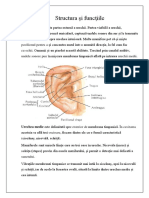
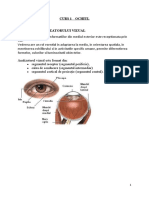
S-ar putea să vă placă și
- Referat OchiulDocument5 paginiReferat OchiulAna-Maria TrandafirescuÎncă nu există evaluări
- Formarea Imaginii Pe RetinaDocument7 paginiFormarea Imaginii Pe RetinaRosalinda MironÎncă nu există evaluări
- Referat Ochiul Si VedereaDocument6 paginiReferat Ochiul Si VedereaRăcăreanu Ştefan100% (1)
- Curs 2-Diviziunea Celulara-Mitoza Si MeiozaDocument11 paginiCurs 2-Diviziunea Celulara-Mitoza Si MeiozaNarco BlancoÎncă nu există evaluări
- Fiziologia Analizatorului VizualDocument3 paginiFiziologia Analizatorului VizualAndrey Laiu100% (6)
- REFERAT Analizator VizualDocument5 paginiREFERAT Analizator VizualFlorentina JianuÎncă nu există evaluări
- Calea Vestibulara RO CURS 6Document4 paginiCalea Vestibulara RO CURS 6cindyCSÎncă nu există evaluări
- OchiulDocument5 paginiOchiulpasztor mariaÎncă nu există evaluări
- Analizatorul VizualDocument16 paginiAnalizatorul VizualTeodora Nanu100% (2)
- Structura Globului OcularDocument6 paginiStructura Globului OcularLidia VlaseÎncă nu există evaluări
- Referat AnatomieDocument12 paginiReferat AnatomieFlorentina Jianu100% (1)
- Boli Ale Analizatorului Vizual (Clasele XI - XII)Document1 paginăBoli Ale Analizatorului Vizual (Clasele XI - XII)Florin Caroli100% (1)
- Reflexia Si Refractia LuminiiDocument6 paginiReflexia Si Refractia Luminiihellen06Încă nu există evaluări
- LentileDocument3 paginiLentileMircea MiculasÎncă nu există evaluări
- Difracţia LuminiiDocument11 paginiDifracţia LuminiiMilky Cookie100% (2)
- Ţesutul OsosDocument4 paginiŢesutul OsosDaniela MariaÎncă nu există evaluări
- Analizatorul VizualDocument9 paginiAnalizatorul VizualRaluca RarincaÎncă nu există evaluări
- Ochiul Uman in FizicaDocument15 paginiOchiul Uman in FizicaFlorea Alin100% (2)
- Sistemul Digestiv La MamifereDocument21 paginiSistemul Digestiv La MamifereIoana Trif100% (1)
- Fenomene Optice Intalnite in NaturaDocument7 paginiFenomene Optice Intalnite in NaturaDorian Mihai CiubotaruÎncă nu există evaluări
- Fenomene OndulatoriiDocument4 paginiFenomene OndulatoriiAndrada PapazianÎncă nu există evaluări
- Particularitățile Structurale Ale Sistemului Respirator La VertebrateDocument11 paginiParticularitățile Structurale Ale Sistemului Respirator La VertebratedanÎncă nu există evaluări
- ADN - Caracteristici Pe ScurtDocument1 paginăADN - Caracteristici Pe ScurtAdonis IchimÎncă nu există evaluări
- LunetaDocument10 paginiLunetaTodoran Maria CristinaÎncă nu există evaluări
- Pancreasul EndocrinDocument2 paginiPancreasul EndocrinElena Vizireanu100% (1)
- Celula ProcariotaDocument10 paginiCelula ProcariotaAndreea VirtejÎncă nu există evaluări
- Mecanism VizualDocument33 paginiMecanism VizualDiana DodoneteÎncă nu există evaluări
- Spectrul de AbsorbțieDocument13 paginiSpectrul de AbsorbțieVlada CovalciucÎncă nu există evaluări
- Fotosinteza Cls A 10-ADocument16 paginiFotosinteza Cls A 10-ACremona Murariu100% (2)
- UrecheaDocument14 paginiUrecheaVlad StatiÎncă nu există evaluări
- CelulaDocument3 paginiCelulaSanislai DanielÎncă nu există evaluări
- Actiunea Radiatiilor Luminoase Si Ultravioletele Asupra OrganismelorDocument8 paginiActiunea Radiatiilor Luminoase Si Ultravioletele Asupra OrganismelorRadulescuCarmen-IoanaÎncă nu există evaluări
- Aparatul DioptricDocument2 paginiAparatul DioptricLidia VlaseÎncă nu există evaluări
- Ochiul Uman - Instrument OpticDocument7 paginiOchiul Uman - Instrument Opticsorina512100% (1)
- Sindromul OndineiDocument8 paginiSindromul OndineiDoina VinagaÎncă nu există evaluări
- 05 Curs Receptie VizualaDocument17 pagini05 Curs Receptie VizualaLoveMed09Încă nu există evaluări
- Organe Anexe Ale Globului Ocular - WWW - IreferateDocument5 paginiOrgane Anexe Ale Globului Ocular - WWW - IreferateStrat Ștefan Gelu100% (1)
- Digestia Si AbsorbtiaDocument32 paginiDigestia Si AbsorbtiaCarmen AmuzaÎncă nu există evaluări
- HIPERMETROPIA ReferatDocument4 paginiHIPERMETROPIA ReferatZubjeschi AlexandruÎncă nu există evaluări
- Celula - Sinteza BACDocument9 paginiCelula - Sinteza BACAndreea Dinita100% (1)
- Defectele OchiuluiDocument2 paginiDefectele OchiuluiAdina BratuÎncă nu există evaluări
- Membrana Celulara PDFDocument40 paginiMembrana Celulara PDFOzana CopyShop100% (1)
- OchiulDocument10 paginiOchiulDorina Velicea100% (1)
- LICHENIIDocument11 paginiLICHENIIPetru GiraÎncă nu există evaluări
- SEGMENTELE AnalizatorilorDocument4 paginiSEGMENTELE AnalizatorilorŞtefania TanislăvescuÎncă nu există evaluări
- Analizatorii (Biologie)Document11 paginiAnalizatorii (Biologie)Crisan SimonaÎncă nu există evaluări
- Tesutul NervosDocument32 paginiTesutul NervosInga PlatonÎncă nu există evaluări
- Instrumente OpticeDocument6 paginiInstrumente OpticeAnonymous obY2jN7OZÎncă nu există evaluări
- Dioda ElectroluminiscentaDocument9 paginiDioda ElectroluminiscentaDamian StefanÎncă nu există evaluări
- Schema Pe TablaDocument4 paginiSchema Pe Tabla3x3cu7or100% (1)
- Referat La Biologie "Analizatorul Vizual"Document18 paginiReferat La Biologie "Analizatorul Vizual"Victoria Chiperi Andreicu100% (1)
- Anatomia Si Fiziologia Omului. Curs 7 - Analizatorul Vizual, Acustico-Vestibular, Motor (Kinestezic)Document9 paginiAnatomia Si Fiziologia Omului. Curs 7 - Analizatorul Vizual, Acustico-Vestibular, Motor (Kinestezic)Palea BiancaÎncă nu există evaluări
- Analizatorul VizualDocument4 paginiAnalizatorul VizualRadu Andrei CătinasÎncă nu există evaluări
- Analizatorul VizualDocument3 paginiAnalizatorul VizualMihaela CanțărÎncă nu există evaluări
- Fiziologia Analizatorului VizualDocument18 paginiFiziologia Analizatorului VizualDieFerÎncă nu există evaluări
- Curs 1 OchiulDocument14 paginiCurs 1 OchiulEvelina ȚaralungaÎncă nu există evaluări
- Analizator Vizual PPTDocument9 paginiAnalizator Vizual PPTbilchirufricanhÎncă nu există evaluări
- Licenta-P1 2Document59 paginiLicenta-P1 2Rudi Mihai100% (1)
- Ochiul, Fiziologia VederiiDocument5 paginiOchiul, Fiziologia VederiiDiana NiculescuÎncă nu există evaluări
- Mezotimpanită Cronică În AcutizareDocument6 paginiMezotimpanită Cronică În AcutizareAlina BraduÎncă nu există evaluări
- Deprinderi Practice DetaliatDocument4 paginiDeprinderi Practice DetaliatAla Grițcan-BîtcaÎncă nu există evaluări
- Orar Lectii OptionaleDocument1 paginăOrar Lectii OptionaleAna ScobioalaÎncă nu există evaluări
- Ciupi md-v12706105Document1 paginăCiupi md-v12706105Ana ScobioalaÎncă nu există evaluări
- Anemii Hemolitice Copii PDFDocument5 paginiAnemii Hemolitice Copii PDFAlina DercaciÎncă nu există evaluări
- Lista Intrebari Urologie 2010 UmfiasiDocument7 paginiLista Intrebari Urologie 2010 Umfiasit1bbiÎncă nu există evaluări
- Anul IV Sem II Med 1Document2 paginiAnul IV Sem II Med 1Irina OdajiuÎncă nu există evaluări
- LuxatiiDocument28 paginiLuxatiiIacob LocomanÎncă nu există evaluări
- Cartilajul ArticularDocument9 paginiCartilajul ArticularDanielle NievesÎncă nu există evaluări
- Fisa UrologieDocument8 paginiFisa UrologieAna ScobioalaÎncă nu există evaluări
- Medicina CalamitatilorDocument12 paginiMedicina CalamitatilorNicolae Zaharia100% (3)
- Fiziopatologia Sistemului CardiovascularDocument57 paginiFiziopatologia Sistemului CardiovascularAlina VerdesÎncă nu există evaluări
- Urologie Nefrologi Chirurgicala 2005Document111 paginiUrologie Nefrologi Chirurgicala 2005alexdaria100% (1)
- Anul IV 2Document2 paginiAnul IV 2Irina OdajiuÎncă nu există evaluări