELDocument4 paginiSubiecte BIOCELAndreea NegoiÎncă nu există evaluări
- Biofizica Receptiei AuditiveDocument18 paginiBiofizica Receptiei AuditiveIoana Mihaela SerbanÎncă nu există evaluări
- Aparatul Auditiv Si VizualDocument8 paginiAparatul Auditiv Si VizualAndra Apostol100% (1)
- URECHEADocument25 paginiURECHEAToma GeorgianaaÎncă nu există evaluări
- Analizatorul AuditivDocument15 paginiAnalizatorul Auditivcrr_29Încă nu există evaluări
- SMMRDocument3 paginiSMMRMihai ArcaleanÎncă nu există evaluări
- Proiect Fizica VIII - 0005Document2 paginiProiect Fizica VIII - 0005Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica V - 0005Document2 paginiProiect Fizica V - 0005Mihai ArcaleanÎncă nu există evaluări
- URSSDocument1 paginăURSSMihai ArcaleanÎncă nu există evaluări
- E Sunt Mafiot BEcaliDocument1 paginăE Sunt Mafiot BEcaliMihai ArcaleanÎncă nu există evaluări
- Proiect Fizica VIII - 0008Document2 paginiProiect Fizica VIII - 0008Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica VII - 0002Document2 paginiProiect Fizica VII - 0002Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica V - 0004Document2 paginiProiect Fizica V - 0004Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica VI - 0008Document2 paginiProiect Fizica VI - 0008Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica V - 0004Document2 paginiProiect Fizica V - 0004Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica V - 0004Document2 paginiProiect Fizica V - 0004Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica V - 0004Document2 paginiProiect Fizica V - 0004Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica VI - 0005Document2 paginiProiect Fizica VI - 0005Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica V - 0002Document1 paginăProiect Fizica V - 0002Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica VIII - 0009Document2 paginiProiect Fizica VIII - 0009Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0003Document2 paginiProiect Fizica IX - 0003Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0007Document2 paginiProiect Fizica IX - 0007Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0005Document2 paginiProiect Fizica IX - 0005Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0008Document2 paginiProiect Fizica IX - 0008Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0006Document2 paginiProiect Fizica IV - 0006Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0006Document2 paginiProiect Fizica IX - 0006Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0004Document2 paginiProiect Fizica IX - 0004Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0008Document2 paginiProiect Fizica IV - 0008Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IX - 0002Document2 paginiProiect Fizica IX - 0002Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0010Document1 paginăProiect Fizica IV - 0010Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0009Document2 paginiProiect Fizica IV - 0009Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0005Document2 paginiProiect Fizica IV - 0005Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0006Document2 paginiProiect Fizica IV - 0006Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0007Document2 paginiProiect Fizica IV - 0007Mihai ArcaleanÎncă nu există evaluări
- Proiect Fizica IV - 0004Document1 paginăProiect Fizica IV - 0004Mihai ArcaleanÎncă nu există evaluări