S-ar putea să vă placă și
- NucleuDocument17 paginiNucleuTudor OstopoviciÎncă nu există evaluări
- NUCLEUL1Document15 paginiNUCLEUL1PetrnelaÎncă nu există evaluări
- Tema 2. Nucleul PDFDocument6 paginiTema 2. Nucleul PDFAlina PinzaruÎncă nu există evaluări
- Nucleul CelularDocument25 paginiNucleul CelularGrațiela CazanÎncă nu există evaluări
- Nucleul CelularDocument7 paginiNucleul Celularcandi7Încă nu există evaluări
- NUCLEULDocument71 paginiNUCLEULAndrada Raicu100% (1)
- Nucleul CeluleiDocument34 paginiNucleul CeluleiElena PruniciÎncă nu există evaluări
- Document 5Document3 paginiDocument 5Good GuyÎncă nu există evaluări
- Nucleolul Şi Funcţiile LuiDocument2 paginiNucleolul Şi Funcţiile LuiNutza Zadic100% (1)
- Biologie Celulara Si Moleculara: Capitolul 9 Nucleul - Organizarea Și Rolul NucleuluiDocument31 paginiBiologie Celulara Si Moleculara: Capitolul 9 Nucleul - Organizarea Și Rolul NucleuluiMiruna-CristianaBirtuÎncă nu există evaluări
- Curs 9 Nucleul Si CromatinaDocument23 paginiCurs 9 Nucleul Si CromatinaTurturea ClaudiuÎncă nu există evaluări
- Curs 4 BiocelDocument26 paginiCurs 4 BiocelBianca TomeciÎncă nu există evaluări
- BioCel Curs 2 Nucleul Partea IDocument10 paginiBioCel Curs 2 Nucleul Partea IMarin TheodorÎncă nu există evaluări
- CELULE Si CROMOZOMI Referat AnatomieDocument20 paginiCELULE Si CROMOZOMI Referat AnatomieLapadat MihaelaÎncă nu există evaluări
- Replicarea ADNDocument11 paginiReplicarea ADNIulia100% (1)
- Nucleul - Cap. 6Document5 paginiNucleul - Cap. 6Nicolae IaşanÎncă nu există evaluări
- Celula EucariotaDocument15 paginiCelula Eucariotaiancu alexandru100% (2)
- Biologie Celulara LPDocument5 paginiBiologie Celulara LPRebeca FlorinaÎncă nu există evaluări
- Curs Celula 2Document50 paginiCurs Celula 2Madalina Elena Răciulă cas RusuÎncă nu există evaluări
- Nucleul Si Reproducerea Celulara Curs 2012 2013Document12 paginiNucleul Si Reproducerea Celulara Curs 2012 2013Bejenaru GeorgeÎncă nu există evaluări
- 1.noţiuni Generale Despre CeluleDocument5 pagini1.noţiuni Generale Despre CeluleAndreea100% (1)
- Curs Genetică 2 UMFCD 2017-2018Document6 paginiCurs Genetică 2 UMFCD 2017-2018StefanChistolÎncă nu există evaluări
- Curs 1 Bio MolecularaDocument123 paginiCurs 1 Bio MolecularaAmalia MoldovanuÎncă nu există evaluări
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- BIOLOGIE CELULARA ReferatDocument19 paginiBIOLOGIE CELULARA Referatpraporgicu mirelaÎncă nu există evaluări
- 7 - NucleulDocument4 pagini7 - NucleulCiobanu MihaiÎncă nu există evaluări
- NucleulDocument3 paginiNucleuldaytayÎncă nu există evaluări
- NUCLEULDocument43 paginiNUCLEULBondreaTiberiuÎncă nu există evaluări
- AMG Teme Examen Teme 8,9,10,11Document18 paginiAMG Teme Examen Teme 8,9,10,11Iulia GeorgianaÎncă nu există evaluări
- Acizii Nucleici Structura Si FunctiiDocument34 paginiAcizii Nucleici Structura Si FunctiiFEVRONIA PAHONTUÎncă nu există evaluări
- Tema 2. NucleulDocument5 paginiTema 2. NucleulAnicika BalicaÎncă nu există evaluări
- Nucleul - Notite de CursDocument19 paginiNucleul - Notite de CursMorgan Rosewood100% (1)
- CitogeneticaDocument0 paginiCitogeneticaGeorge BrinzeaÎncă nu există evaluări
- CelulaDocument15 paginiCelulasebiyÎncă nu există evaluări
- Neuroni Si Celule GlialeDocument8 paginiNeuroni Si Celule GlialeElenaÎncă nu există evaluări
- Referat Anatomie-Nucleul CelularDocument7 paginiReferat Anatomie-Nucleul CelularAdina GheorgheÎncă nu există evaluări
- Cap I - 2 - Elemente de Genetica MolecularaDocument45 paginiCap I - 2 - Elemente de Genetica MolecularaBogdan ButilcaÎncă nu există evaluări
- Organizarea Materialului Genetic La EucarioteDocument17 paginiOrganizarea Materialului Genetic La EucarioteAndra GabrielaÎncă nu există evaluări
- Organite Celulare ComuneDocument13 paginiOrganite Celulare ComuneDennisÎncă nu există evaluări
- NUCLEULDocument3 paginiNUCLEULMihai CodinÎncă nu există evaluări
- Organizarea Materialului GeneticDocument4 paginiOrganizarea Materialului GeneticmireaÎncă nu există evaluări
- Nucleul Clasa A Ix ADocument3 paginiNucleul Clasa A Ix AtrafficiasibikeÎncă nu există evaluări
- Genetica MolecularaDocument3 paginiGenetica MolecularaIonuț BuzatuÎncă nu există evaluări
- Aparatul Genetic Al Celulelor Umane Este Format Din Structuri Celulare Ce Conţin ADNDocument1 paginăAparatul Genetic Al Celulelor Umane Este Format Din Structuri Celulare Ce Conţin ADNDaniela DedovÎncă nu există evaluări
- Biologie CelularaDocument193 paginiBiologie CelularaAndrei GOÎncă nu există evaluări
- NUCLEULDocument9 paginiNUCLEULley_lyÎncă nu există evaluări
- Nucleul - ,,creierul CeluleiDocument33 paginiNucleul - ,,creierul CeluleiDana Semen100% (3)
- Mio LogieDocument80 paginiMio LogieAmal AzkÎncă nu există evaluări
- 06 Nucleul InterfazicDocument48 pagini06 Nucleul InterfazicANCA OANCEAÎncă nu există evaluări
- Nucleul CursDocument11 paginiNucleul CursFekete VictoriaÎncă nu există evaluări
- Referat Biologie-Oprea Beatrice Iulia, Clasa 9BDocument11 paginiReferat Biologie-Oprea Beatrice Iulia, Clasa 9BBeatrice Iulia Oprea0% (1)
- Abdomen AcutDocument3 paginiAbdomen AcutMihail RoscaÎncă nu există evaluări
- Capitol 6 Nucleu ROMDocument10 paginiCapitol 6 Nucleu ROMMihail RoscaÎncă nu există evaluări
- Formularul Fisei de PostDocument3 paginiFormularul Fisei de PostMihail RoscaÎncă nu există evaluări
- Test de Evaluare A CunostintelorDocument2 paginiTest de Evaluare A CunostintelorMihail RoscaÎncă nu există evaluări
- TumorileDocument4 paginiTumorileMihail RoscaÎncă nu există evaluări
- Biofizica ExamenDocument27 paginiBiofizica ExamenIgor CasapciucÎncă nu există evaluări
- Capitol 4 Organite Celulare ROMDocument15 paginiCapitol 4 Organite Celulare ROMVictoria IachimÎncă nu există evaluări
- Capitol 2 Macromolecule ROMDocument10 paginiCapitol 2 Macromolecule ROMAndrei RusuÎncă nu există evaluări
- Muschii Origine Si InsertieDocument13 paginiMuschii Origine Si InsertieAna Maria IvanceaÎncă nu există evaluări
- Ciclul CelularDocument15 paginiCiclul CelularDiana TriliÎncă nu există evaluări
- Capitol 5 Procariote ROMDocument11 paginiCapitol 5 Procariote ROMAndrei RusuÎncă nu există evaluări
- Modul Sanatos de ViataDocument11 paginiModul Sanatos de ViataMihail RoscaÎncă nu există evaluări
- Replicarea Si Repararea Adn 2015Document21 paginiReplicarea Si Repararea Adn 2015Ion Cotoneț100% (2)
- Abdomen AcutDocument3 paginiAbdomen AcutMihail RoscaÎncă nu există evaluări
- FracturileDocument3 paginiFracturileMihail RoscaÎncă nu există evaluări
- Agresivitatea UmanaDocument25 paginiAgresivitatea UmanaMihail RoscaÎncă nu există evaluări
- Combustia TermicaDocument4 paginiCombustia TermicaMihail RoscaÎncă nu există evaluări
- BotulismDocument5 paginiBotulismMihail RoscaÎncă nu există evaluări
- Inervatia PieliiDocument2 paginiInervatia PieliiMihail Rosca50% (2)
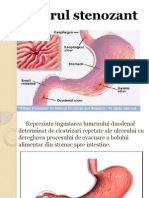
- Ulcerul StenozantDocument15 paginiUlcerul StenozantMihail RoscaÎncă nu există evaluări
- Ulcerul StenozantDocument15 paginiUlcerul StenozantMihail RoscaÎncă nu există evaluări
- Ascultarea ActivaDocument17 paginiAscultarea ActivaMihail RoscaÎncă nu există evaluări
- Nevoia de A EliminaDocument27 paginiNevoia de A EliminaMihail RoscaÎncă nu există evaluări