S-ar putea să vă placă și
- 1 SNC - SomesteziaDocument9 pagini1 SNC - SomesteziaAttila SzakacsÎncă nu există evaluări
- Analizatorii - Doc FBDocument27 paginiAnalizatorii - Doc FBCamelia ManeaÎncă nu există evaluări
- Analizatori ReferatDocument10 paginiAnalizatori ReferatPetrica Anca100% (1)
- Anatomie Curs 3Document6 paginiAnatomie Curs 3Ionela Marinela BadescuÎncă nu există evaluări
- 7 Tratamentul Bolilor GeneticeDocument5 pagini7 Tratamentul Bolilor GeneticeAlexandru PaulÎncă nu există evaluări
- 5 Suport de Curs - Elemente de Botanica - Fiziologie VegetalaDocument24 pagini5 Suport de Curs - Elemente de Botanica - Fiziologie VegetalaDana MoraruÎncă nu există evaluări
- AnalizatoriiDocument2 paginiAnalizatoriiAna Maria ToporascuÎncă nu există evaluări
- Curs 7. Cerebel, Diencefal, Scoarta CerebralaDocument20 paginiCurs 7. Cerebel, Diencefal, Scoarta Cerebralaxaxek59161Încă nu există evaluări
- Recapitulare AnalizatoriDocument6 paginiRecapitulare AnalizatoriiuliamaghiarÎncă nu există evaluări
- Anatomie Curs 13,14-Amg - AnalizatoriiDocument10 paginiAnatomie Curs 13,14-Amg - AnalizatoriiAlexandru GiubegaÎncă nu există evaluări
- Analizatorul KinestezicDocument4 paginiAnalizatorul KinestezicAndreea Mihaela LazărÎncă nu există evaluări
- Sistemul ExcretorDocument37 paginiSistemul ExcretorAndrei CrÎncă nu există evaluări
- AnalizatoriDocument32 paginiAnalizatoriValeriu VazdautanÎncă nu există evaluări
- Nucleii Branhiomotori Din Trunchiul CerebralDocument2 paginiNucleii Branhiomotori Din Trunchiul CerebralMihai Alexandru ButnariuÎncă nu există evaluări
- FarmaDocument41 paginiFarmaOctaviaCosminaÎncă nu există evaluări
- Anatomie I FinalDocument81 paginiAnatomie I Finalאנדריאה סיובאנוÎncă nu există evaluări
- Curs 1 - Org Sist ImunDocument44 paginiCurs 1 - Org Sist ImunDaniel Ciprian50% (2)
- Fiziologia Tesuturilor ExcitabileDocument72 paginiFiziologia Tesuturilor ExcitabileNellu CebotariÎncă nu există evaluări
- Analizatorul Gustativ Si Vestibular - Curs 12Document5 paginiAnalizatorul Gustativ Si Vestibular - Curs 12Ioana Navalici100% (1)
- Histologie 1 Curs 12) Tesutul NervosDocument7 paginiHistologie 1 Curs 12) Tesutul NervosMariaMaryaÎncă nu există evaluări
- Fiziologia Sistemului Nervos VegetativDocument14 paginiFiziologia Sistemului Nervos VegetativAlina Paşca100% (1)
- Curs 2 Mcurs 3 NeuroanatomieDocument36 paginiCurs 2 Mcurs 3 NeuroanatomieWilliambutacuÎncă nu există evaluări
- Strategii de Terapie A Bolilor GeneticeDocument20 paginiStrategii de Terapie A Bolilor GeneticeALIN-MARIAN VASILEÎncă nu există evaluări
- Curs 1 MicroscopieDocument30 paginiCurs 1 Microscopieiustin rotariuÎncă nu există evaluări
- Curs 5 Tesutul Osos 2012Document103 paginiCurs 5 Tesutul Osos 2012Raluca TorsinÎncă nu există evaluări
- Capitolul I CELULA SI TESUTURIDocument14 paginiCapitolul I CELULA SI TESUTURIVioleta DiaconuÎncă nu există evaluări
- Curs 3 Trunchiul CerebralDocument68 paginiCurs 3 Trunchiul CerebralPetrica Taranu100% (2)
- Curs 4 - AmgDocument7 paginiCurs 4 - AmgAndreea VladÎncă nu există evaluări
- Nervii CranieniDocument18 paginiNervii CranieniCristina CurcubătÎncă nu există evaluări
- Biologie XDocument5 paginiBiologie XMaya SingerÎncă nu există evaluări
- Sistemul Nervos.Document5 paginiSistemul Nervos.AlexandruSîrbuÎncă nu există evaluări
- Referat docxVIOLETADocument10 paginiReferat docxVIOLETAtuturici liviuÎncă nu există evaluări
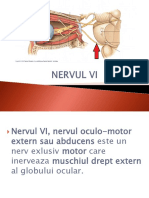
- Nervul ViDocument14 paginiNervul ViOana OanaÎncă nu există evaluări
- Acizii Nucleici 2Document146 paginiAcizii Nucleici 2nircavadim1Încă nu există evaluări
- Analizatorii - GeneralitatiDocument5 paginiAnalizatorii - GeneralitatiDrăgoi Ionuț100% (1)
- Proprietatile Macro Si Microorganismelor Participante La Procesul InfectiosDocument3 paginiProprietatile Macro Si Microorganismelor Participante La Procesul InfectiosAngheluta GabrielaÎncă nu există evaluări
- Curs 2 Curs 2 Sistemul NervosDocument6 paginiCurs 2 Curs 2 Sistemul NervosIulian Catalin GheorghiuÎncă nu există evaluări
- Sistemul Nervos Vegetativ - COLINERGICDocument14 paginiSistemul Nervos Vegetativ - COLINERGICMihai MarinÎncă nu există evaluări
- Factori de CrestereDocument6 paginiFactori de Cresterelouis_miuÎncă nu există evaluări
- Olimpiada11 XI Subiecte Cluj Judet 2022Document17 paginiOlimpiada11 XI Subiecte Cluj Judet 2022Mara DickÎncă nu există evaluări
- 2014 Biologie Locala Arad Clasa A Xia SubiectebaremDocument13 pagini2014 Biologie Locala Arad Clasa A Xia SubiectebaremJane458Încă nu există evaluări
- Curs AMG 13 - Sistem Endocrin HemolimfaticDocument24 paginiCurs AMG 13 - Sistem Endocrin HemolimfaticAndreea Gheorghiță100% (1)
- 2 NeuronulDocument7 pagini2 NeuronulAncaMedÎncă nu există evaluări
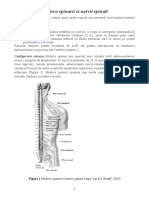
- Maduva Spinarii Si Nervii SpinaliDocument9 paginiMaduva Spinarii Si Nervii SpinalinetprintÎncă nu există evaluări
- AMG - Caractere Generale Ale VirusurilorDocument29 paginiAMG - Caractere Generale Ale VirusurilorSorina UrsacheÎncă nu există evaluări
- Coaste, Stern, VertebreDocument21 paginiCoaste, Stern, VertebreAlina LambaÎncă nu există evaluări
- Curs 2 - Muschi, FacialDocument8 paginiCurs 2 - Muschi, FacialCristina IoanaÎncă nu există evaluări
- CEREBELULDocument8 paginiCEREBELULVlad PerpeleaÎncă nu există evaluări
- Genomica - Mutatiile DinamiceDocument11 paginiGenomica - Mutatiile DinamiceBogdan Tecioiu100% (1)
- Nervul Trigemen AnatomieDocument14 paginiNervul Trigemen AnatomieDunas DanÎncă nu există evaluări
- Analizatorii Si Glandele EndocrineDocument12 paginiAnalizatorii Si Glandele EndocrineMonica Amalia Fasie Halbac100% (1)
- Curs 3 - Trunchiul CerebralDocument68 paginiCurs 3 - Trunchiul CerebralAlina Anca100% (1)
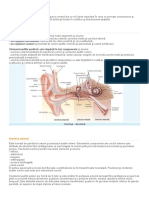
- UrecheaDocument4 paginiUrecheaMarian MaryanÎncă nu există evaluări
- BactDocument63 paginiBactLaura Tarlapan RomanÎncă nu există evaluări
- ImunoDocument152 paginiImunoTeodora MunteanuÎncă nu există evaluări
- VIRUSOLOGIE Suport Curs 2018 - 2019 PDFDocument24 paginiVIRUSOLOGIE Suport Curs 2018 - 2019 PDFEcaterina CapusneacÎncă nu există evaluări
- Analizatorul Cutanat FiziologieDocument19 paginiAnalizatorul Cutanat FiziologieLarisa Roșca100% (3)
- AnalizatoriiDocument49 paginiAnalizatoriiMihaela LupanÎncă nu există evaluări
- AnalizatoriiDocument3 paginiAnalizatoriiDanÎncă nu există evaluări
- AnalizatoriiDocument36 paginiAnalizatoriideaconuandrei635Încă nu există evaluări
- Program Congres AMRDocument16 paginiProgram Congres AMRCostin CaruntuÎncă nu există evaluări
- Program Congres AMRDocument16 paginiProgram Congres AMRCostin CaruntuÎncă nu există evaluări
- Analizatorii Gustativ Si Olfactiv 2008Document8 paginiAnalizatorii Gustativ Si Olfactiv 2008Costin CaruntuÎncă nu există evaluări
- Analizatorii Gustativ Şi OlfactivDocument8 paginiAnalizatorii Gustativ Şi OlfactivCostin CaruntuÎncă nu există evaluări
- Curs Analizatorul Cutanat 2008Document14 paginiCurs Analizatorul Cutanat 2008Costin CaruntuÎncă nu există evaluări
- Analizatorii Gustativ Şi OlfactivDocument8 paginiAnalizatorii Gustativ Şi OlfactivCostin CaruntuÎncă nu există evaluări
- Curs Analizatori ChimiciDocument41 paginiCurs Analizatori ChimiciCostin CaruntuÎncă nu există evaluări
- Ghid Dermato-VenerologieDocument74 paginiGhid Dermato-VenerologieCristian100% (3)