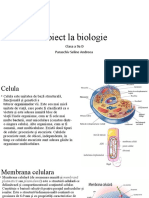
S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- NucleuDocument17 paginiNucleuTudor OstopoviciÎncă nu există evaluări
- LP6 NucleuDocument15 paginiLP6 NucleuMihail RoscaÎncă nu există evaluări
- Tema 2. Nucleul PDFDocument6 paginiTema 2. Nucleul PDFAlina PinzaruÎncă nu există evaluări
- Nucleul CelularDocument7 paginiNucleul Celularcandi7Încă nu există evaluări
- Nucleul Si Reproducerea Celulara Curs 2012 2013Document12 paginiNucleul Si Reproducerea Celulara Curs 2012 2013Bejenaru GeorgeÎncă nu există evaluări
- Nucleolul Şi Funcţiile LuiDocument2 paginiNucleolul Şi Funcţiile LuiNutza Zadic100% (1)
- BioCel Curs 2 Nucleul Partea IDocument10 paginiBioCel Curs 2 Nucleul Partea IMarin TheodorÎncă nu există evaluări
- NUCLEULDocument71 paginiNUCLEULAndrada Raicu100% (1)
- Biologie Celulara LPDocument5 paginiBiologie Celulara LPRebeca FlorinaÎncă nu există evaluări
- Biologie Celulara Si Moleculara: Capitolul 9 Nucleul - Organizarea Și Rolul NucleuluiDocument31 paginiBiologie Celulara Si Moleculara: Capitolul 9 Nucleul - Organizarea Și Rolul NucleuluiMiruna-CristianaBirtuÎncă nu există evaluări
- CELULE Si CROMOZOMI Referat AnatomieDocument20 paginiCELULE Si CROMOZOMI Referat AnatomieLapadat MihaelaÎncă nu există evaluări
- Curs 4 BiocelDocument26 paginiCurs 4 BiocelBianca TomeciÎncă nu există evaluări
- Nucleul CelularDocument25 paginiNucleul CelularGrațiela CazanÎncă nu există evaluări
- Nucleul CeluleiDocument34 paginiNucleul CeluleiElena PruniciÎncă nu există evaluări
- NUCLEULDocument3 paginiNUCLEULMihai CodinÎncă nu există evaluări
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- Curs 9 Nucleul Si CromatinaDocument23 paginiCurs 9 Nucleul Si CromatinaTurturea ClaudiuÎncă nu există evaluări
- NUCLEULDocument3 paginiNUCLEULcristi123321Încă nu există evaluări
- 1.noţiuni Generale Despre CeluleDocument5 pagini1.noţiuni Generale Despre CeluleAndreea100% (1)
- NucleulDocument3 paginiNucleuldaytayÎncă nu există evaluări
- Document 5Document3 paginiDocument 5Good GuyÎncă nu există evaluări
- NUCLEULDocument9 paginiNUCLEULley_lyÎncă nu există evaluări
- Nucleul - Cap. 6Document5 paginiNucleul - Cap. 6Nicolae IaşanÎncă nu există evaluări
- Replicarea ADNDocument11 paginiReplicarea ADNIulia100% (1)
- Nucleul CelularDocument49 paginiNucleul CelularRaduBercaÎncă nu există evaluări
- Neuron UlDocument6 paginiNeuron UlMariana BudnicovÎncă nu există evaluări
- Curs - Nucleul CelularDocument49 paginiCurs - Nucleul CelularOvidiu DumitruÎncă nu există evaluări
- Celula EucariotaDocument15 paginiCelula Eucariotaiancu alexandru100% (2)
- Neuron UlDocument4 paginiNeuron Uldoctor_roxiÎncă nu există evaluări
- Nucleul - ,,creierul CeluleiDocument33 paginiNucleul - ,,creierul CeluleiDana Semen100% (3)
- Curs Genetică 2 UMFCD 2017-2018Document6 paginiCurs Genetică 2 UMFCD 2017-2018StefanChistolÎncă nu există evaluări
- Proiect BioDocument12 paginiProiect BioLaura GrindeiÎncă nu există evaluări
- 7 - NucleulDocument4 pagini7 - NucleulCiobanu MihaiÎncă nu există evaluări
- BIOLOGIE CELULARA ReferatDocument19 paginiBIOLOGIE CELULARA Referatpraporgicu mirelaÎncă nu există evaluări
- Curs 1 Bio MolecularaDocument123 paginiCurs 1 Bio MolecularaAmalia MoldovanuÎncă nu există evaluări
- CitogeneticaDocument0 paginiCitogeneticaGeorge BrinzeaÎncă nu există evaluări
- Proiect La Biologie: Clasaa9aD Paraschiv Seline AndreeaDocument7 paginiProiect La Biologie: Clasaa9aD Paraschiv Seline Andreea٭BUNNYÎncă nu există evaluări
- Mio LogieDocument80 paginiMio LogieAmal AzkÎncă nu există evaluări
- Celula - Sinteza BACDocument9 paginiCelula - Sinteza BACAndreea Dinita100% (1)
- Nucleul Clasa A Ix ADocument3 paginiNucleul Clasa A Ix AtrafficiasibikeÎncă nu există evaluări
- Genetica MolecularaDocument3 paginiGenetica MolecularaIonuț BuzatuÎncă nu există evaluări
- NeuronulDocument6 paginiNeuronulFilip Filipelu100% (1)
- Curs Celula 2Document50 paginiCurs Celula 2Madalina Elena Răciulă cas RusuÎncă nu există evaluări
- Nucleul - Notite de CursDocument19 paginiNucleul - Notite de CursMorgan Rosewood100% (1)
- Referat Anatomie-Nucleul CelularDocument7 paginiReferat Anatomie-Nucleul CelularAdina GheorgheÎncă nu există evaluări
- 73 e 5 Dfa 1Document4 pagini73 e 5 Dfa 1rovanaÎncă nu există evaluări
- NUCLEULDocument43 paginiNUCLEULBondreaTiberiuÎncă nu există evaluări
- WWW - Referateok.ro: NeuronulDocument7 paginiWWW - Referateok.ro: NeuronulBuzatu MadalinaÎncă nu există evaluări
- Organizarea Materialului GeneticDocument4 paginiOrganizarea Materialului GeneticmireaÎncă nu există evaluări
- proiect bio clasa a 9-aDocument4 paginiproiect bio clasa a 9-abadeaionutdavid19Încă nu există evaluări
- GeneticaDocument31 paginiGeneticaadygorincioiÎncă nu există evaluări