Documente Academic
Documente Profesional
Documente Cultură
Carte de Buzunar Botanica Sem 1
Încărcat de
Holly MollyTitlu original
Drepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Carte de Buzunar Botanica Sem 1
Încărcat de
Holly MollyDrepturi de autor:
Formate disponibile
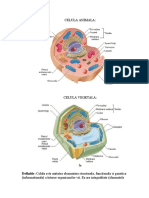
Cap.1 Citologia Citologia este ramura morfologiei plantelor, care se ocupa cu studiul celulei, din toate punctele de vedere.
1.1 Tipuri de celule vegetale Biologia celulara distinge 2 tipuri de celule principale: celula procariota(celule fara nucelu tipic dar cu echivalent al nucelului numit nuceloid), celula eucariota(celule cu nucleu bine diferentiat numit veritabil sau tipic). Celula procariota- specifica bacteriilor si algelor albastre verzi.celule procariote au o structura relativ simpla, lipsesc majoritatea organitelor celulare cu exceptia ribozomilor.La nivelul lor sunt prezente cele mai importante cai metabolice implicate in glicoliza, respiratie, fotosinteza.Materialul genetic este reprezentat de o macromolecula de ADN impachetat intr-o maniera complexa, cu un grad mare de condensare. Celula eucariota- contin mai mult ADN localizat intr-un nucleu si inalte organite celulare(mitocondrii,cloroplaste).Mai prezinta un citoschelet implicat in organizarea citoplasmei si in mobilitate.Peretele celular procariot este necelulozic, iar cel eucariot este celulozic.Legaturile intre celulele eucariote sunt asigurate de plasmodisme( legaturi citoplasmatice) fapt ce conduce la un organism pluricelular veritabil. 1.2Structura si functiile celulei vegetale eucariote Celula vegetala este alcatuita dintr-un perete celular, hialoplasma si organite celulare(RE, dictiozomi, rizozomi,preoxizomi,mitocondrii, plastide, nucleu). 1.2.1 Componentele protoplasmatice Biomembranele reprezinta totalitatea membranelor biologice, care au o structura de baza comuna, fiind alcatuite din molecule lipoproteice mentinute impreuna prin legaturi necovalente.Sunt de 2 feluri: plasmalema( membrana celulara ce delimiteaza fiecare celula in raport cu mediul inconjurator, reprezentand limita externa a protoplastului. Functioneaza ca un filtru de inalta selectivitate mentinand hemostazia.este o structura dinamica, polarizata si asimetrica) citomembranele(compartimenteaza organitele celulare, au permeabilitate selectiva specifica asigurand un anumit comportament functional al organitelor in structura carora intra.Existenta lor impiedica interferenta reactiilor enzimatice simultane din cadrul organitelor). Citoplasma are 2 faze distincte: citosolul(plasma fundamentala sau hialoplasma.compozitia ei 85-95% apa, aminoacizi, monozaharide, acizi grasi, ioni, saruri minerale.) si granuloplasma(faza particulara a citoplasmei reprezentata de diferite organite celulare si de citoschelet. Citoscheletul este alcatuit din 3 categorii de elem de natura proteica: a)microfilamentele;b)filamente intermediare;c)microtubuli.). Reticolul endoplasmatic- organit extins format din cavitati intracitoplasmatice, delimitate de o singura membrana ce reprezinta mai mult de 50 % din membranele celulare.Membrana sa se continua cu membrana externa a invelisului nuclear, lumenul sau fiind despartit doar de membrana nucleara interna de continutul nuclear.Desmotubulii, asigura legaturile intercelulare, intrand in structura plasmodesmelor.Contine numeroase enzime inclusiv pe cele implicate in sinteza lipidelor.Intalnim 2 tipuri de RE in functie de prezenta ribozomilor(sunt structuri mici de natura ribonuceloproteica, cu rol in sinteza proteinelor, sunt de 2 tipuri: 70S si 80 S.)a)- RE neted, b)RE rugos. Aparatul Golgi- totalitatea dictiozomilor formeaza complexul golgi.Componente:1)cisterne sau saci turtiti; 2)vezicule mari; 3- vezicule mici.Acesta se mareste odata cu celula, este o structura polarizata: fata cis asociata fetei RE rugos si fata trans unde se formeaza macrovezicule.Are rolul de a directiona traficul din celula(alcatuieste macromolecule).COntine in membrana dictiozomilor markeri responsabili de recunoasterea proteinelor ce trebuiesc transportate. Lizozomii- vezicule cu o singura membrana specializate in digestia intracelulara.COntine o mare varietate de enzime hidrolitice(hidrolaze acide).Enzimele sunt produse de RE apoi impachetate la nivelul dictiozomilor(au aspect rugos).Sunt de 2 tipuri: primari si secundari( se formaza din dictiozomi primari impreuna cu substratul ce trebuie hidrolizat). Peroxizomii- componente protoplasmatice cu o singura membrana ce au rol in reactii oxidative.Sunt de 2
tipuri : implicati in fotorespiratie, transforma oxigenul in dioxid de carbon; si glioxizomi(transforma acizii grasi in glucide in interiorul semintelor germinate). Mitocondria- organit celular cu 2 membrane, autonoma(capabila de autoreproducere).Sunt de 3 tipuri: 1sferica sau usor ovala numita mitocondrie; 2- in forma de bastonas numita condrioconta; 3- in forma de sirag numita condriomita.Invelisul dublu membranar formeaza 2 compartimente: spatiul intermembranar si spatiul matriceal. Plastidele formeaza plastidomul; sunt componente dublu membranate, semiautonome si autoreproductibile.Sunt de 2 tipuri: 1- plastide fotosintetizatoare(cloroplaste, rodoplaste, )2- plastide nefotosintetizatoare(leucoplaste, cromoplaste).Cloroplastele sunt organite implicate in fotosinteza ce transforma radiatia solara si seva bruta in produsi organici si oxigen; sunt de forma biconvexa alcatuite din membrana dubla(peristrom), spatiu stromatic si spatiu tilacoidal. Nucleul- controleaza intreaga activitate celulara pe baza informatiei stocata biochimic in ADN(poate fi transcris, translatat, replicat).Dimensiunile, forma si pozitia nucleului tin cont de tipul celulei, starea si functionalitatea ei. In celule cu activitate sintetica ridicata nucleul ocupa 3/4 din volumul celular, se reduce cu imbatranirea. Dpvd fiziologic nucleul se poate afla in 3 faze: 1- interfaza; 2- de divizune; 3- de travaliu(coordoneaza activitatea celulara in celule care nu se mai divid).Nucleul interfazic este alcatuit din : invelis nuclear, stroma nucleara si nucleol.Invelisul nuclear(anvelopa nucleara ) delimiteaza continutul nuclear in raport cu citoplasma, este alc din 2 membrane concentrice, un spatiu perinuclear, si o lamina nucleara.Cele 2 membrane sunt diferite din punct de vedere biochimic, si sunt in interactiune cu citoplasma(cea externa) si nucleoplasma (cea interna).Lamina nucleara se observa sub forma unui strat electono dens pe fata interna a membranei interne spre nucleoplasma.Este perforat, prezinta pori acolo unde cele 2 membrane au fuzionat. Stroma nucleara formata din 3 categorii de componente: nucleoplasma(partea fluida apoasa diferita prin compozitie de citosol), matricea nucleara(scheletul microfibrilar de care sunt ancorate fibrele de cromatina), cromatina( forma cromozomilor interfazici ).Fibra de cromatina este alcatuita din ADN si proteine histonice, proteinele avand rolul de suport pentru macromolecula de ADN fiica si induce structura secundara si tertiara a ADN-ului, pe langa faptul ca blocheaza accesibiliateatea ADN-ului pentru enzime si mareste stabilitatea acestuia.Fibra de cromatina trece prin mai multe faze de impachetare, cea mai simpla: fibra nucleosomala este alcatuita din o regiune centrala(proteine histonice)+ un segment de ADN + un segment de ADN linker. Nucleolul contine o mare cantitate de ARN si proteine; este fara membrana dar foarte specializat. Prezinta 3 componente partial individualizate.Este responsabil de producerea celor 2 subunitati ribozomale 40S si 60 S ale lui 80S.Pot depasi 25% din volumul nucleului in celulele cu activitate proteosintetica ridicata. 1.2.2 Componentele neprotoplasmatice Sucul vacuolar se gaseste in vacuole, au o singura membrana denumita tonoplast.Totalitatea vacuolelor formeaza vacuomul. Vacuolele iau nastere in celulele tinere prin fuzionarea veziculelor ce apartin aparatului Golgi si RE.Au 2 functii: de stocare a apei si de stocare a componentelor organice.Sucul vacuolar(incolor) contine apa, subst anorganice(NaCl, MgCl2, Azotati, fosfati) si subst oragnice(monozaharide, acizi grasi, glucozide, alcaloizi, aminoacizi, taninuri, si uleiuri). Substante ergastice produsi finali/intermediari ai metabolismului celular care pot sau nu intra in activitatea metabolica a celulelor. Pot fi grupate in 2 categorii: 1- substante ergastice organice(amidon, aleurora, inulina, mucilagii, gumele, cauciucul, alcaloizii) 2- subst ergastice anorganice(cristale minerale, CaSO4, SiO2,CaC03). 1.3Diviziunea celulara Manifestarile caracteristice viului: cresterea, dezvoltarea si reproducerea se realizeaza la nivelul lumii vegetale pe cai variate a caror esenta nu este in total cunoscuta.Ciclul de viata al unei celule vegetale comporta 2 etape distincte: interfaza, care cuprinde 3 faze succesive(G1= presintetica,S= de sinteza, G2= postsintetica) si faza de diviziune care nu este parcursa de toate celulele corpului. O celula poate parcurge caile:1- parcurge mai departe ciclul celular complet, dand nastere la celule identice cu ea(celule meristematice) 2- se poate opri pentru un timp in interfaza(remeristematizarea celulelor parnechimatice) 3- se poate opri definitiv si inreversibil in interfaza(celule puternic specializate) 4- poate sufei la un moment dat mioza (gametii); 5- poate suferi modificari morfostructura;le puternice.Diviziunea consta in 2 procese succesive : 1- diviziunea nucleului(cariochineaza) 2- diviziunea citoplasmei (citochineza).
MITOZA.Diviziunea nucleului (cariochineza ecvationala) numita si mitoza cuprinde etapele : 1.Profaza- cromozomii incep sa se scurteze si sa se ingroase devenind vizibili la microscop.Fiecare cromozom e duplicat(replicatia ce a avut loc in faza de sinteza a interfazei). Biogeneza ribozomilor e stopata iar nucleolii se fragmenteaza, apoi se dezorganizeaza in interiorul nucleului. 2.Prometafaza-incepe odata cu dezorganizarea invelisului nuclear si patrunderea fusului de diviziune in arealul nucleului.Fragmentele invelisului nuclear raman vizibile in jurul fusului de diviziune pe tot parcursul cariochinezei. 3.Metafaza- in cursul intregii cariochineze cromozomii se comporta pasiv iar miscarile lor sunt rezultatul interactiunii fibrelor chinetocorale cu restul fusului de diviziune si a interactiunii microtubulilor polari intre ei. 4.Anafaza- separarea sincrona a cromozomilor dedublati in cromozomi monocromatidici, definitivarea replicarii ADN-ului in regiunea centromerului si inceputul deplasarii cromozomilor spre polii fusului de diviziune.La sfarsitul anafazei cromozomii ajung la jumatatea distante dintre ecuator si polii fusului de diviziune. 5.Telofaza- cromozomii individualizati ajung la poli, iar fibrele chinetocorale in urma depolimerizarii totale se reface invelisul nuclear formandu-se nucleele interfazice ale celor 2 celule fiice, procesul foloseste materiale rezultate din dezorganizarea nucleului celulei mame. Citochineza se realizeaza in interiorul celulei prin constituirea unui perete celular nou, intre cele 2 celule fiice, perpendicular pe fibrele fusului de diviziune.Peretele celular se formeaza in asociere cu microtubulii polari reziduali ai fusului de diviziune.La acestea se adauga profile de RE(fragmoplast).Peretele celular primordial disciform se extinde progresiv pana atinge peretele celular primar. MEIOZA.Asigura formarea celulelor reproducatoare(haploide), mijlocind astfel reproducerea organismelor vegetale.Mecanismul meiozei cuprinde 2 diviziuni succesive: meioza 1 (etapa reductionala , se injumatateste nr de cromozomi) si meioza 2 (etapa ecvationala in care se realizeaza multiplicarea nucleilor haploizi rezultati din meioza 1).Interfaza, etapa pregatitoare meiozei, include in replicatia ADN faza S premeiotica fiind mult mai lunga decat cea premeiotica. Profaza 1.(Leptoten, zigoten, pachitem, diplotem, diachineza). Leptoten-cindensare gradata a cromozomilor ce devin vizibili la microscop.La sfarsitul leptotenului cromozomii omoligi si partial condensati si gata de asociere. Zigoten- cromozomii omologi formeaza sinapse rezultand cromozomii bivalenti(au 4 cromatide).Imperecherea cromozomilor omologi se realizeaza in 2 etape succesive.Sfarsitul zigotenului apare odata cu edificarea sinapsei. Pachiten- cromozomii omologi sunt tinuti impreuna de catre complexul psinaptimental si are loc crossingoverul.Aceasta faza dureaza foarte mult. Diploten-desfacerea complexului sinaptinemal si se recunoaste dupa tendinta de indepartare a cromozomilor omologi din tetrade.Acestia raman totusi conectati prin chiasme(unde a a avut loc crossingoverul). Diachineza- cromozomii se spiralizeaza, nucleolii si invelisul nuclear se dezorganizeaza. Metafaza 1- dezasamblarea invelisului nuclear si aparitia fusului de diviziune.Cromozomii omologi(legati prin chiasme) formeaza in zona ecuatoriala placa ecuatoriala dubla unde centomerii cromozomilor sunt orientati spre polii opusi ai fusului de diviziune. Anafaza 1- Cromozomii bicromatidici incep sa se apropie de poli . Telofaza 1-se incheie cu formarea a 2 nuceli fii haploizi, delimitati de catre un invelis nuclear propriu si localizati in interiorul aceleiasi celula mama. Interchineza- Aceasta perioada nu este similara interfazei mitotice deoarece nu mai au loc procesele de sinteza ale ADN, ARN si proteine. Profaza 2- cromozomii incep sa se scurteze si sa se ingroase devenind vizibili la microscop.Fiecare cromozom e duplicat(replicatia ce a avut loc in faza de sinteza a interfazei). Biogeneza ribozomilor e stopata iar nucleolii se fragmenteaza, apoi se dezorganizeaza in interiorul nucleului. Metafaza 2- centromerii se divid iar cromatidele se separa dand nastere la cate 2 cromozomi noi, individualizati care insa din cauza crossing-overului nu sunt identici ca in cazul mitozei. Anafaza 2-separarea sincrona a cromozomilor dedublati in cromozomi monocromatidici, definitivarea replicarii ADN-ului in regiunea centromerului si inceputul deplasarii cromozomilor spre polii fusului de diviziune.La sfarsitul anafazei cromozomii ajung la jumatatea distante dintre ecuator si polii fusului de diviziune.
Telofaza 2- se incheie cu formarea a 4 nuclee haploide respectiv a 4 celule haploide distincte. Generarea elementelor reproducatoare haploide necesita o reducere a nr de cromozomi.Meioza este diviziunea celulara ce asigura la plante producerea gametilor. Cap.2 Histologia(studiaza tesutul vegetal) Tesutul esteo grupare permanenta de celule de aceeasi forma si origine, independente care indeplinesc aceeasi functie.celulele ce alcatuiesc un tesut sunt unite intre ele, o unitate anatomica permanenta superioara celulei.UN tesut are 3 caracteristici: 1- unitate anatomica permanenta alcatuita din celule interconectate prin lamela mijlocie, puctuatiuni si plasmodesme capabila sa functioneze ca un tot unitar; 2- unitate fiziologica bazata pe specializarea celulelor componente pentru anumite functii; 3- unitate de integrare distincta subordonata organismului.Tesuturile se impart in 5 sisteme: sistemul generator; sistemul protector; sistemul conducator; sistemul fundamental; sistemul tesuturilor speciale. 2.1 Sistemul generator Dezvoltarea organismelor generale nu este un fenomen finit.Meristemele (tesuturi de origine) alcatuiesc sistemul generator care determina cresterea si dezvoltarea si formarea celulelor reproducatoare.Din inmultirea lor se produce initial un material celular relativ omogen care ulterior se diversifica pe baza procesului de diversitate celulara.Cresterea si dezvoltarea se realizeaza pe baza a 2 procese: 1diferentiere celulara produse de meristeme primare;2-dediferentiere celulara, formata de meristeme secundare. 2.1.1.Meristeme primordiale- sunt alcatuite din una sau mai multe celule initiale si din derivatele lor cele mai apropiate.Ele sunt focalizate in varfurile de crestere ale axeleor supraterane si subterane denumite varfuri vegetative meristematice.Celulele initiale se divid mitotic formand in general 2 celule fiice integrale dintre care cea mai mica va functiona in continuare ca celula initiala, iar cea mare va reprezenta celula segment(derivata). 2.1.2 Meristemele primare- sunt reprezentate de complexe de celule care prezinta deja un inceput de diferentiere exprimat si se divid intens furnizand materialul celular necesar formarii celulelor definitive.Dupa pozitia pe care o ocupa in corpul plantei meristemele primare sunt apicale si intercalare. Meristemele primare apicale sunt localizate la mica distanta de varful vegetativ meristematic al tulpinii sau al radacinii. Meristemele primare intercalare.Meristemul intercalar este un tesut cu crestere activa situat la o anumita distanta de varful vegetativ meristematic.Se intalneste la tulpinile articulate fiind situat in internoduri. 2.1.3 Meristemele secundare(laterale) sunt reprezentate de 2 tipuri de cambiu(cambiul vascular si cambiul suberofelodermic), cu dispozitie concentrica in corpul plantelor vasculare, cu activitate bifaciala, responsabile de cresterea secundara, in grosime a acestora.Cambiul vascular este un meristem lateral care se dezvolta la plantele vasculare cu cresterea secundara in grosime in interiorul cilindrului central.Cambiul suberofelodermic reprezinta cel de-al doilea meristem secundar al plantelor vasculare.Felogenul se formeaza in afara cambiului vascular, printr-un proces de remeristematizare a unor celule mai mult sau mai putin specializate. 2.2 Sistemul aparator . Tesuturile protectoare sunt localizate la exteriorul organelor plantelor si mai rar in interiorul acestora, avand rolul de a apara celelalte tesuturi pe care le delimiteaza.Dupa originea lor tesuturile se impart in : 1- tesuturi aparatoare primare (epiderma, exoderma, endorderma si tesutul caliptral); 2- secundare(suberul). 2.2.1 Tesuturi protectoare primare Epiderma(rizoderma) este tesutul principal de aparare al plantelor reprezentata prin stratul de celule care delimiteaza si protejeaza spre exterior organele vegetative si reproducatoare ale acestora.La radacina acest tesut se numeste rizoderma.Rolul protector al epidermei reiese din particularitatile celulelor epidermale.Variate ca forma, marime si dispozitie, acestea se pot grupa dupa functiile pe care le indeplinesc in 3 categorii:1- celule epidermale propriu-zise; 2- celule epidermale specializate care intra in alcatuirea stomatelor si perilor; 3- emergente.Celulele epidermale propriu-zise pot fi parenchimatice sau prosenchimatice, in functie de tipul de organ pe care il acopera.Celulele epidermice specializate sunt stomatele si perii.Stomatele sunt structuri care asigura respiratia sau transipratia celulelor din tesuturile sbepidermale.Se formeaza de obicei pe toate organele supraterane ale plantelor dar mai ales pe frunze.Perii sunt formatiuni de origine epidermala prezenti pe organele vegetative si reproducatoare.Dupa rolul pe care il indeplinesc in viata plantei se clasifica in peri cu rol protector si peri cu rol secretor.
Exoderma este un tesut primar protector caracteristic radacinilor, localizat sub rizoderma. Este formata din celule pe peretii usor ingrosati, suberificati uneori lignificati.Printre acestea se intercaleaza in dreptul perisorilor absorbanti, celulele de pasaj sau de trecere, cu peretii celulari subtiri , prin care circula apa absorbita de perisorii absorbanti, in drum spre cilindrul central. Endoderma este tesutul cel mai intern al scoartei primarem intalnit mai ales la radacina, mai rar la tulpina si frunza. La tulpinile supraterane aceasta structura nu se diferentiaza in mod deosebit ci se prezinta sub forma de teaca amilifera.La tulpinile subterane si la cele ale plantelor acvatice exista insa o endoderma tipica.Endoderma are rolul de a a apara tesuturile vii ale cilindrului central, de a mentine forma normala a acestuia si de a orienta circulatia apei cu sarurile minerale. Tesutul caliptral este caracteristic radacinilor avand rolul de a proteja varfurile vegetative meristematice ale acestora.La cele mai multe plante isi are originea in caliptrogen, este deci un tesut protector primar. 2.2.2. Tesuturi protectoare secundare . Suberul este un tesut de origine secundara rezultat din activitatea felogenului.Acest tesut este slab permeabil pentru apa si gaze si slab conducator de caldura.Este alcatuit din celule de obicei izodiametrice, tabluare sau paralelipipedice, cu dispozitie radiara, lipsite de spatii intercelulare.Suberul se diferentiaza la suprafata organelor vegetative persistente(radacini, tulpini, rizomi, tuberculi) ca tesut secundar de protectie, insa este prezent si la unele tipuri de fructe ca si in structura frunzelor protectoare ale mugurilor.Pe tulpini, radacini, si petioluri acoperite cu un strat de suber apar mici ridicaturi, formatiuni suberoase de origine secundara numite lenticele. 2.3.Sistemul tesuturilor conducatoare.Functia de transport este asigurata in corpul plantelor vasculare, de catre tesuturi speciale,numite conducatoare.Dupa originea lor tesuturile conducatoare sunt primare si secundare. 2.3.1 Tesutul conducator lemnos(lemnul, hadromul sau xilemul) Tesutul conducator lemnos sau xilemul, alcatuit din vase lemnoase, si fibre lemnoase este dupa originea sa primar si secundar.In structura xilemului primar se disting, dupa momentul aparitiei lor 2 categorii de elemente grupate sub denumirea de protoxilem sau metaxilem.Protoxilemul este constituit din primele elemente lemnoase care apar in viata plantei, diferentiate inaintea de terminarea cresterii in lungime a organului, iar metaxilemul din elementele formate ulterior de calibru mai mare, cu peretii mai puternic ingrosati si mai intens lignifiati. 2.3.2 Tesutul conducator liberian(liberul, leptomul sau floemul) Tesutul conducator liberian sau floemul este alcatuit din elemente perforate(celule si tuburi ciuruite), celule anexe(prezente numai la angiosperme), parenchim liberian si fibre liberiene.Dupa originea sa floemul este primar si secundar la fel ca si xilemul.In liberul primar se disting doua categorii de elemente, formate succesiv in ontogenia plantei: 1- protofloemul si 2- metefloemul; care impreuna alcatuiesc leptomul. 2.4Sistemul fundamental este alcatuit din tesuturi parenchimatice si ocupa volumul cel mai mare din corpul plantelor superioare.Aceste tesuturi iau nastere din meristemul fundamental si sunt alcatuite aproape exclusiv din celule vii.Cuprinde: 1- parenchimurile(asimilator, acvifer, aerifer, de inmagazinare a substantelor de rezerva); 2- tesuturile mecanice(colenchimul si sclerenchimul). 2.4.1.Perenchimurile.Tesuturile parenchimatice sunt prezente in aproape toate organele vegetative si reproducatoare ale plantelor intrand inclusiv in componenta tesuturilor conducatoare, sub forma de parenchim lemnos si liberian.Sunt considerate tesuturi cu o specializare relativ redusa, insa cu sarcini fiziologice importante. Parenchimul de absorbtie este format din tesuturi a caror functie principala este de absorbtie a apei si a substantelor minerale dizolvate in apa, solutii pe care le transmite prin vasele conducatoare.Parenchimul de absorbtie poate sa absoarba apa si sub forma de vapori din atmosfera umeda.Sunt si parenchimuri a caror functie este de a absorbi substante organice deja elaborate.In functie de aceste particularitati se deosebesc 3 tipuri de parenchimuri de absorbtie: rizoderma(cuprinde totalitatea perisorilor absorbanti impreuna cu celulele epidermice si parenchimul cortical implicate in absorbtia si transmiterea apei cu sarurile minerale spre tesuturile conducatoare lemnoase); velamen radicum (este tesutul radacinilor aeriene care indeplineste o dubla functie, de protectie si absorbtie) epiderma cotiledonara( indeplineste dubla functie de a secreta mai intai o enzima hidrolizanta, care transforma amidonul din endospermul semintei in zahar, pentru ca apoi sa-l absoarba si sa-l transmita embrionului in stare de dezvlotare). Parenchimul asimilator este unul dintre cele mai importante tesuturi fundamentale, fiind implicat in procesul de fotosinteza.Acest tesut este bine reprezentat in organele supraterane ale plantelor, in special in frunze si in unele tipuri de tulpini.Este alcatuit din celule vii cu peretii celulozici subtiri , localizate subepidermal, caracterizate prin prezenta a numeroase cloroplaste si a unui sistem vacuolar dezvlotat.Multe tipuri de celule din corpul plantelor sunt dotate cu cloroplaste(epidermale, corticale,
celulele radacinilor aerifere )si realizeaza sinteza de substante organice in prezenta luminii, dar nu pot fi considerate asimilatoare propriu-zise. Parenchimul aerifer caracterizeaza speciile de plante acvatice si palustre.Celulele acestui tesut lasa intre ele spatii mari sau lacune de origine lizigena care iau foema de canale mari ce strabat intregul corp al plantei.In aceste spatii se inmagazineaza mari cantitati de aer care determina micsorarea greutatii specifice a plantei respective, dand posibilitatea organelor vegetative si reproducatoare sa pluteasca sau se se mentina in pozitie verticala in apa. Parenchimul acvifer este un tesut specializat pentru depozitarea apei caracteristic plantelor suculente care vegeteaza in regiuni secetoase, plante xerofile si plantelor halofile care se dezvolta pe soluri saraturate.In frunzele plantelor xerofile parenchimul acvifer insoteste tesutul asimilator caruia ii cedeaza apa in perioadele de seceta. Parenchimul de depozitare a materiilor de rezerva . Asimilatia clorofiliana sau fotosinteza se produce in general cu atata energie in parenchimul asimilator incat produsele de sinteza asigura nu numai necesitatile curente ale intregului organism ci se si depun ca matriale de rezerva in tesuturile de depozitare.Tesuturile de depozitare sunt alcatuite din celule vii nucleate care isi pastreaza vitalitatea pe toata durata consumului lor.Parenchimul de depozitare a substantelor de rezerva cumuleaza totalitatea tesuturior in care se stocheaza materii de rezerva(glucide, proteine, lipide, cristale de oxalat de Ca).Materiile de rezerva se depoziteaza sub forma de substante ergastice in citoplasma, in aminoplaste, in vacuole mai rar chiar in peretii celulari(lupin, cafea, samburii de curmal). 2.4.2. Tesutul mecanic de sustinere.Cormofitele ierboase si lemnoase la inceputul dezvoltarii lor isi mentin propria greutate si rezista la actiunea factorilor extremi datorita turgescentei precum si a tesutului conducator lemnos, acestora adaugandu-se lor tesuturi speciale de sustinere colenchimul si sclerenchimul.Tesuturile mecanice sunt alcatuite din stereide.Tesuturile mecanice formeaza din punct de vedere functional stereomul.Pot fi primare, sunt formate din meristeme primare si secundare sau secundare(generate de cambii). Colenchimul alcatuit din celule vii este un tesut elastic si rezistent la intindere.Caractristic organelor vegetale tinere in curs de crestere.Se diferentiaza in tulpini, frunze , elemente florale, pe ricarpul fructelor si mai rar chiar in unele radacini.Dupa modul caracteristic de ingrosare a peretelui celular se deosebesc 3 tipuri de colenchim:1-colenchim angular(celule heterodiametrice strans unite intre ele, lipsite de spatii intercelulare ai caror pereti sunt ingrosati predominant in lungul muchiilor);2- colenchimul tabular(ingrosarea pronuntata a peretilor tangentiali externi/ interni si externi in timp ce peretii radiari raman subtiri dand forma de placa celulei); 3- colenchimul angular(celule de marimi si forme diferite cu spatii intercelulare in dreptul carora peretii sunt mai ingrosati ). Sclerenchimul tesut mecanic rezistent la tractiuni si presiuni alcatuit din celule specializate care l-a completa maturitate isi pierd vitalitatea; se gaseste in toate organele plantelor ; celulele acestui tesut au peretii uniform ingrosati si adesea lignificati si sunt lipsiti de spatii intercelulare.Sclerenchimul se imparte in 2 categorii: 1- sclereide(celule sclerenchimatice scurte) ; 2-fibre sclerenchimatice (celule sclerenchimatice lungi).Sclereidele sunt celule mai mult sau mai putin izodiametrice cu lumen redus, pereti celulari puternic ingrosati si strabatuti de puctuatiuni simple sau ramificate prin intermediul carora se mentin legaturile plasmatice dintre celulele vecine tinere.Fibrele sclerenchimatice sunt elemente unicelulare heterodiametrice in mod obisnuit foarte lungi, inguste ascutite la capete diferite prin grosimea si compozitia chimica a peretelui celular variate ca origine si dezvoltare. 2.5. Sistemul tesuturilor speciale.Tesuturile speciale cuprind tesuturile secretoare si tesuturile de separatie 2.5.1.Tesuturile secretoare .Unele celule produc ca rezultat al metabolismului multe substante care par a fi neuitilizabile in procesele vitale.Ele nu alcatuiesc intotdeauna un tesut unitar fiind reprezentate de celule singulare sau de grupuri de celule dispersate printre parechimuri.Celulele direct implicate in secretie isi mentin peretii celulari primari la numeroase mitocondrii, RERugos bine dezvoltat, dictiozomi abundenti, sistem vacuolar redus, plastide rare si slab functionale.Tesuturile secretoare se grupeaza in 2 categorii: tesuturi secretoare externe si interne. Tesuturile secretoare externe (papile secretoare, osmofore , peri secretori , hidatodele, glande saline, emergente secretoare, si nectariile). Tesuturi secretoare interne(cavitati secretoare, canale secretoare, ) 2.5.2Tesuturile de separatie asigura detasarea sau separarea unor tesuturi, complexe de tesuturi sau a unor organe vegetative si reproducatoare de restul corpului vegetativ al plantei.
Cap.3 Organografia 3.1 Radacina este organul dezvoltat in partea inferioara a plantei care creste ortotrop si se adanceste in pamant.Se deosebeste de alte organe subterane prin lipsa frunzelor si a mugurilor si o structura anatomica proprie.Are urmatoarele functii: - fixeaza planta in sol si sustine partile aeriene;- absoarbe apa si sarurile minerale;- depoziteaza substante de rezerva;- stabileste simbioze;- regenereaza uneori intreaga planta. 3.1.1.Morfologia radacinii are 4 zone :1- varf vegetativ; 2- zona neteda; 3- zona pilifera; 4- zona aspra.Varful vegetativ are forma conica, este ocupat de una sau mai multe celule initiale si de meristemele derivate din ele.Este zona in care celulele se inmultesc prin diviziune mitotica si de aceea are caracter meristematic.La exterior varful vegetativ este protejat de calpitra.Zona neteda corespunde zone de crestere in lungime a radacinii.Celule sale nu se mai divid dar cresc prin intindere marindu-si dimensiunea initiala si sporind astfel forta datorita careia radacina patrunde in sol. Zona pilifera (zona perisorilor sugatori ).Zona aspra este ultima zona a radacinii cuprinsa intre zona pilifera si colet. Tinand seama de raportul de marime dintre radacina principala si radicele se deosebesc 3 ripuri de de radacini: pivotante(la care radacina principala este bine dezvoltata avand forma de tarus); fasciculate(la care radacina principala se dezvolta foarte putin sau deloc fiind inlocuita cu radacini adventive subtiri si lungi care se formeaza din nodurile tulpinale); ramuroase(la care radicele de ordinul 1 au aprope aceleasi dimensiuni sau chiar o depasesc). 3.1.2.1 Structura primara a radacinii.Tesuturile din care este formata rezulta din activitatea meristemelor primare apicale.Radacinile cu structura primara sunt alcatuite din tesuturi definitive(aparatoare, conducatoare si parenchimatice si mecanice) dispuse concentric in 3 zone caracteristice: rizoderma, scoarta si cilindrul central.Rizoderma este tesutul protector extern, alcatuit din celule parenchimatice vii strans unite intre ele cu pereti subtiri, celulozici, nemodificati secundar.LA nivelul zonei pilifere majoritatea celulelor sunt transformate in perisori sugatori. Scoarta are 3 zone concentrice (exoderma, scoarta mijlocie si endoderma).Cilindrul central este format din tesuturi conducatoare(floem si xilem) care formeaza fascicule separate lemnoase si liberiene dispuse in alternanta intr-un parenchim fundamental si tesuturi neconducatoare.La exterior este delimitat de un tesut numit periciclu alcatuit din celule parenchimatice mici care alterneaza cu celulele endodermei.Sub periciclu se formeaza proxilemul alcatuit din vase cu lumen mic, cu pereti celulari partial ingrosati secundar si metaxilemul catre meristemul medular, alcatuit din vase lemnoase cu lumen mare. 3.1.2.2.Structura secundara a radacinii La majoritatea plantelor radacina prezinta o structura secundara care determina cresterea in grosime a radacinii.Meristemele secundare formeaza tesuturi noi(protectoare, conducatoare, mecanice si parenchimuri) care se alatura celor primare devenind cu timpul predominanante.Cambiul vascular apare sub forma unor arcuri generatoare concave localizate in partea inferioara a fasciculelor liberiene si convexe dispuse la partea superioara a fasciculelor lemnoase.Acesta furnizeaza tesuturi conducatoare in dreptul fiecarui tesut conducator.Xilemul secundar este alcatuit din vase lemnoase cu diametru mare, solidarizate intre ele prin celule de parenchim lemnos si fibre lemnoase.Liberul secundar este in general alcatuit ca se cel primar cu precizarea ca acesta are lumenul mai mare si este insotit de fibre liberiene.Felogenul este meristemul secundar ce asigura cresterea in grosime a plantei prin diferentiere la exterior in suber iar la interior in feloderm.Alternanta zonelor subero-felodermice primare si secundare duc la aparitia ritidomului. 3.1.3 Metamorfozele radacinii sunt radacinile care indeplindesc alte functii pe langa cele de fixare si absorbtie. Radacini tuberizate-functie predominant de depozitare a substantelor de rezerva.Tuberizarea poate afecta uneori toata radacina principala si axa hipocotila schimbandu-si treptat forma, marimea si culoarea.In alte cazuri tuberizarea afecteaza doar partea superioara a radacinii superioare si axa hipocotila.TUberizarea mai poate afecta atat partea superioara a radacinii pivotante si a hipocotilului cat si o zona din epicotil. Radacini contractile al unele plante perene cu organe hibernante(bulb, rizom, tuberculi) si muguri de regenerare in sol se manifesta fenomenul de heterorizie. Ele formeaza pe langa radacinile normale si radacini modificate, contractile , capabile sa se scurteze dupa terminarea cresterii in lungime si sa se adanceasca toamna in sol. Radacini inmagazinatoare de apa caracteristice plantelor din regiunile aride, au radacini aeriene capabile sa absoarba apa din atmosfera prin intermediul unui parenchim special numit velamen radicum localizat sub rizoderma.
Radacini proptitoare sunt radacini adventive care se formeaza la nivelul nodurilor tulpinale si rameale.Ajungand in sol se ingroasa se ramifica si constituie un sistem eficient de sustinere rezistent la presiune si tractiune. Radacini cu nodozitati pe radacinile plantelor leguminoase cat si pe cele ale unor arbori.Se formeaza excrecente de forme marimi si culori diferite denumite nodozitati.Ele sunt rezultatul existentei relatiei de simbioza intre organismele fixatoare de azot si planta. Radacini sugatoare (hausotori) sunt organe speciale de absorbtie de origine endogena care se diferentiaza pe tulpinile plantelor parazite si semi-parazite,plante la care radacina este prezenta doar in stadiul de plantula.Cu ajutorul haustorilor absorb apa si sarurile minerale sau seva elaborata din vasele lemnoase, respectiv cele liberiene ale plantei parazitate. Radacini cu pneumatofori radacini ale unor plante din regiunile tropicale si padurilor mlastinoase de mangrove si-au format niste structuri speciale denumite pneutomatofori, care permit accesul aerului atmosferic catre radacinile afundate in namol. Radacini hidromorfe apar la plantele acvatice si se caracterizeaza prin: lipsa perisorilor absorbanti si exodermei; prezenta unui sistem cortical de canale aerifere mari; reducerea elementelor mecanice si xilemice din structura cilindrului central. 3.2 Tulpina este un organ vegetativ axial cu geotropism negativ mai rar plageotropa sau chiar pendula.Este adaptata morfologic si structural pentru indeplinirea a 2 functii : de sustinere (a frunzelor , florilor si ramurilor )si de conducere (a sevei brute si a sevei elaborate).unele tulpini pot contrubui in unele cazuri la nutritia plantei prin fotosinteza la inmagazinarea apei si a substantelor de rezerva sau pot functiona ca organ de multiplicare vegetativa. 3.2.1 Morfologia tulpinei Tulpina este alcatuita dintr-o axa principala, care poate sa ramana simpla sau poate sa se ramifice.In ultimul caz se formeaza ramuri de ordine diferite (primare, secundare. tertiare).In partea terminala a tulpinii este localizat meristemul apical caulinar.Spre baza varfului vegetativ se formeaza primordiile organelor laterale care au in primele stadii de dezvoltare aspectul unor mici excrescente.Din altele , numite primordii foliare se vor forma frunzele iar din altele situate la subosoara celulelelor foliare se vor diferentia ramurile.Tulpina este de cele mai multe ori un organ articulat, format din noduri si internoduri.Zonele putin umflate care se observa in lungul unei tulpini articulate la nivelul carora se diferentiaza frunzele se numesc noduri, iar portiunile de tulpina cuprinse intre 2 noduri se numesc internoduri.Tulpina este de cele mai multe ori un organ articulat de care se formeaza muguri atat la nivelul nodurilor cat si la partea apicala.Mugurele este considerat un lastar scurt constituit dint-o axa cu noduri foarte apropiate de pe care se diferentiaza primordiile foliare ce se incovoaie peste meristemul apical pe care il acopera si il protejeaza.Mugurii sunt de regula protejati de niste frunze mici, solzoase numite catafile.Atunci cand frunzele protectoare lipsesc mugurii se numesc nuzi.Dupa pozitie mugurii se grupeaza in: terminali, axiali si aventivi.Dupa felul organelor ce se formeaza din muguri acestea se impart in : foliari , florali si mixti.La tulpina ca si la radacina se deosebesc 2 tipuri principale de ramificatie: dihotomica si monopodiala. IN ramificatia dihotomica formarea mugurelui este terminala pentru ca ea depinde de o singura celula initiala localizata in varful vegetativ caulinar care se divide in 2 celule egale care vor da nastere la cate un mugure respectiv o ramura.In ramificatia monopodiala formarea ramurilor este subterminala.In functie de comportamentul meristemului terminal al axei principale se disting 2 subtipuri: ramificatia racemoasa si ramificatia cimoasa.La ramificatia racemoasa meristemul apical al axei principale ramane activ toata perioada de viata a plantei asigurand alungirea continua a tulpinei prin mugurele sau terminal ramurile laterale care se dezvolta centripetal nu depasesc in lungime axa principala. 3.2.2. Anatomia tulpinii Structura primara ia nastere din activitatea meristemelor primare apicale. Este completata si la unele plante de activitatea meristemelor intercalare si caracterizeaza tulpina tuturor cormofitelor tinere.Ea se mentine pe tot parcursul vietii la majoritatea plantelor monocotiledonate si la unele dicotiledonate erbacee. 3.2.2.1. Structura primara a tulpinii cuprinde 3 zone histologice concentrice: epiderma, scoarta si cilindrul central. Epiderma tesut protector, acopra si delimiteaza tesuturile tulpinii in raport cu exteriorul.Este de regula unistratificata alcatuita din celule vii strans unite intre ele, lipsite de spatii intercelulare si acoperita de cuticula.La plantele tinere si erbacee celule epidermale prezinta de asemenea cloroplaste participand la procesul de fotosinteza.Epiderma tulpinara prezinta stomate, peri protectori si sau protectori foarte diferiti ca forma si produsi de secretie.Scoarta mai ingusta decat cea a radacinii este alcatuita din cateva straturi de celule in general parenchimatice.Cele externe pot fi implicate in fotosinteza pe cand cele interne
reprezinta spatii de depozitare intre diferite categorii de substante de rezerva.Scoarta primara este alcatuita cu zone continue sau fragmentare de tesut mecanic(colenchim si sclerenchim).Endoderma tipica prezinta doar la tulpinile subterane si la cele ale plantelor acvatice.La tulpinile supraterane stratul cel mai intern al scoartei se transforma intr-o zona de depozitare a amidonului denumit in consecinta teaca amilifera.Centrul tulpinii este ocupat de o coloana intacta de tesuturi conducatoare si neconducatoare care formeaza cilindrul central sau stelul.Tulpinile cormofitelor se impart in 3 grupe in functie de nr stelilor: monostelice, polistelice si astelice.Dupa pozitia tesuturilor in interiorul cilindrului central se deosebesc urmatoarele tipuri de stel: protostel, actinostel si plectostel, sifonostel, eustel, atactostel. 3.2.2.2.Structura secundara a tulpinii Cresterea in grosime se realizeaza pe baza meristemelor secundare reprezentate de cambiul vascular si felogen.Zona generatoare libero-lemnoasa este localizata in interiorul cilindrului central si isi are originea in segmentele de cambiu intra si inter fasciculare.Din activitatea sa bifaciala se formeaza catre exterior elementele liberului secundar iar spre interior elementele xilemului secundar.Liberul secundar este alcatuit din tuburi ciuruite, celule anexe, parenchim liberian si elemente mecanice de tip sclerencimatic reprezentate de fibre liberiene.La sfarsitul fiecarei perioade de vegetatie elementele liberiene se gloesc de continut si cu exceptia fibrelor sunt ulterior strivite prin efectul mecanic indus de formare a noilor structuri secundare.Din activitatea cambiului vascular se formeaza si razele medulare secundare care strabat lemnul si liberul secundar, servind la conducerea substantelor nutritive in sens radiar.Functionarea periodica a cambiului vascular genereaza noi structuri concentrice de tesuturi conducatoare secundare care se organizeaza sub forma a cate unui manson de liber secundar.Lemnul de primavara este alcatuit din vase de lemn mare care faciliteaza circulatia apei si a sarurilor minerale in timp ce lemnul de toamna este constituit din vase cu lemn mult mai mic si de numeroase fibre lemnoase.La esentele tari lemnul secundar se diferentiaza in 2 zone distincte:1- zone externa(functionala de culoare alba sau galbuie numita albur); 2- zona interna (nefunctionala lipsita de vitalitate, de culaoare intunecata,cu rol mecanic numita inima lemnului sau duramen).Felogenul functioneaza bifacial ca si cambiul vascular.El produce spre exterior suber iar spre interior feloderm.Are insa o activitate de scurta durata anuala fiind periodic inlocuit de o noua zona generatoare suberofelodermica localizata tot mai profund in corpul plantei.Se formeaza astfel scoarta secundara care este echivalenta de unii autori cu periderma.Din suprapunerea mai multor periderme care si-au pierdut vitalitatea in decursul anilor se formeaza un tesut mixt cu rol de aparare. 3.2.3.Tulpini metamorfozate Metamorfozele afecteaza tulpinile supraterane, subterane si acvatice.Tulpinile supraterane se pot modifica indeplinind functia de absorbtie inlocuind sistemul foliar care in general se reduce sau prin tuberizare cand devin organe de depozitare pt substantele de rezerva. Tulpini asimilatoare sunt reprezentate de tulpinile carnoase ale plantelor suculente cu frunze mai mult sau mai putin reduse, uneori solziforme sau transformate in spini.Din punct de vedere structural predomina tesutul acvifer alcatuit din celule capablie sa retina apa datorita unor substante mucilaginoase pe care le contine. Tulpini tuberizate au rol in inmultirea vegetativa a plantelor.Se pot tuberiza mai multe noduri de la baza tulpinii(gulie) axa mugurilor foliari(tuberule) frunzulitele mugurelui foliar(bulbiile).Tulpinile subterane sunt caracteristice plantelor erbacee perene si se pot confunda cu radacina.Tulpinile subterane(rizomii , tuberculii, bulbii) au rol in depozitarea substantelor de rezerva si in supravietuirea pe durata a iernii.Rizomii sun tulpini alcatuite din internoduri (scurte sau lungi) pe care se formeaza muguri axiali sau terminali pe care se vor dezvolta tulpini supraterane florifere.Tuberculii sunt rezultatul tuberizarii partiale sau totale ale ramurilor subterane.Pe tubercul in axila unor frunze rudimentare se diferentiaza mugurii axiali(ochi) din care vor iesi tulpini supraterane.Bulbii sunt tulpini subterane scurte acpoerite de frunze carnoase modificate in care se depun substante de rezerva, tulpina e scurta si latita in forma de disc.Din mugurele terminal in fiecare an se va dezvolta o tulpina supraterana iar din mugurii axiali noi bulbi(bulbi de inlocuire).Dupa modul de inserare a frunzelor pe bulb se disting:1- bulbi tunicati;2- bulbi scvamosi. Tulpinile plantelor acvatice Plantele acvatice constituie o categorie de plante caracterizate printr-o serie de trasturi specifice. Hidrofitele pot fi submerse sau amfibie.Tulpinile acvatice au in structura lor tesut aerifer bine dezvoltat, tesutul cortical este strabatut de numeroase canale aerifere.Tesutul conducator lemnos este slab dezvoltat ca si cel mecanic in predominant la plantele submerse. 3.3.1.Frunza se dezvolta din primordiile foliare ca o expansiune laterala a tulpinii cu crestere definitiva formand impreuna cu aceasta lastarul.Este organul cu cea mai mare plasticitate morfostructurala si reprezinta un criteriu important in identificarea speciilor de plante.Frunzele platelor sunt direct implicate in
procesele de fotosinteza, respiratie si trasnpiratie.In afara de aceste functii principale unele frunze metamorfozate indeplindesc si alte functii: de absorbtie de depozitare a unor substante, sau implicate in nutritia plantelor carnivore sau servesc ca organe de inmultire vegetativa. 3.3.2. Dispozita frunzelor pe tulpina Modul de asezare a frunzelor pe tulpina (filotaxia) se manifesta inca din mugure si este determinata de forma si dezvoltarea primordiilor foliare in interiorul acestora, de spatiere a lor, de distanta fata de varful vegetativ precum si de o serie de factori externi cum ar fi gravitatia si lumina.Se deosebesc 3 tipuri de dispozitie foliara: alterna(in spirala si se caracterizeaza prin cresterea unei singure frunze la fiecare nod), verticilara(3 sau mai multe frunze la fiecare nod) si opusa(2 frunze la fiecare nod; astfel incat frunzele de la nodul superior sa nu se suprapuna peste cele localizate la nodul inferior ci sa ocupe spatiul liber delimitat de frunzele de la nodul precedent). 3.3.3.Succesiunea frunzelor pe tulpina se deosebesc 4 categorii de frunze: cotiledoane(sunt primele frunze care iau nastere in viata unei plante), catafile(servesc de regula ca organ de aparare si mai rar de depozitare fiind prezente la muguri si tulpini subterane), nomofile(sunt frunze tipice complet dezvoltate intotdeauna verzi, asimilatoare, alcatuite din limb petiol si teaca. au rol in fotosinteza ,respiratie si transpiratie), hipsofilele(sunt ultimele organe foliare care se formeaza pe tulpina si au rol de protectie pt flori si inflorescente). 3.3.4. Morfologia frunzei Nomofilele, au 3 parti componente: limb petiol si teaca. Limbul foliar este partea principala a frunzei latita si de reguila turtita intr-un singur plan.Se disting 2 tipuri de frunze : simple si compuse.Frunzele simple au lamina formata intr-o singura parte si pot avea forma: eliptica, ovala, lanceolata, orbiculara, reniforma, triunghiulara, romboidala, deltoita, cordiforma, sagitala, hastata, spatulata , cuneata , lineala, ensiforma, subulata, aciculata, setacee, cilindrica.Frunzele cu marginea usor incizata pot fi :sinuate cu contur sinuos, serate, dintate, crenate.Frunzele cu marginea adanc incizata pot fi: palmate(palmat lobate, palmat fidate, palmat partite, palmat sectate) sau penate.In cazul frunzelor simple cu lamina intraga sau prevazuta cu incizii mici baza limbului prezinta la diferite plante forme variate: rotunda , cordata, sagitala , hastata, reniforma, cuncata, atenuata in petiol, auriculata.Varful limbului prezinta de asemenea diferite forme: ascutit, acumiat, rotund, obtuz, triunghiat, emarginat, obcordat, cuspicat, spinos.Frunzele simple lipsite de petiol se numesc sesile si exista mai multe tipuri: amplexicaule, perfoliate, decurent, conate.Spre deosebire de frunzele simple frunzele compuse au lamina divizata in parti distincte denumite foliole, inserate pe o axa comuna numita rahis.Dupa dimensiunea rahisului si dupa modul de dispunere al folioleleor se deosebesc 2 tipuri de frunze compuse : palmat compuse si penat compuse.Primele au rahisul scurt, situat la extremitatea apicala a petiolului si lamina alcatuita din 3 sau mai multe foliole dispuse in evantai.A 2 a categorie au rahisul lung iar lamina este formata din numeroase foliole dispuse simetric de o parte si de alta a rahisului.Se impart in 2 categorii: imparipenat compuse si paripenat compuse.Limbul frunzelor difera si prin modul de dispunere al nervurilor(sunt reprezentate de fascicule conducatoare care se continua din tulpina prin petiol in limbul foliar).Dupa grosimea si pozitia lor se impart in mediane principale, secundare, tertiare.Nervatiunea penata se caracterizeaza prin existenta unei singure nervuri mediane, groase, evidente care strabat limbul in tot lungul sau si de pe care se desprind nervuri secundare ce se ramifica ulterior, cu formare de nervuri tertiare.Nervatiunea palmata se caracterizeaza prin dezvoltarea aproape egala a nervurilor laterale comparativ cu nervura principala si prin faptul ca pornesc aproape toate printrun punct situat la baza limbului.Nervatiunea paralela: nervatiunile sunt paralele, se unesc la varful limbului , iar intre nervuri se formeaza anastamoze perpendiculare. Petiolul sustine si leaga limbul de tulpina, il orienteaza fata de lumina, si asigura legaturile vasculare cu tulpina.Este fixat de obicei la baza limbului.Poate avea diferite forme: cilindric,aripat, turtit si turtit dorsiventral. Baza frunzei este portiunea prin care aceasta se racordeaza la tulpina sau ramura.Frunzele sesile se fixeaza pe tulpina in baza laminei iar cele petiolate cu extremitatea inferioara a petiolului(mai dilatata si reprezinta baza frunzei). Anexele foliare: Stipelele sunt excrescente ale bazei frunzei de regula perechi de diferite forme, marimi, consistenta si durata.La baza frunzelor mai tinere se diferentiaza cate 2 stipele mici, membranoase, la inceput verzi, implicate in fotosinteza.Ulterior, odata cu maturarea frunzelor ele se usuca si cad(stipele caduce).Stipele inguste si concrescute cu baza petiolului se intalnesc la unele plante erbacee si lemnoase. Ochrea provine din concresterea stipelelor in forma de cornet sau palnie de consistenta membranoasa , care inveleste si protejeaza baza internodului si mugurele foliar. Ligula se prezinta ca o expansiune membranoasa continuand teaca de la articulatia sa cu limbul.Are
rolul de a impiedica apa sa intre intre teaca si petiol.Dupa forma ei ligula poate fi : scurta si dintata; lunaga si membranoasa, paroasa. 3.3.5.1. Structura limbului .Frunza la spermatofite.Frunzele gimnospermelor sunt bifaciale si prezinta diferite particularitati structurale determinate de conditiile in care traiesc plantele respective. La gimnosperme se intalnesc 3 tipuri structurale de frunze: dorsiventrala, invers dorsiventrala si omogena.Structura bifaciala dorsiventrala este prezenta la frunzele aciculare de brad. Celule epidermale sunt mici cu pereti puternic si uniform ingrosati.Peretii externi sunt cutinizati si cerificati.Sub epiderma se afla hipoderma, discontinua si sclerenchimatica.Tesutul palisadic este format din 2-3 straturi de celule heterodiametrice, bogate in cloroplaste, situate sub epiderma superioara.Tesutul lacunar dispus sub epiderma inferioara este alcatuit din celule izodiametrice intre care se formeaza lacune de dimensiuni variate.IN mezofil se gasesc 2 canale rezimiforme mari, in apropierea fetei dorsale.In partea centrala a laminei este localizata nervura delimitata de o endoderma tipica.Cele 2 fascicule conducatoare libero lemnoase din structura nervurii sunt inconjurate de un tesut de transfuzie alcatuit din celule parenchimatice si traheide cu punctuatiuni areolate. 3.3.3.6 Metamorfozele frunzei In anumite conditii frunzele sufera o serie de modificari morfo-structurale induse de necesitatea indeplinirii unor functii speciale(rol protector de agatare nutritiv).Frunzele metamorfozate prezinta deseori caractere morfologice cu totul diferite fata de cele normale asimilatoare aparand ca organe noi, cum sunt carceii, spinii, urnele, capcanele. Carceii sunt adaptati pentru agatarea plantei de suport.Se poate transforma intreaga frunza in carcel, numai foliolele de la varf sau numai petiolul. Spinii.Se pot metamorfoza frunzele, stipele sau numai lobii sau dintii in spini. Filodiile provin din modificarea petiolului care creste in volum, se aplatizeaza si isi dezvolta un tesut clorofilian cu rol de asimilatie. Cele mai multe variatiuni si structuri foliare se intalnesc la plantele cu nutritie heterotrofa complementara, caracteristica plantelor carnivore. -frunzele superioare, cu varful incovoiat prezinta peri glandulari, lipiciosi, si mobili. -transformarea totala a frunzelor in capcane sau partiala : urne foliare si urne cu capacel. La uele plante frunzele sunt atat de reduse incat se recunoaste cu greu natura lor foliara.Aceste plante se numesc afile si sunt intalnite in regiunile aride.La plantele acvatice sau semi-acvatice se constata dezvoltarea pronuntata a sistemului foliar in raport cu restul corpului(frunzele pot fi: subtiri si transparente, tubulare, sectate cu segmente filiforme, in forma de benzi) Cap.4 Inmultirea plantelor 4.1.Inmultirea asexuata Monogamia se caracterizeaza prin aceea ca indivizii noi iau nastere plecand de la o celula sau un grup de celule apartinand aceluias organism, fara sa fi avut loc un proces sexual.Se diferentiaza 2 forme de inmultire: asexuata si vegetativa.Multiplicarea vegtativa a fost semnalata la multe grupe de cormofite si talofite.Printre modalitatile de multiplicare vegetativa se numara bipartitia indivizilor unicelulari, fragmentarea indivizilor pluricelulari, sporularea si formarea unor organisme specializate pentru acest scop. Bipartitia este cel mai simplu mod de reproducere asexuata si este frecventa la plantele unicelulare.Bipartitia indivizilor pluricelulari este caracteristica talofitelor pluricelulare cat si cormofitelor.Sporii sunt implicati in reproducerea vegetativa a multor grupe de alge, dar cel mai adesea, acestia fac parte dintr-un ciclu de viata complet, iar aparitia lor alterneaza cu procesele sexuale complexe.Sunt foarte raspanditi la talofite si variati ca morfologie si structura: 1- zoosporii se pot forma dintr-o celula oarecare a corpului sau dintr-un organ specializat numit sporange; 2- gonidiile se transforma intr-o celula vegetativa iar sporii sunt imobili; 3- tetraspori. Plantele superioare prezinta organe specializate de multiplicare vegetativa, cu morfologie si structura specifica procesului de propagare.In aceasta categorie fac parte : stolonii, drajonii, tuberculii si mugurii tuberizati(tuberule, bulbiile, si turionii). 4.2.Inmultirea sexuata presupune contopirea a 2 gameti diferiti rezultand un zigot . Celulele care se contopesc se numesc gameti, iar celule care le produc se numesc gametociti(sunt produsi de gametocist).In gametocistul masculin(anteridie) se diferentiaza gametii barbatesti (numiti anterozoizi) iar in gametul feminin (oogon) se formeaza gametii femeiesti(numiti oosfere).Luand in considerare caracteristicile gametilor(forma, dimensiuni, mobilitate) care participa la fecundatie, reproducerea sexuata se poate realiza printr-un proces de izogamie, anizogamie sau oogamie. In urma procesului de fecundatie, rezulta o celula noua, diploida , denumita zigot.Meioza si fecundatia reprezinta cele 2 procese care marcheaza ciclul de viata al unui organism in general si se imparte in 2
perioade: 1- generatia sporofitica (diploida) care incepe cu fecundatia si se sfarseste cu meioza; 2 generatia gametofitica(haploida) initiata prin meioza si incheiata prin fecundatie. Inflorescente Florile angiospermelor pot fi solitare sau grupate in inflorescente.Sunt cazuri cand tulpina poarta o singura floare apicala.La cele mai multe specii, florile sunt dispuse in mod grupat, pe ramificatiile lastarului florifer, formand inflorescente, simple si compuse.Inflorescentele simple se impart in 2 mari categorii: 1- monopodiale si 2- simpodiale. 1-Inflorescentele monopodiale se caracterizeaza printr-o axa principala unitara de pe care se diferentiaza axe laterale secundare, purtatoare de flori.Tipuri de inflorescente monopodiale: racemul(este alcatuit dintr-o axa principala, de pe care se dezvolta altern, in axila a cate unei bracte, flori pedicelate; pedicelii florali au aproape aceeasi lungime); spicul(este inflorescente derivata din racem cu flori sesile, are axa principala flexibila, pendenta, cu flori unisexuate si se numeste ament); corimbul(este format din flori diferentiate in lungul axei principale cu pedunculi de lungimi diferite, care ajung la acelasi nivel); umbela (este alcatuita din flori ce se diferentiaza de la noduri foarte apropiate, opeducelii florili la fel de lungi care ajung la acelasi nivel ); spadixul (este o inflorescenta cu axa principala ingrosata pe care se diferentiaza flori sesile, inflorescenta este protejata de o bractee mare, numita spata fie de mai multe bractei numite panusi) ; capitulul (prezinta axa principala globuloasa sau disciforma cu flori sesile sau aproape sesile); calatidiul (este o inflorescenta apropiata de capitul, de care se deosebeste prin aca principala mult latuita, prezenta involucrului); 2- Inflorescente simpodiale-axa principala nu este unitara ci este alcatuita dintr-o succesiune de axe de ordine si varste diferite, care se termina fiecare cu cate o floare.Dezvoltarea florilor se face bazipetal, de la varful inflorescentei spre baza acesteia sau centrifugal, de la centru spre periferia acestuia.Aceste inflorescente se impart in 3 grupe: monohaziu(prezinta o axa principala formata dintr-o succesiune de axe florifere asezate cap la cap si diferentiate unilateral); dihaziu(axa principala se termina cu cate o floare cea mai varstnica ); polihaziu (este alcatuit dintr-o axa principala, care poarta sub apical mai multe axe secundare de varste diferite). Alcatuirea florii Floarea angiosperma grupeaza elemente specializate, adaptate pentru protectia gametilor si a embrionului, in raport cu cele mai variate conditii de mediu.Floarea completa este alcatuita din 4 straturi de elemente: sepale, petale, stamina si carpene.Portiunea terminala a acesteia reprezinta de fapt pedunculul floral.Partea sa apicala , adesea umflata , ingrosata sau latita, formeaza axa florii numita receptacul pe care se diferentiaza elementele florale. Invelisul floral Stratul extern de elemente florale reprezentat de sepale formeaza caliciul.Urmatorul rand de elemente florale este reprezentat de frunze modificate in general modificate numite petale ce formeaza corola.Petalele pot fi libere sau concrescute.Sunt si cazuri in care corola lipseste iar astfel de flori se numesc apetale.Sepalele si petalele nu sunt direct implicate in procesul de reproducere.Ele reprezinta partile sterile ale florii, care joaca un rol important in polenizare , prin coloritul lor viu , si prin nectarul produs de glandele nectarifere.Androceul este reprezentat de totalitatea staminelor localizate spre interiorul florii si reprezinta partea masculina a florii. Fiecare stamina este alcatuita dintr-o parte sterila, reprezentata printr-un filament subtire lung si cilindric si o parte fertila latita, numita antera, alcatuita din 2 jumatati simetrice denumite tece.Filamentul se prelungeste la capat printr-o portiune de tesut steril denumit conectiv.Gineceul este organul feminin de reproducere sexuata format din totalitatea carpelelor localizate in interiorul florii.Carpelele sunt frunze modificate, ca si staminele, care insa si-au pastrat mai mult din aspectul foliar. Rolul gineceului este de a produce ovule. Gineceul este format din ovar(produce ovule, stil, si stigmat). Samanta Dupa fecundatie micropilul ovulului se inchide si sfarsitul procesului sexual este marcat de vestejirea, uscarea si in general caderea caliciului si a corolei.Ovarul continua sa se dezvolte , formanad fructul in timp ce ovulele vor deveni seminte.Samanta este un complex tisular alcatuit din tegument seminal protector, embrion si rezervele nutritive ale acestuia.Tegumentul seminal provine din dezvoltarea invelisului ovular si este format din 2 straturi, unul extern si unul intern.Embrionul este alcatuit din radicula, hipocotil, si unul sau 2 cotiledoane, intre care este localizat mugurasul, insotit de primele frunze.Tesuturile nutritive ale plantei sunt reprezentate de endospermul secundar si perisperm.Substantele nutritive vor fi mobilizate in momentul germinarii semnintelor, furnizand energie necesara cresterii si diferentierii ulterioare a embrionului. Fructul este o formatiune caracteristica angiospermelor si rezulta dupa fecundatie din gineceu . Pericarpul(peretele fructului ) ia nastere din peretele ovarului, si in functie de consistenta sa se disting 4 tipuri ecologice de fructe: capsula, nuca , baca, si drupa.
S-ar putea să vă placă și
- TOT CE TREBUIE SĂ ȘTII CA SĂ FII AS LA BIOLOGIE ÎNTR-UN DITAMAI CAIETULDe la EverandTOT CE TREBUIE SĂ ȘTII CA SĂ FII AS LA BIOLOGIE ÎNTR-UN DITAMAI CAIETULEvaluare: 5 din 5 stele5/5 (3)
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- Manual BiologieDocument25 paginiManual BiologieteomalancuÎncă nu există evaluări
- Anatomie - Curs1 - CelulaDocument27 paginiAnatomie - Curs1 - Celulaalfa5017Încă nu există evaluări
- BioDocument23 paginiBioSilvia Silviutza100% (1)
- Călătorie În Interiorul Unei Celule EucarioteDocument6 paginiCălătorie În Interiorul Unei Celule EucarioteAleee 06100% (1)
- Celula Animala Si Celula VegetalaDocument4 paginiCelula Animala Si Celula VegetalaBogdan Alban ValentinÎncă nu există evaluări
- Curs 1 BotanicaDocument58 paginiCurs 1 BotanicaAndreea Cocarcea100% (1)
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- Biologie Celulara19.05.2022Document64 paginiBiologie Celulara19.05.2022Pop MariaÎncă nu există evaluări
- Celula Si Organitele CelulareDocument4 paginiCelula Si Organitele CelulareMonica AoaneiÎncă nu există evaluări
- Anatomie Si Fiziologi1Document5 paginiAnatomie Si Fiziologi1Ion PahomiÎncă nu există evaluări
- L 2 Celula Structura ProprietatiDocument10 paginiL 2 Celula Structura ProprietatirahatnikitaÎncă nu există evaluări
- Celula EucariotaDocument3 paginiCelula EucariotaAlex Aurică100% (2)
- Curs II CelulaDocument44 paginiCurs II CelulaMaria-Luminita ChiritaÎncă nu există evaluări
- CELULADocument10 paginiCELULAIoana-Camelia OlteanuÎncă nu există evaluări
- Anatomie Curs 1Document16 paginiAnatomie Curs 1Danut Ancuța MihaelaÎncă nu există evaluări
- Biocel Cursuri WordDocument38 paginiBiocel Cursuri Wordbubu_lina_ruza100% (2)
- 3.3. Structura Celulei de DrojdieDocument4 pagini3.3. Structura Celulei de DrojdieVasile SîrbuÎncă nu există evaluări
- Celula Și Organitele CelulareDocument10 paginiCelula Și Organitele CelulareTurlea FloareaÎncă nu există evaluări
- Teza BiologieDocument9 paginiTeza BiologieAndrei IliciÎncă nu există evaluări
- Bac Bio 9-10Document78 paginiBac Bio 9-10Denisa Badoi100% (1)
- Celula (Structură, Proprietăţi)Document37 paginiCelula (Structură, Proprietăţi)Sorina DobreÎncă nu există evaluări
- Sinteza IX Sem IDocument13 paginiSinteza IX Sem IBogdan BogdanÎncă nu există evaluări
- Celula Animala vs. VegetalaDocument14 paginiCelula Animala vs. VegetalaCristina Gabriela100% (1)
- Organizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternDocument4 paginiOrganizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternVata LuminitaÎncă nu există evaluări
- Cito PlasmaDocument19 paginiCito PlasmaAlexandra Vidz100% (1)
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Rezumat Curs BotanicaDocument34 paginiRezumat Curs BotanicaispasÎncă nu există evaluări
- Notiuni Biologie Clasa A VIII-aDocument5 paginiNotiuni Biologie Clasa A VIII-aDmitrii BudaiÎncă nu există evaluări
- CelulaDocument5 paginiCelulaGeorge BrinzeaÎncă nu există evaluări
- Rolul Fiziologic Al Componentelor CelulareDocument9 paginiRolul Fiziologic Al Componentelor CelulareNovac IulianÎncă nu există evaluări
- CELULADocument7 paginiCELULAElisei CostinÎncă nu există evaluări
- Bac 2014Document27 paginiBac 2014Tudorica Petronela0% (1)
- 726 1259312854Document2 pagini726 1259312854Buzatu MadalinaÎncă nu există evaluări
- Cap 3Document8 paginiCap 3Marfescu CristianÎncă nu există evaluări
- Citologia VegetalaDocument5 paginiCitologia VegetalaGabura TaniaÎncă nu există evaluări
- BiologieDocument8 paginiBiologieRăcilă VasileÎncă nu există evaluări
- Celula Animala vs. Celula VegetalaDocument13 paginiCelula Animala vs. Celula VegetalaCristina GabrielaÎncă nu există evaluări
- Curs 2 BiocelDocument39 paginiCurs 2 BiocelNicoleta NikoÎncă nu există evaluări
- Curs BiocelDocument75 paginiCurs BiocelLavinia BragaÎncă nu există evaluări
- MicrobiologiaDocument41 paginiMicrobiologiaDenis PirauÎncă nu există evaluări
- Biologiei Celulare Si MoleculareDocument33 paginiBiologiei Celulare Si MoleculareI IiÎncă nu există evaluări
- 2.1. Celula: Celula Reprezintă Unitatea Morfo-Funcțională Și Genetică A OrganizăriiDocument6 pagini2.1. Celula: Celula Reprezintă Unitatea Morfo-Funcțională Și Genetică A OrganizăriiMedeea GabrielaÎncă nu există evaluări
- Celula - PrezentareDocument23 paginiCelula - PrezentareIulia Manuela Plesa100% (1)
- Celula Curs 2Document12 paginiCelula Curs 2raluca.cortelÎncă nu există evaluări
- Informații Suplimentare Despre Organitele BimembranareDocument15 paginiInformații Suplimentare Despre Organitele BimembranareIrina SpoialoÎncă nu există evaluări
- Celula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Document1 paginăCelula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Gabor IzabellaÎncă nu există evaluări
- Document RTF NouDocument43 paginiDocument RTF NounicuracoviteanuÎncă nu există evaluări
- BiologiebababDocument7 paginiBiologiebababCraciunica DragosÎncă nu există evaluări
- BiologieDocument2 paginiBiologieamarandiÎncă nu există evaluări
- Anatomia Si Fiziologia Omului Compendiu - FragmentDocument13 paginiAnatomia Si Fiziologia Omului Compendiu - FragmentBaltaroiu MariusÎncă nu există evaluări
- Tema 2 FSASM Celula PDFDocument8 paginiTema 2 FSASM Celula PDFSergiu GucaÎncă nu există evaluări
- Tema 2 CelulaDocument13 paginiTema 2 CelulaCristina CurasovÎncă nu există evaluări
- Zoologie Notite 2018Document27 paginiZoologie Notite 2018Alle BedereagÎncă nu există evaluări
- Celula-Eucariota-2 - ModificataDocument10 paginiCelula-Eucariota-2 - ModificataRu MiuÎncă nu există evaluări
- Celula Animala Si Celula VegetalaDocument4 paginiCelula Animala Si Celula VegetalaBenn BennetÎncă nu există evaluări
- BioDocument7 paginiBioMihaela StoenescuÎncă nu există evaluări
- 5 Biofizica CelularăDocument70 pagini5 Biofizica CelularăBalint Ana MariaÎncă nu există evaluări