Documente Academic
Documente Profesional
Documente Cultură
ANATOMIEE
Încărcat de
Ioana MuntianuDrepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
ANATOMIEE
Încărcat de
Ioana MuntianuDrepturi de autor:
Formate disponibile
NIVELURI DE ORGANIZARE ANATOMICA SI FIZIOLOGICA Anatomia este disciplina care se ocupa cu descrierea corpului uman.
Corpul uman este un intreg functional, armonios, compus din mai multe aparate si sisteme. Aparatul este ansamblul de organe cu structura si origine diferita care serveste la indeplinirea unei functii.De ex: aparatul circulator este compus din inima, artere, capilare, vene, vase si noduli limfatici; aparatul digestiv este compus din cavitatea bucala, esofagul, stamacul, intestinul subtire, intestinul gros, anus, ficat, pancreas. Sistemul este ansamblul de elemente materiale sau ideale care functioneaza ca un intreg datorita interdependentei dintre ele. Notiunea de sistem trebuie privita la diferite nivele: fizic, biologic, energetic etc... Organismul uman este ca orice organism viu un sistem deschis, care ace schimb permanent cu substante si energie din mediul incinjurator. Omul poate fi privit din punct de vedere anatomic, histologic, fiziologic ca fiind alcatuit din ansamble diferite de sisteme. Sistemul circulator mentine constant, cantitativ si calitativ lichidul tisular, deci mediul intern, in toate regiunile organismului. Pe de alta parte, circulatia, deplasarea continua a sangelui de-a lungul circuitului inima-periferie-inima reprezinta una din principalele legaturi materiale dintre mediul extern (ambiant0 si organismul ca sistem deschis energetic si metabolic. Sangele circula in interiorul arborelui vascular datorita diferentelor de presiune dintre diferitele segmente ale acestuia, deplasarea fiind conditionata si determinata de numerosi factori printre care cel mai important este activitatea ritmica a inimii. Acest sistem integrator al organismului are rolul de a aduce oxigenul si substantele nutritive la nicelul tesuturilor si de a duce dioxidul de carbon si produsii de dezasimilare (metabolitii) la nivelul organelor unde se realizeaza eliminarea acestora. Sistemul circulator este alcatuit dintr-un organ central numit inima si un sistem de vase prin care circula sangele (artere si vene0 si limfa (vasele limfatice). Structura comuna si esentiala a intregului sistem este endoteliul, reprezentat de un epiteliu simplu pavimentos, care provine din insule sangvine din a caror elemente centrale se diferentiaza hematiile primitive. Sistemul imunitar este extrem de complex, cu roluri importante in mentinerea homeostaziei si sanatatii. Ca si sistemul endocrin, exercita un reglaj al organismului odata cu functiile de aparare, datorita unor componente circulante capabile de a functiona in locuri foarte indepartate de punctul lor de origine. Complexitatea sistemului rezulta dintro retea intricata de comunicari capabile de a exercita efecte multiple, avand la baza un numar relativ redus de tipuri celulare distincte. In felul acesta mecanismul de reglare imunitar poate amplifica enorm un raspuns dat sau il poate diminua considerabil, in functie de necesitatile de moment ale organismului. Un sistem imunitar care functioneaza normal reprezinta o aparare eficienta impotriva unor particule straine cum sunt agentii microbieni patogeni si impotriva unor celule native care au fost supuse unei transformari neoplazice. O functionare defectuoasa a sistemului imun duce la imbolnaviri.
Sistemul neuroendocrin este unul din sistemele de integrare ale ogranismului cu rol esential, supraordonat celorlalte. Legaturile foarte stranse, anatomice vasculare, si nervoase existente intre sistemul nervos si endocrin la nivelul formatiunilor hipotalamo-hipofizare, cat si unele structuri microscopice precum receptorii endocrini din encefal, la care se adauga neurotransmitatorii sintetizati in diferite etaje ale sistemului nervos central (cu rol asupra glandelor endocrine) justifica o integrare a functiilor nervoase si endocrine ce formeaza o unitate sub raport functional.Sistemul neuroendocrin impreuna cu sistemele vascular si imunitar aflate sub controlul sau, realizeaza mentinerea in limite fiziologice a homeostaziei organismului, asigurand adaptarea acestuia la diferitele conditii ale mediului extern. Aparatul de import al materiei este complexul structural format din aparatul respirator ce asigura oxigenul necesar proceselor metabolice si aparatul digestiv pe calea caruia sunt introduse in organism si apoi transportate substante alimentare care contin glucide, proteine, lipide, hidroelectroliti si vitamine. Aparatul de export al materiei cuprinde aparatul renal si pielea, organe cu rol principal in eliminarea produsilor toxici rezultati in organism in urma complexelor procese metabolice eliminatoare de energie. Aparatul de reproducere. Producerea gametilor (ovule si spermatozoizi), unirea lor, dezvoltarea zigotului (celula-ou rezultata din unirea spermatozoidului cu ovulul) necesita o diferentiere specifica a organelor genitale. Aceste organe au in alcatuirea lor: gonadele, ce produc gametii si hormonii sexuali, caile genitale ce servesc transportului gametilor, iar la sexul feminin dezvoltarea oului intrauterin, glandele de pe traiectul aparatului genital, organele genitale externe. Aparatul de sustinere si miscare este format din sistemul osos, sistemul articular si sistemul muscular.
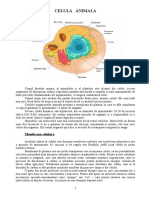
CELULA Celula, unitatea morfofunctionala de baza a organismului, prezinta aspecte specifice de totalitate date de ansamblul partilor ei componente: hialoplasma, nucleu, mitocondrii, lizozomi, ribozomi. Activitatile specifice fiecarui component sunt posibile tocmai in cadrul ansamblului celular, pentru ca celula nu reprezinta numai o sumare cumulativa a structurilor componenete ci si rezultanta relatiilor functionale dintre ele. Hialoplasma reprezinta partea omogena, nestructurata a citoplasmei, zona de mediere a schimburilor si proceselor metabolice specifice organitelor celulare. In hialoplasma se orienteaza microtubuli, forme diferentiate de organizare a unor complexe macromoleculare proteice cu rol de citoschelet (mentine forma celulei si realizeaza supartul prelungirilor ei). In acelasi timp este si un suport al contractiilor fundamentale ale hialoplasmei si un sistem de transport intracelular.
La periferie, hialoplasma se dispune diferentiat in structuri cu multiple activitati enzimatice realizand plasmalema sau membrana celulara. Aceasta are in general organizare neomogena, imprimand asimetrie functionala, cum este marginea in perie la celulele absorbante, sinapsa la celulele nervoase. Functia principala a membranei celulare este de a asigura schimbul permanent si adecvat de substante dintre celula si mediul extracelular prin mecanisme de transport si endocitoza. Endocitoza este mecanismul prin care celula inglobeaza in citoplasma particule din mediul inconjurator. Se desfasoare in mai multi timpi: aparitia unei pungi de invaginare, formarea veziculelor de endocitoza, deschiderea si asocierea veziculelor endocitate in lizozomi. In cadrul acestora enzimele lizozomale devin active digerand continutul. Resturile nedigerate formeaza corpii reziduali care sunt asocitati. Exocitoza este procesul ce se efectueaza intr-o secventa inversa fata de a endocitozei si are rol functional deosebit de important pentru celulele secretorii. In unele celule ca celulele endoteliale capilare,, o buna parte a veziculelor endocitate traverseaza intreaga citoplasma, nu se descarca intracelulr si nu sunt folosite de celula. Ele ajung la polul opus formarii lor, unde sunt exocitate. Nucleul este centrul coordonator si reglator al activitatii celulare, depozitarul si transmitatorul specificitatii biologice. Functii: -depozitar al informatiilor genetice in molecula de ADN sub forma secventelor de nucleotide. Molecula ADN contine secvente de trei nucleotide succesive numite triplete ce pot specifica un acid aminat in lantul polipeptidic, formand un codon. Segmentele de ADN responsabile de sinteza unei proteine se numesc gene.Totalitatea genelor din molecula de ADN se numeste genom. -transmiterea informatiei genetice la sediile de sinteza a proteinelor din citoplasma. Transmiterea mesajului genelor, inmagazinat in ADN se realizeaza prin ARN-ul sintetizat tot in nucleu. Exista trei tipuri de ARN: ARNm (mesager), ARNt(transportor), ARNr (ribozomal). ARN mesager este sintetizat la nivelul diferitelor segmente de ADN realizand transcrierea mesajului genetic. -stabilizarea tiparelor ARNm si transferul lor in citoplasma. Nucleolul elaboreaza o proteina ce incorporeaza mesagerul intro structura rezistenta la degradarea intranucleara si intracitoplasmatica. Atat timp cat proteina este legata de ARNm, acesta din urma nu se poate atasa de ribozomi si deci este blocata posibilitatea de cadificare a sintezei proteinelor.Deblocarea este dependenta de prezenta unor metaboliti. -rol in diviziunea celulara si transmiterea informatiei genetice la descendenti. Materialul ereditar este organizat sub forma cromozomilor, de aspect filiform, vizibili mai clar in fazele diviziunii celulare. Morfologic, apar niste formatiuni alungite din doua filamente dispuse paralel si alipite numite cromatide. In cromatide se gasesc dispuse liniar genele. Celulele tesuturilor umane poseda un numar diploid de cramozomi(2n), spre deosebire de celulele sexuale sau gametii care au numar haploid (n) de cromozomi. Celula somatica are doua seturi de cate 23 (23 perechi) adica 46 cromozomi. 22 perechi sunt autozomi si o pereche cromozomi sexuali. Crz. sexuali la femeie pot fi simbolizati XX, iar la barbat XY. -rol in reglarea reactivitatii celulare. Celulele anucleate au un metabolism scazut. VAloarea raportului nuceo-citoplasmatic este mai mare la celulele tinere, care au si metabolism mai intens.
Reticulul endoplamatic face parte impreuna cu mitocondria, aparatul Golgi din sistemul de endomembrane care separa diferite faze intracelulare. Este un sistem circulator endocelular foarte mobil, cu functii ce deriva din raporturile spatiale dinamice cu celelalte sisteme intracelulare. Dupa structura elementelor componente, in raport cu absenta sau prezenta la suprafata membranei canaliculare a granulelor ribozomale este divizata in reticul endoplasmatic neted sau agranular si reticul endoplasmatic rugos sau granular. Are functii legate de: schimburile citoplasmo-extracelulare si nucleocitoplasmatice, sinteza de substante specifice, transmiterea excitatiei si cuplarea cu contractia (fibra musculara), sinteza mediatorilor sinaptici. Ribozomii reprezinta structura citoplasmatica la nivelul careia aminoacizii activati sunt dispusi in lanturi polipeptidice ale proteinelor sintetizate in conformitate cu informatia genetica adusa din nucleul celular de catre ARNm. Ribozomul nu este specializat numai pentru sinteza unei anumite proteine. Lizozomii prin continutul bogat in hidrolaze acide (proteaze, fosfaze, lipaze) sunt capabili sa degradeze constituenti ai materiei vii. Autofagia sau autoliza focala face parte din reinnoirea normala a unor constituenti celulari ca si din procesele de diferentiere. Este tot un proces de digestie intracelulara, dar obiectul degradarii sunt mici portiuni din celula care au suferit un proces degenerativ. In unele cazuri continutul lizozomal poate fi eliberat direct in citoplasma sau chiar in afara celulei, producan zone de liza circumscrisa sau procese degenerative intinse care se soldeaza obisnuit cu moartea celulei.Autoliza celulara a fost descrisa si in procesele de regresiune fiziologica a organelor. Continutul lizozomal poate fi eliberat si exocitat, in acest caz enzimele pot digera molecule si structuri racelulare, cum se produce in osteoclaste in procesele de remaniere a osului. Mitocondria este centrul respirator si energetic celular. Este o structura ovalara alcatuita dintr-o membrana dubla, cea interna prezentand creste ce proemina in matricea mitocondriala. Numarul de mitocondrii dintr-o celula este proportional cu intensitatea proceselor metabolice si amplitudinea miscarilor. Aparatul Golgi este o diferentiere a sistemului vacuolar si canalicular citoplasmatic, fiind format din sacule, microvezicule, vacuole canaliculare, la nivelul carora au loc maturarea si condensarea materialelor sintetizate de celula si apoi eliminarea lor. Centrozomul reprezinta centrul de organizare al aparatului motor nemuscular din celula (aparatul mitotic, cilii, flaagelii). In cursul diviziunii celulare structura sa se complica prin formarea fusului de diviziune ce face posibila separarea si deplasarea cromozomilor spre cei doi poli celulari. Centrozomul participa si la sinteza proteinelor fibrilare din structura cililor si flagelilor si in acelasi timp asigura motilitatea lor. DIVIZIUNEA CELULARA (MITOZA SI MEIOZA) Cromozomul este corpusculul nucleo-proteic ce se observa cu microscopul in timpul diviziunii celulare in mitoza si meioza. Este principalul purtator al bagajului genetic specific speciei si individului. Numarul, marimea si forma cromozomilor sunt constante pentru o anumita specie si de la o generatie la alta.
Substanta nucleara din care sunt alcatuiti cromozomii se numeste cromatina. De-a lungul cromozomilor sunt insirate genele care controleaza caracterele ereditare ale organismului. In diviziune, cromozomii apar formati din doua unitati functionale longitudinale numite cromatide. Acestea reprezinta starea diploida (2n). Diviziunea celulara mitotica prezerva de la o generatie la alta starea diploida. Diviziunea meiotica reduce la jumatate numarul de cromozomi din celula, in asa fel incat in fiecare celula rezultata se gaseste doar o singura unitate functionala, cromatida. Structura redusa (1n) se numeste haploida. Prin unirea gametilor care sunt purtatori de 1n cromozomi in timpul fecundatiei, se reconstituie structura pereche de 2n cromozomi care contine bagajul genetic nou, amestecat matern si patern. Gametogeneza cuprinde o linie masculina in urma careia sunt generati spermatozoizii (are loc in organismul masculin, in testicul) si o linie feminina in urma careia sunt generate ovule (in organismul feminin, in ovar). Celula umana contine ca bagaj genetic 22 perechi de cromozomi somatici 2n(autozomi) care transmit caractere morfologice somatice si o pereche de cromozomi sexuali numiti gonozomi care determina sexul genetic al individului. Meioza sau diviziunea reductionala reprezinta totalitatea fenomenelor premergatoare formarii gametilor (spermatozoid si ovul). Acestea au loc doar in celulele germinale ale tuturor organismelor cu reprpducere sexuata. Formarea gametilor este precedata de doua diviziuni nucleare succesive, una reductionala (sau primara indicata I), si una normala mitotica (sau secundara, indicata II). Prima diviziune se numeste si heterotipica deoarece din celule diploide cu doua garnituri de cromozomi (2n) una de provenienta materna, alta paterna, iau nastere doi nuclei haploizi (n). A doua diviziune de maturatie sau homotipica este asemanatoare mitozei obisnuite. In aceasta diviziune are loc separarea cromatidelor in fiecare din cei doi nuclei haploizi, ceea ce determina in final producerea a 4 nuclei haploizi care in urma unor procese de maturare dau nastere gametilor. Mitoza este un proces complex prin care nucleul celular se divide. Are loc in tesuturile aflate in cretere. Aceasta diviziune asigura cresterea si diferentierea individuala, precum si continuitatea genotipului si evolutia speciei. Diviziunea mitotica se petrece in celulele-mama diploide (2n crz.). In urma diviziunii, celula se duplica, luind nastere doua celule identice in continut cu celulamama initiala, deci celulele fiice vor fi tot diploide (2n). TESUTUL EPITELIAL acopera intreaga suprafata a corpului si margineste cele mai multe cavitati ale organismului (cavitatea peritoneala, pleurala dar nu si spatiile articulare sinoviale). Tesutul epitelial formeaza si glandele exo si endocrine. Majoritatea glandelor exocrine se asociaza sistemului digestiv. Caracterele generale ale epiteliului: 1. Are origine embriologica din oricare din cele trei foite germinale: epidermul se formeaza din ectoderm, endoteliul vascular din mesoderm, epiteliul tractului gastrointestinal dn endoderm. 2. Epiteliul este un tesut bogat celular, cu spatiu extracelular minim. Jonctiunile intercelulare sunt bine dezvoltate si mentin apropierea dintre celulele epiteliului.
3. Absenta vaselor sangvine-Epiteliul este avascular, vasele se afla in apropiere dar totdeauna separate de celulele epiteliale de un tesut conjunctiv perivascular. 4. Polarizarea. Atat celulele epiteliale cat si tesutul epitelial sut polarizate. -suprafetele celulare o celula epiteliala a unui epiteliu simplu are trei suprafete distincte: suprafata apicala care este suprafata libera ce delimiteaza un spatiu, suprafata laterala care priveste celulele epiteliale vecine, suprafata bazala ce priveste tesutul conjunctiv al epiteliului, de obicei atasata de o lama bazala. -domeniile celulare domeniul apical catre suprafata, domeniul bazo-lateral ca un domeniu secundar ca functie. -suprafetele tisulare suprafata libera, ce priveste fie catre spatiul exterior organismului, fie catre spatiile interne izolate de exteriorul corpului (cavitate peritoneala, pleurala, lumenul sistemului cardio-vascular). -suprafata bazala, adiacenta tesutului conjunctiv. Functiile epiteliului 1. Absorbtia. Multe epitelii sunt localizate la interfata dintre corp si mediul extern sau intern. Solventii, solutiile, substantele metabolice, gazele sunt absorbite pasiv sau activ prin epitelii. Exemple de epitelii absorbante sau transportoare sunt: epiteliul intestinal, epiteliul pulmonar alveolar, endoteliul vascular, epiteliul tubular renal. 2. Protectia. Epiteliile sunt specializate sa protejeze si sa mentina compozitia mediului intern. Un asemenea epiteliu prezinta fata de mediul extern ostil, o bariera celulara continua de aparare. Un epiteliu poate prezenta catre mediul fata de care se delimiteaza o lama de celule scuamoase extensibile. Un exemplu ar fi epidermul sau epiteliul colonului. 3. Secretia. -glandele unicelulare o celula epiteliala individuala poate avea structura secretorie, si de obicei astfel de celule sunt inconjurate de celule epiteliale nonsecretoare. -suprafatele secretorii sunt formate din multiple celule secretorii. -glande multicelulare atat exocrine cat si endocrine se dezvolta din invaginatii ale epiteliului de suprafata. In glandele exocrine caracteristicile epiteliale sunt vizibile atat in regiunile secretorii cat si in ductele ce leaga glandele de o suprafata epiteliala. In glandele endocrine portiunea initiala de duct ale invaginatiilor embriologice se pierde iar structura epiteliala de glanda adulta poate fi mai putin evidenta. Clasificarea epiteliilor se face dupa forma celulei si numarul de straturi celulare. Dupa forma celulelor: 1. epiteliul scuamos contine celule turtite, la care inaltimea este mult mai mica decat lotimea. 2. epiteliul cuboidal contine celule cuboide la care inaltimea si latimea sint apropiate. 3. epiteliul columnar contine celule inalte la care inaltimea este mult mai mare ca latimea. Dupa numarul de straturi celulare: 1. epiteliu simplu, format dintr-o singura lama de celule 2. epiteliu stratificat format din doua sau mai multe straturi celulare.In descrierea tipului de tesut epitelial stratificat doar forma celulelor de la suprafata este luata in considerare in denumire. 3. epiteliu pseudostratificat este un aranjament de celule cuboidale si columnare in care toate celulele iau contact cu membrana bazala dar, doar celulele columnare ajung la suprafata libera (traheea).
4. epiteliul de tranzitie este un aranjament celular care se adapteaza suprafetelor care isi schimba forma (vezica, ureterele). In epiteliulde tranzitie celulele pot apare turtite atunci cand organul este destins sau groase cand organul in cauza este golit. Toate celulele unui epiteliu de tranzitie contacteaza membrana bazala dar aceasta nu se vede totdeauna in sectiunile obisnuite. Diviziunea mitotica la nivel de epiteliu si reinnoirea tesutului. O caracteristica generala este inlocuirea graduala a tesutului dupa o perioada de timp. Acest timp variaza de la cateva zile in cazul epiteliului intstinal la o luna in cazul epidermului. Compartimentul mitotic se restrange la regiuni ale epiteliului, lama bazala. Aici sunt prezente celule stem ramase in portiuni restranse Celulele pot trece prin mai multe diviziuni pe masura ce acestea migreaza catre spatiul de diferentiere postmitotica. Compartimentul postmitotic ia nastere prin migrarea dinspre compartimentul mitotic catre stratul de diferentiere postmitotica. In ciclul de reinnoire a tesutului epitelial se trece prin urmatoarele stadii: diviziune celulara, migrare, diferentiere, senescenta (imbatrinire), moarte celulara. TESUTUL CONJUNCTIV Tesutul conjunctiv inconjoara diverse tipuri de tesuturi cu structuri diferite si functii diferite. Tesutul conjunctiv este caracterizat printr-un numar relativ mic de celule intr-un volum mare de matrice extracelulara care include fibre, substanta de baza si lichid tisular. Fibrele tesutului conjunctiv sunt de trei feluri : colagen, fibre reticulare, fibre elastice, toate sintetizate de celule fibroblaste. 1. COLAGENUL este o familie de fibre glicoproteice ce constituie majoritatea componentei fibroase ale tesutului conjunctiv. Colagenul formeaza fibrele albe ale tesutului conjunctiv lax si este componenta mejora din tendoane, ligamente, oase si stroma corneei. Fibrele colagene (care au o putere de rezistenta la intindere echivalenta cu cea a cablului de otel) sunt fascicule de fibrile colagene puternic flexibile dar inelastice. -fibrele colagene apar la examinarea microscopica, optica ca structuri sub forma de val de diferite grosimi si lungime nedeterminata (de coloratie rosiatica). -fibrilele colagene apar la microscopia electronica ca un manunchi de subunitati fibrilare ale unei fibre colagene. Fibrilele individuale au o lungime nedeterminata si o grosime diferita in functie de tesutul din care fac parte. -tropocolagenul este unitatea structurala de baza a unei unitati colagene. Tropocolagenul este o molecula cu polaritate axiala careia i se identifica cap si coada. Molecula de tropocolagen este compusa din trei lanturi polipeptidice rasucite sub forma unui triplu helix rasucit la dreapta. Moleculele de tropocolagen se asambleaza longitudinal si lateral dupa un model precis. Moleculele se aliniaza cap la coada intr-un aranjament linear ce formeaza un rand, iar randurile se aranjeaza lateral. Intinderea fibrilei si prin aceasra a fibrei se datoreaza legaturilor covalente dintre moleculele de tropocolagen din randurile adiacente. Procolagenul, precursorul moleculei de colagen este sintetizat in interiorul celulei. Pe masura ce procolagenul este eliberat in matricea extracelulara, procolagenul este
convertit in tropocolagen. Colagenul tesutului conjunctiv este sintetizat si secretat in principal de catre fibroblaste, dar si alte tipuri de celule sintetizeaza colagen. 1. Sinteza procolagenului: -lanturile polipeptidice sunt produse de poliribozomii reticulului endoplasmatic rugos, iar noile lanturi sintetizate sunt eliberate in cisternele acestei retele endoplamatice. -modificarile postsintetice apar atat din cisternele reticulului endoplasmatic rugos cat si complexelor Golgi. Cand lanturile polipeptidice nu sunt inca helicoidale prolina si lisina din acestea sunt hidroxilate. Vitamina C este coenzima esentiala pentru aceasta hidroxilare si in lipsa ei legaturile de H esentiale pentru realizarea triplu helixurilor nu se pot forma. In scorbut, (deficitul sever de vit C) structura normala a moleculelor de colagen nu mai este respectata. Ranile nu se mai vindeca, formarea osului este deficitara, dantura devine laxa, si chiar rani vechi si vindecate se pot deschide. Se formeaza structura de triplu helix din lanturile polipeptidice rasucite cu exceptia capetelor, care raman nerasucite.Apar legaturile de H intralant si interlanturi polipeptidice care dau forma finala si stabilizeaza molecula finala. -procolagenul paraseste celula prin exocitoza veziculelor secretorii. 2. Sinteza tropocolagenului. Pe masura ce veziculele ce contin procolagen fuzioneaza cu membrana plasmatica o procolagen peptidaza cliveaza portiuni din terminatiile nonhelicoidale ale moleculei transformand-o in tropocolagen. 3. Asamblarea fibrilelor de colagen. Celulele pot controla aranjamentul noilor fibrile colagene directionand veziculele secretorii in portiuni localizate ale membranei plasmatice pentru exocitoza dirijata. Lanturile care formeaza triplu helix al colagenului variaza din punctul de vedere al secventei aminoacide astfel incat por finaliza 16 tipuri de colagen. -tipul I colagen este cel mai raspandit si apare in tesutul conjunctiv lax si dens. Doua lanturi sunt identice si unul diferit. -tipul II colagen apare ca fibrile foarte fine. -tipul III colagen contine trei lanturi identice. -tipul IV colagen nu formeaza fibrile ordonate, si este tipul constituent la membranelor bazale. Fibrele reticulare Fibrele reticulare ofera un suport in retea pentru constituentii celulari ai variatelor tipuri de tesuturi si organe. Fibrele reticulare formeaza o retea laxa in jurul fibrelor musculare, nervoase, vaselor de sange, organe glandulare sau tubulare ca si lama bazala a epiteliilor. Formeaza de asemenea reteaua structurala a organelor hematopoietice si limfatice. Fibrele reticulare sunt compuse din tipul III de colagen si sunt cele mai mici fibre din tesutul conjunctiv. Fibrele reticulare sunt sintetizate de cateva tipuri de celule din organism: -fibroblastele -celulele reticulare din maduva osoasa, splina, nodulii limfatici. -celulele Schwann ai nervilor periferici. -celulele musculare netede ale vaselor de sange si din tractul digestiv. Fibrele elastice
Ofera mecanismul structural ce permite tesuturilor sa raspunda la intindere si distensie. Fibrele elastice si lamelele elastice sunt componenta majora a peretilor arteriali, ligamentelor elastice, bronhiolelor, pielii, cartilajelor elastice. Fibrele elastice sunt fibrele galbene puternic retractile ale tesutului conjunctiv lax. Lamelele elastice sunt straturi nefibroase de elastina. Fibrele elastice sunt mult mai subtiri decat fibrele colagene, nedeterminate in lungime, aranjate dupa un schelet ramificat ce formeaza o retea tridimensionala. Fibrele elastice sunt in general invaluite cu fibre colagene. Aceasta limiteaza distensia tesutului si previne ruperea cauzata de inteinderea excesiva. Fibrele elastice sunt compuse din doua elemente structurale, elastina si microfibrilele care isi schimba proportia pe masura ce cresc. Elastina este o molecula globulara. Microfibrilele contin glicoproteine fibrilare si sunt relativ drepte si inguste. Ele inconjoara elastina fibrei elastice in crestere, iar unele devin prinse in fibra pe masura ce aceasta creste. Fibrele elastice si lamelele sunt produse de fibroblaste si celulele musculare netede: -elastina este secretata pe aceeasi cale ca si colagenul. Ambele procese apar in mod normal simultan in fibroblaste si alte celule ce secreta ambele molecule impreuna cu alte proteine, glicoproteine si componente carbohidrate ale tesutului conjunctiv. -tropoelastina. Elastina este secretata de celule ca lanturi de tropoelastina. In matricea extracelulara adiacenta, moleculele de tropoelastina agrega si formeaza legaturi incrucisate pentru a forma fibrele elastice. Legaturile covalente de legatura intrre lanturile polipeptidice de elastina sunt produse de desmozina si isodesmozina, doi aminoacizi unici ai elastinei. SUBSTANTA DE BAZA A TESUTULUI CONJUNCTIV Este compoenenta tesutului conjunctiv care umple spatiul neocupat de celule si fibre. In vivo, substanta de baza este vascoasa, clara, alunecoasa. Are un bogat continut de apa, care combinat aparent cu lipsa de structura specifica, ofera putine date de caracterizare morfologica. Din punt de vedere chimic, substanta de baza consta din complexe de glicozaminoglicani, proteoglicani, acid hialuronic, glicoproteine. Lichidul tisular din substanta de baza ofera mediul prin care au loc schimburile intre sange, limfa si celulele tisulare. Intregul substrat metabolic si rezidurile ce se schimba intre sistemul vascular si celulele tesuturilor si organelor trebuie sa difuzeze in lichidul tisular al substantei de baza a tesutului conjunctiv. Substantele absorbite de epiteliul gastrointestinal trebuie sa traverseze substanta de baza pentru a intra in sistemul vascular. Miscarea lichidului dinspre sistemul vascular catre substanta de baza a tesutului conjunctiv si retinerea ei aici este responsabila de edemul tisular. CELULELE TESUTULUI CONJUNCTIV Pot fi descrise ca celule rezidente, ce apartin tesutului, sau celulele de tranzitare. Tipul celulelor si numarul lor reflecta statusul functional al tesutului. Celulele rezidente sunt: fibroblastele, macrofagele, mastocitele, celulele adipoase, celulele mesenchimale nediferentiate.
1. FIBROBLASTELE sunt celulele specifice tesutului conjunctiv, stabile, care au o mica misscare celulara, fiind considerate permanente. Fibroblastele sunt principala sursa de secretie a colagenului, se afla de obicei in apropierea fibrelor de colagen pe care le-a secretat si sunt inconjurate de substanta de baza secretata tot de ele. Desi toate fibroblastele sunt capabile sa se divida si sa repare rani, fibroblastele din unele localizari constituie populatii celulare de replicare regulata. Fibroblastele endometrului (mucoasa) uterin prolifereaza si diferentiaza, degenereaza in fiecare ciclu mestrual sub controlul hormonilor steroizi. Fibroblastele situate imediat sub epiteliul intestinal se inmultesc, migreaza si se diferentiaza in paralel si sincron cu migratia celulelor epiteliale.Fibroblastele imediat de sub epiderma se inmultesc si migreaza in interiorul dermului profund. Miofibroblastele detin atat proprietati specifice fibroblastelor cat si ale celulelor musculare netede. Contin manunchiuri de microfilamente de actina si corpi densi specifici citoplasmei celulelor musculare. 2. MACROFAGELE sunt celule fagocitice iar in tesutul conjunctiv sunt numite si histiocite. Ele raspund provocarilor date de organisme si materiale straine, expunerii la exterior al tesuturilor traumatizate prin fagocitoza. Materialele prea mari pentru a fi fagocitate pot fi izolate prin formarea celulelor gigante sau modulatiei epiteloide. Fagocitoza implica de regula legatura efector-receptor la suprafata externa a membranei plasmatice a macrofagului. Materialul de ingerat este initial invaginat printr-o inchidere ca fermoarul oferita de membrana plasmatica in jurul acelui material. Macrofagele sunt invelite cu receptori de anticorpi. Particulele straine, organismele si moleculele acoperite cu anticorpi, se leaga de suprafata macrofagelor si induc fagocitoza. Vacuolele ce contin materialul ingerat pot fuza cu lizozomii pentru a forma fagolizozomi sau pot fuza cu lizozomi preexistenti in care materialul este digerat de enzime lizozomale. Formarea celulelor gigante. Unele organisme patogene si alti corpi straini sunt ingerati dar nu pot fi digerati de macrofage. Aceste cazuri se refera la bacilii tuberculozei, leprei, Toxoplasma, la particule de poluare industriala ca azbestul, praful de carbune, fibrele de bumbac. In aceste cazuri, macrofagele de obicei fuzioneaza pentru a forma celule gigante straine, multinucleate numite si Langhans. Modularea epiteloida . Macrofagele pot forma si jonctiuni pentru a se asambla in straturi de celule ce seamana epiteliului. Astfel de celule epiteloide adesea marginesc locuri de inflamatie cronica sau infectie. Macrofagele sunt esentiale in recunoasterea de catre un organism a self-ului, nonself-ului si self-ului modificat. Recunoasterea non self-ului este esentiala in indepartarea organismelor patogene, materialelor toxice. Recunoasterea self-ului modificat este esentiala in indepartarea celulelor transformate (tumorale) cu origine in organism. Macrofagele participa la raspunsul imun ca celule ce prezinta antigenul, dar si prin rolurile sale directe sau indirecte in diferentierea si functionarea limfocitelor. Macrofagele leaga, captureaza antigene pe suprafata lor, sau pot ingera, procesa si resecreta antigeni pe suprafata lor. Antigenii sunt apoi prezentati limfocitelor initiind proliferarea limfocitelor T sau B prin procesul de transformare blastica.
10
Macrofagele secreta agenti imunostimulatori (limfokine) care stimuleaza raspunsul imun. Printre acestia sunt interleukina 1, 6. Macrofagele pot sa secrete si factori ce pot actiona ca agenti imunosupresivi. Macrofagele pot fi activate de produsele de secretie ale limfocitelor. Macrofagele sunt esentiale in procesele de inflamatie si vindecare a ranilor: -maresc raspunsul inflamator la leziune prin secretarea factorilor ce atrag polimorfonucleare si monocite la locul respectiv. -fagociteaza si digera bacteriile, celulele lezate, polimorfonuclearele epuizate, fibrina, matricea extracelulara. -activitatea macrofagelor este esentiala in stimularea si proliferarea fibroblastelor se celulelor endoteliale la locul inflamatiei. -daca inflamatia sau activarea macrofagelor este prevenita cum se intampla in administrarea de corticosteroizi, proliferarea fibroblastelor si endeliului sunt mult incetinite , iar vindecarea ranii oprita. MASTOCITELE sunt de obicei gasite in apropierea vaselor de sange. Mastocitele sunt similare dar nu inrudite cu celulele bazofile ale sangelui. Mastocitele contin unele substante vasoactive si imunoreactive. Histamina este un agent vasoactiv care se elibereaza din celula impreuna cu substantele lent reactive ale anafilaxiei (slow-reacting substance of anaphylaxis SRSA), care nu sunt inmagazinate in granule si cresc permeablitatea vaselor vecine cu aparitia edemului in tesutul inconjurator. Heparina este un anticoagulant si un agent antilipemic. Factorul chemotactic al eozinofilelor este de asemenea continut in mastocite. Acesta stimuleaza migrarea oezinofilelor la locul de eliberare a acestor substante.Secretiile eozinofilelor contracaeaza stimulii de inflamatie acuta dati de histamina si substantele lent reactive ale anafilaxiei (SRS-A). Eliberarea granulelor mastocitelor este initiata de legarea unui antigen la imunoglobulina specifica tip E de pe suprafata mastocitului. Degranularea mastocitului prin fuziunea membranei granulare cu membrana plasmatica, ceea ce produce canale intracelulare ce faciliteaza eliberarea rapida a continutului granulos in matricea tesutului conjunctiv. Anticorpii legati de suprafata mastocitului sunt produsi de celulele plasmatice ce au fost stimulate de prezenta unui antigen sa sintetizeze si sa elibereze imunoglobuline in tesutul conjunctiv. Acesta este procesul de sensibilizare. In timp ce unele clase de imunoglobuline sunt produse de celulele plasmatice, doar imunoglobulinele E, specifice unor antigeni individuali se leaga de receptorii membranei plasmatice ai mastocitelor. La o urmatoare expunere la acelasi antigen ce a determinar producerea de IgE , apare o reactie antigen-anticorp ce determina descarcarea masiva a granulelor celulelor mastocite. Mastocitele nu se divid in tesutul conjunctiv, au o viata lunga si sunt considerate a fi celule rezidente ale tesutului conjunctiv. Mastocitele degranulate sintetizeaza noi produsi de secretie sub forma de granule. CELULELE ADIPOASE sunt specializate pentru depozitarea grasimilor neutre (trigliceride) si pot fi prezente in numar mare in tesutul conjunctiv lax. CELULELE MEZENCHIMALE NEDIFERENTIATE persista in tesutul conjunctiv aal adultilor. Aceste celule sunt asociate cu tunica cea mai externa a venulelor
11
si cu tesutul conjunctiv mai putin organizat din jurul capilarelor venoase si venulelor postcapilare. Acestea au rol in procesele de vindecare a leziunilor. Populatia de celule tranzitorii La locul unei inflamatii acute sau cronice, unele tipuri de celule pot parasi circulatia si sa intre in tesutul conjunctiv lax, unde exista tranzitor. Acestea sunt leucocitele primare ce cuprind limfocite, monocite, neutrofile, eozinofile si bazofile. Celulele plasmatice ce deriva din limfocitele B sunt si ele parte din pupalatia celulara tranzitorie. 1. Limfocitele sunt implicate in raspunsul imun. Sunt cele mai mici celule libere din tesutul conjunctiv. Un numar mic de limfocite T si B se afla in tesutul conjunctiv lax in intreg organismul. Numarul lor creste puternic la locul infectiei sau inflamatiei. Limfocitele sunt numeroase in tractul respirator, alimentar, genito-urinar unde au rol esential in imunosupraveghere fata de organismele patogene si substantele straine ce intra in organism prin aceste cai. 2. Celulele plasmatice sunt producatoare de anticorpi derivate din limfocitele B, gasite in tesutul conjunctiv lax si organele limfatice. 3. Neutrofile,eozinofile, bazofile, monocite sunt de asemenea observate in tesutul conjunctiv lax si reflecta o reactie acuta inflamatorie. -neutrofilele sunt caracterizate de un nucleu multilobat, si migreaza din circulatie catre locul injuriei sau infectiei. -eozinofilele sunt caracterizate de prezenta granulelor citoplasmatice si nucleul bilobat, de asemenea parasesc circulatia catre locul injuriei sau inflamatiei . -bazofilele seamana cu celulele mastociteca functie , continut granular, acoperire cu IgE, dar parasesc circulatia catre tesutul conjunctiv. -monocitele parasesc circulatia catre tesutul conjunctiv unde se diferentiaza in macrofage. Clasificarea testutului conjunctiv Se face pe baza tipurilor de celule prezente, tipului de fibre si caracterul substantei de baza.Tesuturile conjunctive se clasifica in : conjunctiv propriu-zis, conjunctiv specializat, conjunctiv embrionic. Tesutul conjunctiv propriu-zis se imparte in lax si dens. Tesutul conjunctiv lax se mai numeste si areolar este caracterizat de un aranjament larg al fibrelor, abundenta de substanta de baza si de prezenta diferitelor celule rezidente si tranzitorii. Se gaseste majoritatea organelor, in mezentere, fascia superficiala, tesutul inconjurator al majoritatii vaselor de sange, nervi si celule musculare. Difuziunea oxigenului , substratului metabolic si resturilor ca si a agentilor de reglare dintre vase si celule are loc prin substanta de baza a tesutului conjunctiv lax. Celulele adipoase sunt prezente in grade diferite. Cand aceste celule predomina, tesutul este numit adipos. Tesutul conjunctiv lax dubleaza epiteliul ce acopera corpul. Acesta este locul unde antigenele sau alte substante straine ca bacteriile pot fi detectate si distruse de sistemul imun. Tesutul conjunctiv dens este mai putin celular decat cel lax si are mai multe fibre groase. Poate fi: -dens neregular in care fibrele sunt valurite in dermul profund, periost, submucoasa canalului alimentar.
12
-dens regular de obicei aranjat in cordoane sau benzi de largimi diferite.Fibrele sunt aranjate paralel ca in tendoane, ligamente, aponevroze, stroma corneei. Tesutul conjunctiv elastic si reticular In trecut tesutul elastic si reticular erau considerate entitati separate ale tesutului conjunctiv spacializat. Acum sunt considerate a fi ca varietati ale tesutului conjunctiv propriu-zis. Tesutul elastic include ligamentele elastice asociate cu coloana vertebrala si tunica medie a arterelor mari. Tesutul reticular este caracteristic tesutului hematopoietic din maduva rosie a oaselor si a splinei, a nodulilor limfatici. Tesutul conjunctiv specializat include acele forme ale tesutului distincte de cel propriu-zis atat in structura cat si functie.Acestea sunt tesutul adipos, cartiilagiul, osul, sangele, tesutul hematopoietic si limfatic. TESUTUL MUSCULAR Functia primara a tsutului muscular este contractia in sensul exercitarii lucrului mecanic. Celulele tesutului muscular sunt specializate pentru contractie prin aranjamentul regulat al majoritatii proteinelor contractile care sunt actina si miozina. Aranjamentul acestor elemente contractile permite clasificarea histologica a muschilor in doua categorii: striat care include muschii scheletici si cardiac si tesut muscular neted. Proteinele contractile: actina si miozina sunt proteinele contractile ale tesutului muscular striat si neted. Actina F este un polimer helicoid format din molecule globulare de actina G monomer, care se asambleaza in filamente lungi, flexibile, subtiri. Actina G monomer are doua locusuri de legare: -unul ocupat de ATP (adenozin-tri-fosfat) sau ADP (adenozin-di-fosfat) necesare asamblarii actinei G in forma filamentara F. -un locus pentru miozina care functioneaza in crearea legaturilor necesare pentru contractie. Miozina este o molecula mare proteica cu o portiune filamentara sub forma de coada si o portiune cu doua capete globulare. Portiunea capului este o ATP-aza actin activata capabila sa activeze miscarea pe seama ATP-ului. Portiunea cozii regleaza asamblarea moleculelor de miozina in filamente groase caracteristice celulelor musculare striate. Unele molecule de miozina se asambleaza coada la coada pentru a produce un complex molecular bipolar, avand zona H la mijloc. Capetele se proiecteaza in afara filamentului gros. Zona centrala in care nu sunt prezente capete formeaza zona H. Clasificarea tesutului muscular: striat si neted. Muschiul striat prezinta striatii transverve la microscopia optica. Dupa localizare muschiul striat poate fi scheletal si cardiac. Muschiul scheletal este atasat oaselor si responsabil pentru miscarea scheletului. Muschii strati viscerali sunt identici cu cei scheletali , dar restransi la zone bine delimitate si tesuturi moi: limba, faringe, diafragm, portiunea superioara a esofagului. Muschiul cardiac formeaza miocardul.
13
Muschiul neted nu prezinta striatii transverse. Este muschiul intrinsec al canalului alimentar, vaselor de sange, tractului genitourinar, respirator si alte organe tubulare. Muschiul neted se organizeaza si in iris corpii ciliari ai ochiului, ca fibre izolate in legatura cu foliculii pilosi. Muschiul scheletic consta in manunchiuri de fibre musculare striate numite fascicule tinute impreuna de tesutul conjunctiv si legate prin tesutul conjunctiv la os. Muschiul este responsabil de miscarile mari si fine ale membrelor, degetelor, mentinerea pozitiei corpului, expresia faciala, miscarile de precizie ale ochilor. Caracterisici generale ale fibrei musculare scheletice: -fibrele se formeaza in cursul dezvoltarii embionare prin fuziunea unor celule mici individuale numite mioblaste. -miofibrila este principala subunitate structurala si functionala a fibrei musculare. Miofibrilele sunt compuse din manunchiuri de filamente de actina si miozina. Fiecare miofibrila este inconjurata de un reticul endoplasmatic lax specializat impreuna cu mitocondii si glicogen care ofera energia necesara contractiei. -citoplasma este numita sarcoplasma, iar membrana plsmatica se numeste sarcolema. Tesutul conjunctiv asociat muschiului scheletic cuprinde: -endomisium- o lama delicata de fibre reticulare si tesut cinjunctiv lax care inconjoara fiecare fibra musculara. In acest tesut conjunctiv se afla capilare sangvine, ramuri fine ale nervilor periferice. -perimisium- o lama mai groasa de tesut conjunctiv care inconjoara un grup de fibre musculare ce formeaza un manunchi sau un fascicul de fibre. In perimisium se afla vase si nervi mai mari. -epimisium- este partea cea mai externa a tesutului conjunctiv dens care inconjoara totalitatea fasciculelor unui muschi. La acest nivel se afla pediculii vasculari si inervatia majora. Epimisiumul este fascia observata in cursul disectiei. -jonctiunea cu tendonul- este un tesut conjunctiv care inconjoara fibrele musculare si fasciculele si se continua cu tendoanele si aponevrozele oferind astfel legatura cu scheletul. Caracteristile histologice ale muschiului scheletal Striatiile transversale rezulta din aranjamentul regulat al elementelor contractile de actina si miozina in interiorul citoplasmei celulelor. Striatiile transversale sunt observate in coloratia cu hematoxilina-eozina prin alternanta benzilor intunecate cu a celor clare. Benzile intunecate sau A se numesc asa deoarece apar anizotropice. Benzile clare sau I se numesc asa deoarece sunt isotropice. Fiecare dintre benzile A sau I sunt intersectate de o zona ingusta densa: -discul Z este sona densa care intersecteaza banda I -zona H este o zona clara care imparte banda A. In preparatul bine realizat se observa si o linie M care imparte zona H. Sarcomerul este segmentul de miofibrila intre doua discuri Z adiacente. Aceasta este unitatea contractila de baza a muschiului striat. Reticulul sarcoplasmatic este aranjat in retele in jurul manunchiurilor de miofilamente definind astfel miofibrilele. Reticulul sarcoplasmatic se situeaza ca un manson de tubuli aplatizati ce inconjoara scurte portiuni din miofibrila de la o jonctiune
14
de banda A-I la alta. Capatul fiecarui mansaon formeaza compartimente numite cisterne terminale care inmagazineaza ioni de calciu. Tubulii transversi sau sistemul T sunt invaginatii ale membranei plasmatice care penetreaza adanc in fibra musculara. Tubulii T se extind intre cisternele terminale ale reticulului sarcoplasmatic. Asocierea dintre un tubul T si doua cisterne de fiecare parte formeaza o triada musculara. Mecanismul contractiei muschiului striat modelul alunecarii filamentelor. Cand muschiul se contracta, fiecare sarcomer se scurteaza, devine mai gros, dar miofilamentele raman de aceeasi lungime. 1. Superpozitia filamentelor. Pentru a initia scurtarea, trebuie sa se mareasca gradul de suprapunere al miofilamentelor, adica filamentele groase si subtiri trebuie sa gliseze unele peste altele. -banda I se scurteaza in lungime in timp ce banda A ramane de aceeasi lungime. -zona H devine mai groasa pe masura ce filamentele fine patrund in zona H. -filamentele subtiri gliseaza peste filamentele groase in timpul contractiei. 2. Interactiunea dintre filamentele de miozina si actina in timpul contractiei. Un cap de miozina se leaga de un filament adiacent de actina si o schimbare de conformatie a capului miozinic gliseaza miozina de-alungul actinei. O interactiune ATP dependenta intre capul miozinic si filamentul actinic adiacent genereaza o forta care produce glisarea celor doua filamente unul de-alungul celuilalt. Fibrele groase de miozina sunt bipolare astfel aceeasi interactiune dintre capetele miozinice si filamentele de actina apar la fiecare capat de filament. Discurile Z dintr-un sarcomer sunt impinse unul catre celalat rezultand o scurtare a sarcomerului. 3. Reglarea contractiei. Este o functie de reglare a interactiunii miozina-actina. Aceasta depinde de prezenta proteinelor reglatoare tropomiozina si troponina care sunt asociate cu actina F si de disponibilitatea ionilor de calciu in sarcoplasma. Tropomiozina este o proteina atasata actinai F. Troponina este un complex de trei molecule . Cand toate trei subunitatile de troponina sunt prezente ca un complex, functiile lor combinate devin sensibile la Ca. Cand Ca se leaga de troponina, conformatia complexului se schimba si permite moleculelor de tropomiozina sa isi schimbe pozitia astfel incat capetele de miozina sa lege filamentele de actina. Ca este sechestrat in cisternele terminale ale reticulului sarcoplasmic. Ca urmare a stimularii fibrei musculare de inervatia motorie, calciu stocat se elibereaza in sarcoplasma inconjuratoare, se leaga la troponina si initiaza contractia. Cand stimularea nervoasa inceteaza calciu se regrupeaza in reticulul sarcoplasmic. Jonctiunea neuromusculara este asocierea functionala dintre sistemul nervos si tesutul muscular scheletal. Muschiul striat scheletic este bogat inervat de neuroni motori cu localizare in maduva spinarii sau creier.Axonii neuronilor motori din apropierea celulei musculare se ramifica si trimit terminatii fine ce se distribuie individual fiecarei celule musculare cheletale. La loculde contact teaca de mielina a terminatiei nervoase se termina. Terminatiile axonice se impart in ramificatii fine ce se plaseaza pe depresiuni fine pe suprafata fiecarei fibre musculare. Terminatiile axonice contin numeroase mitocondrii si vezicule sinaptice ce contin acetilcolina ca neurotransmitator. Terminatia axonica si
15
membrana celulei sunt separate de o fanta sinaptica. Membrana celulei musculare care participa la fanta sinaptica are numeroase pliuri jonctionale adanci. Receptorii pentru acetilcolina se afla pe membrana celulei musculare in varful acestor pliuri. Acetilcolinesteraza (enzima ce neutralizeaza acetilcolina si deci actiunea ei) este prezenta de-alungul fantei sinaptice si a pliurilor membranare. Initierea contractiei este dictata de activitatea sinaptica a neuronului motor. Potentialul de actiune membranar la nivelul terminatiei axonice soseste in spatiul presinaptic si induce eliberarea acetilcolinei de catre neuron. Acetilcolina difuzeaza prin fanta sinaptica si se leaga de receptorii de acetilcolina de pe membrana plasmatica a celulei musculare la varful pliurilor postsinaptice. Depolarizarea acestei membrane urmeaza legarii acetilcolinei de receptorii sai. Valul de depolarizare se propaga dealungul intregii suprafete a fibrei musculare si este condus adanc in interiorul acesteia prin tubulii T. Pentru a preveni stimularea continua, acetilcolina este rapid indepartata din sinapsa prin difuzare si inactivare de catre acetilcolinesteraza. O singura unitate motorie poate inerva doar cateva fibre musculare. Neuronul motor si toate fibrele musculare inervate de acesta definesc unitatea motorie. Muschiul cardiac (miocardul) este muschiul striat al inimii. Poate fi prezent si in venele mari din vecinatatea inimii. Tesutul muscular cardiac consta din celule unice, mononucleate organizate ca fibre lungi care se ramifica si anastomozeaza cu fibrele invecinate. Deoarece muschiul cardiac are acelasi mod de aranjare a elementelor contractile ca si muschiul scheletic, prezinta striatii transversale. Fibrele musculare se unesc una cu alta dupa o distributie lineara prin jonctiuni intercelulare specializate numite discuri intercalare. Lama aderenta este componeneta majora a acestei portiuni transverse. Lamele aderente servesc la legarea celulelor musculare cardiace una la ealalta la capetele libere pentru a forma fibra. Filamentele de actina din capetele terminale ancoreaza in materialul dens al membranei plasmatice. Golurile jonctionale sunt specifice portiunilor laterale ale celulalor cardiace. Acestea furnixeaza o cale pentru conducerea ionica intre celulele adiacente, ceea ce permite tuturor celulelor cardiace adiacente sa primeasca semnalele contractile una de la alta. Acest aranjament in care toate celulele musculare cardiace sunt cuplate electric intre ele, le permite sa se comporte ca un sincitiu functional in timp ce isi pastreaza integritatea celulara. Nucleul celular este unic in centrul celulei, miofibrilele trec in jurul nucleului. Regiunea perinucleara este bogata in organite celulare; mitocondrii, aparaat Golgi, granule de glicogen,etc...Miofilamentele celulelor cardiace sunt organizate in sarcomere ca si celulele musculare scheletale. Toate celulele musculare cardiace au un ritm intrinsec de activitate coordonat de celule specializate, modificate ale muschiului cardiac care formeaza tesutul excitoconductor. Aceste celule sunt organizate in noduli si fascicule care transmit impulsul contractil la diferite parti ale miocardului in secventa proprie. Muschiul neted apare la modul general ca buchete sau lame de celule fusiforme alungite. Este specializat in contractia lenta, prelungita.
16
Celulele musculare netede nu contin un aranjament inalt ordonat al filamentelor contractile subtiri si groase, astfel incat nu apare striat. Celulele au nucleu unic localizat central. Organitele celulare sunt concentrate la oricare pol nuclear. Restul citoplasmei este plin cu filamente de actina si miozina. Filamentele de actina sunt similare cu actina celulei musculare striate, se aliniaza de-alungul celulei luand contact cu corpii densi care sunt tot aglomerari de filamente actinice.Miozina este similara structural cu cea a muschiului striat si se dispune in filamente groase. Muschiul nedet nu are sistem T. Contractia muschiului neted depinde de Ca si miozina. Interactiunea dintre capetele miozinice si actina este dependenta de un ciclu de fosforilare-defosforilare a lanturilor miozinice. Ca urmare a cresterii concentratiei de Ca intracelular este activata o enzima calmodulina care catalizeaza fosforilarea miozinei in filamente groase care pot interactiona cu filamentele de actina pentru a initia contractia. Fosforilarea apare lent, iar contractiei ii ia de multe ori secunde pentru a atinge nivelul maxim. Astfel contractia este lenta si poate fi prelungita. Defosforilarea miozinei duce la disocierea actinei de miozina si la relaxare musculara. Nu exista jonctiuni neuromusculare specializate la nivelul muschiului neted. Terminatiile nervoase sunt localizate in tesutul conjunctiv adiacent celulelor musculare. Terminalele nervoase contin vezicule sinaptice cu neurotransmitator. Acesta este eliberat la stimularea nervoasa in tesutul conjunctiv inconjurator de unde difuzeaza la receptorii membranei plasmatice a celulelor musculare netede din vecinatate. Celulele musculare netede sunt conectate una la cealalta prin jonctiuni cu goluri. Astfel, stimularea uneia sau mai multor celule din vecinatatea unei terminatii nervoase va fi transmisa celulelor de la distanta prin conducere electrotonica. Contractia celulei musculare poate fi reglata si de hormoni. Oxitocina si mai putin vasopresina (amandoua eliberate de lobul posterior al glandei pituitare) pot stimula contractia. Hormonii medularei suprarenale, epinefrina si norepinefrina pot fie stimula sau inhiba contractia. In unele cazuri muschiul neted poate avea si activitate contractila intrinseca in absenta inervatiei sau stimulilor hormonali. Tesutul muscular scheletic lezat poate regenera. Celulele musculare cardiace nu capacitate regenerativa. Celulele musculare netede sunt capabile de diviziune pentru a mentine sau spori numarul lor. TESUTUL NERVOS Sistemul nervos central consta in creier si maduva spinarii. Subtanta neurala ce le formeaza poate fi clasificata in substanta alba si substanta cenusie. Substanta alba este caracterizata de fibre nervoase mielinizate si de absenta corpurilor celulare neurale. Substanta cenusie se caracterizeaza prin prezenta a numeroase corpuri celulare nervoase si dendrite si absenta proceselor nervoase mielinizate. Sistemul nervos periferic consta din nervii spinali si cranieni si tesutul asociat lor. Sistemul nervos autonom consta in sistemul simpatic, parasimpatic si enteric.
17
Componentele tesutului nervos sunt neuronii si celulele de suport. Celulele gliale sunt celulele de suport ale SNC. Acstea includ oligodendrocitele, astrocitele, microglia si elulele ependimale. Celulele Schwann si celulele satelite sunt celulele de suport ale SNP. Abilitatea de a percepe si raspunde la evenimente din mediul intern si extern depinde de sistemul nervos. Neuronii reactioneaza la stimulii mecanici si chimici si efectul stimularii lor este transmis la alte parti ale aceluiasi tip de celula. In acelasi timp efectul stimularii este oferit si altor celule nervoase, musculare sau glandulare. NEURONUL Este unitatea structurala si functionala a sistemului nervos. Numarul este estimat la 9-15 bilioane. Neuronii au o mare variabilitate de dimensiune si forma. Corpul celular (pericarionul) este portiunea dilatata ce contine nucleul. Este centrul metabolic si al functiilor integrative al neuronului. De obicei este rotun sau ovoid, dar poate fi si stelat. Forma caracteristica se datoreaza numeroaselor procese ( axoni si dendrite) ce pleaca de la corpul celular. Nucleul este mare, sferic, cu nucleol proeminent, iar citoplasma contine un bogat aparat Golgi, reticul endoplasmic rugos, neurofilamente, mitocondrii si lizozomi. Corpii Nissl si aparatul Golgi indica o activitate intensa de sinteza si metabolica. Corpii Nissl sunt o aglomerare de reticul endoplasmatic rugos si poliribozomi ceea ce denota o capacitate de sinteza proteica. Se extind si in dendrite, dar nu in axon. Prelungirile celulei nervoase sunt axonii si dendritele. Prezenta acestora este singurul caracter comun al neuronilor.Neuronii se clasifica pe baza acestor prelungiri in: -unipolari cu un singur proces scurt care se divide aproape de corpul celular intr-o dendrita catre periferie si un axon catre centrul celular. -bipolari care au un axon si o dendrita distincte. -multipolari cu multe prelungiri, de obicei numeroase dendrite si doar un axon. Dendritele sunt prelungirile receptoare a caror principala functie este sa primeasca informatia de la alte celule nervoase si sa o transmita corpului celular. Dendritele au de obicei un diametru mai mare decat axonii la locul de origine din corpul celular si sunt nemielinizate. Neuronii au mai multe dendrite ce pot forma o arborizatie extinsa crescand suprafata de receptie a neuronului. Axonul este prelungirea efectoare ce preia impulsul nervos de la corpul celular si il conduce spre alt neuron sau o alta structura efectoare cum ar fi celula musculara. Axonii neurosecretori cum ar fi cei din neurohipofiza conduc hormoni de la corpul celular catre terminatia axonica apropiata de un sistem de capilare fenestrate. Axonii sunt de obicei terminatii fine, unice. Lungimea este variabila. Locul de origine al axonului nu contine substanta Nissl iar microtubulii si neurofilamentele trec prin acesta in axon. Segmentul initial al axonului este o zona scurta intre locul de origine si inceputul tecii de mielina. Acesta este locul initierii unui potential de actiune ca raspuns la suma impulsurilor primite de dendrite si corpul celular. Transportul axonal este intracelular bidirectional intre corpul celular si axon. Este o cale de comunicare intracelulara. Transportul anterograd este miscarea materialului de la corpul celular catre partile distale ale axonului. Transportul retrograd este miscarea materiallui de la partile distale ale axonului catre corpul celular.
18
Transportul lent vizeaza elemente structurale necesare pentru mentinerea si cresterea celulei. Apare doar in sens anterograd. Transportul rapid vizeaza componente membranare si neurotransmitatori asociati cu transportul de tip vezicular asociat cu microtubuli. Transportul rapid apare anterograd si retrograd. SINAPSELE sunt puncte de contact functional intre doi neuroni sau intre un neuron si o celula musculara sau glandulara. Pot fi subclasificate in sinapse chimice sau electrice. Sinapsele chimice Exista o stransa apropiere intre celule la locul interactiunii sinaptice astfel incat se formeaza o fanta sinaptica. Copmunicarea intre cei doi neuroni se realizeaza prin eliberarea unui neurotransmitator. Componenetele sinapsei sunt presinaptica, portiunea dincare se elibereaza neurotransmitatorul si portiunea postsinaptica, regiunea suprafetei celulare care este afectata de eliberarea neurotransmitatorului. Numarul sinapselor gasite pe suprafata neuronului si procesele sale variaza de la sute la zeci de mii. Numarul de sinapse este direct propoartional cu gradul de receptionare si integrare. Orice parte a neuronului poate forma sinapsa cu orice parte a altui neuron. Cele mai numeroase sinapse sunt cele axodendritice si axosomatice, dar exista si axoaxonale si dendrodendritice. Componenta presinaptica este caracterizata de prezenta veziculelor sinaptice. Vezicula sinaptica este sferica si aparent goala, dar unele contin catecolamine care dau aspect electron-opac. Veziculele sinaptice elibereaza continutul de neurotransmitator prin exocitoza in spatiul sinaptic. Exocitoza duce la suplimentarea zonei membranare a suprafetei presinaptice prin supraadugarea suprafetelor veziculelor eliberate si astfel integrate suprafetei finale. Fiecare astfel de zona mica adaugata deriva din veziculele sinaptice. Totusi, suprafata presinaptica nu creste in mod normal. Are loc o deplasare a membranei sinaptice dinspre zona de eliberare a veziculelor sinaptice spre periferia capatului presinaptic. In acelasi timp membrana este retrasa sau internalizata prin endocitoza, ulterior aceste membrane fuzioneaza cu reticulul endoplasmatic in aompartimentul presinaptic si este refolosit in formarea de noi vezicule sinaptice. Componenta postsinaptica este regiunea receptoare a sinapsei. Regiunea suprafetei neuronului de pe partea opusa a fantei sinaptice formeaza componenta postsinaptica, similara ca forma si arie cu cea presinaptica. Densitatea postsinaptica este o lama de material dens de grosime variabila atasat la partea sitoplasmatica a membranei postsinaptice. receptorii membranei postsinaptice leaga neurotransmitatorul determinand modificari de potential electric in membrana locusului postsinaptic. Domeniile citoplasmatice a moleculelor receptoare se ancoreaza in densitatea postsinaptica. Neurotransmitatorul determina depolarizarea membranei postsinaptice. Acetilcolina este un astfel de exemplu de neurotransmitator excitator. Unii neurotransmitatori produc o hiperpolarizare ca efect pe membrana postsinaptica si au efect inhibitor, cum este acidul -amino-butiric. Sinapsele electrice furnizeaza cai de minima rezistenta intre neuroni. Neurotransmitatorii chimici nu functioneaza in aceste sinapse. Sinapsele electrice sunt echivalentul jonctiunilor celulare cu gauri din alte tesuturi decat cel nervos.
19
CELULELE DE SUPORT ALE SISTEMULUI NERVOS Pentru SNC: Celulele non-neurale de suport sunt celulele gliale, mai mnumeroase decat neuronii. In practica histologica doar nucleii celulelor gliale sunt vizibile. 1. Oligodendrocitele au doar cateva prelungiri, deseori aliniate in randuri intre axonii din substanta alba. Sunt celule formatoare de mielina ale sistemului nervos central. 2. Astrocitele sunt caracterizate prin multe prelungiri care dezvolta un picior final la jonctiunea cu alte celule nervoase sau non-nervoase. Terminatiile in piciorus se afla dealungul vaselor de sange sau la suprafata creierului, maduvei spinarii si la nivelul suprafetelor ventriculare. Zonele goale ale axonilor mielinizati, la nivelul nodurilor Ranvier si sinapselor acumuleaza un invelis de astrocite. Astrocitele perisinaptice pot functiona in sensul de incredintare a neurotransmitatorului fantei sinaptice si de metabolizare a excesului de neurotransmitator. 3. Microglia indica totalitatea celor mai mici celule gliale. Prelungirile lor sunt mai mici si putin elaborate decat ale oligodendrocitelor si astrocitelor.Microglia este membra a sistemului fagocitar mononuclear si este pezenta in numar mic in SNC, dar creste la locul de contaminare sau leziune, unde este necesara functia fagocitica. 4. Celulele ependimale sunt celule epiteliale gliala care delimiteaza ventriculii creierului si canalul central al maduvei spinarii. Pentru SNP: 1. Celulele Schwnn sunt celulele formatoare de mielina ale SNP. Ele formeaza mielina in jurul axonilor mari ai nervilor periferici dar invelesc si axonii mai mici, nemielinizati ai nervilor periferici. Deoarece celulele Schwann acopera cea mai mare parte a suprafetei nervilor periferici pot regla metabolic schimburile dintre axon si spatiul perixonal. 2. Celulele satelit formeaza o lama in jurul corpurilor celulare nervoase in SNP. Ceea ce realizeaza celulele Schwann pentru axoni, realizeaza celulele satelit pentru corpurile celulare, mentinand un mediu inconjurator favorabil. Aceste celule nu formeaza mielina in jurul corpului celular. Teaca de mielina a nervilor periferici ai sistemului nervos. Insiruirea celulelor Schwann si infasurarea lor in jurul axonului formeaza mielina. Mielina izoleaza axonul de spatiul extracelular, si creste viteza de conducere a impulsului nervos. Teaca de mielina este sintetizata de celulele Schwann. In dezvoltare, celulele Schwann se dispun de-alungul axonilor si dezvolta un sant in care se potriveste fascicolul nervos.Doar un singur axon se gaseste intr-un singur sant de pe suprafata unei celule Schwann. Este rezultatul unei cresteri a numarului de celule schwann si o redistribuire corespunzatoare a acestora. Se creaza un spatiu intercelular uniform intre axon si celula Schwann si integ complexul este inconjurat de o lama externa sintetizata de celula Schwann. Apropierea stransa dintre capetele santului includ axonul si produce un mezaxon. Un singur axon poate avea mii de celule Schwann aliniate de-alungul sau. Fiecare celula Schwann elaboreaza mielina unui segment internodal al axonului. Primele lamele de mielina care se formeaza sunt apropiate dar mai tarziu devin compacte. Au fosi propuse doua mecanisme de formare a tecii de mielina.
20
1. Modelul de rotatie Celula Schwann este fizata la suprafata celulara a axonului si regiunea care contine nucleul se roteste in jurul axonului. In timpul acestui proces mezaxonul se alungeste si formeaza lamele de mielina. 2. Modelul de alungire Regiunea nucleara a celulei Schwann ramane fixata ca o limba pe suprafata neuronului si se insinueaza intre celula Schwann si axon. Cresterea continua spiralata a limbii produce lamele de mielina. Nodurile Ranvier in SNP. Un nod Ranvier este localizat la jonctiunea dintre doua segmente internodale mielinice. Zona ramasa denudata este partial acoperita de prelungiri fine de la cele doua celule Schwann adiacente. Mielina este mai putin compact aranjata in ariile paranodale. Conducerea rapida - Axonii mielinizati conduc impulsurile mai rapid decat axonii nemielinizati. Lungimea internodala a mielinei este proportionala cu diametrul axonului. Axonii mai mari conduc impulsul mai repede decat axonii mai mici si au o lungime internodala mai mare a segmentelor de mielina. Conducerea saltatorie - Pe masura ce trece printr-un nerv mielinizat, un impuls nervos sare din nod in nod. Acest tip de salt nu apare la axonii nemielinizati. Nodurile sunt singurele locuri de-alungul axonului mielinizat in care membrana axonului vine in contact cu lichidul extracelular. In aceste locuri este necesar schimbul ionic pentru propagarea potentialului de actiune. Nodurile Ranvier sunt astfel locuri de regenerare a potentialului de actiune. Teaca de mielinain sisatemul nervos central. Se formeaza prin alinierea membranelor plasmatice ale oligodendrocitelor . Portiunea centrala a oligodendrocitelor se afla distal de axonul care se mielinizeaza. Se produce un proces care seamana cu limba si care gaseste drumul spre un axon , iar apoi intr-o maniera nu prea bine inteleasa se rasuceste in jurul axonului pentru a produce lamele de mielina. De obicei un singur oligodendrocit trimite de obicei cateva astfel de procese simultan. Fiecare proces formeaza un segment de mielina internodal, iar in final fiecare segment internodal poate apartine unui alt oligodendrocit. Ca si in SNP, in SNC nodurile sunt intreruperi intre segmentele succesive di mielina. Regiunile nodale din SNC sunt mai expuse decat cele din SNP. Nodurile Ranvier din SNC au functie de regenerare a potentialului de actiune asa cum actioneaza sinodurile din SNP. DEGENERAREA SI REGENERAREA SISTEMULUI NERVOS Tesutul nervos lezat degenereaza. Regenerarea poate apare in SNP dar nu apare IN SNC. Degenerarea anterograda apare in SNP si SNC pe directia axonului, distal de locul leziunii. Axonul aflat distal de locul leziunii degenereaza din cauza intreruperii transportului axonal. Celule fagocitice derivate din celulele Schwann la nivelul SNP sau microglia din SNC indeparteaza fragmentele de mielina si de axon degenerat. Degenerarea retrograda apare proximal de locul leziunii, intre locul de leziune si corpul celular al neuronului. Cromatoliza (reactia axonului) este o serie de modificari care apar in corpul celular a unui neuron lezat. Se reduc corpii Nissl, creste volumul celular,se repozitioneaza nucleul din central, devine lateralizat.
21
Formarea cicatricei. In SNP se dezvolta o cicatrice din tesutul conjunctiv intre cele doua capete ale nervului intrerupt. Daca cantitatea de tesut cicatriceal nu este mare, capatul taiat probabil va regenera. Apropierea chirurgicala a capetelor taiate ale nervului va creste probabilitatea unei regenerari de succes. In SNC tesutul cicatriceal derivat din astrocitele proliferate impiedeca regenerarea. SANGELE Sangele este un tip de tesut specializat in care matricea extracelulara este un fluid. Sangele este continut in vasele sangvine. Sangele contine tipuri diferite de celule si elemente firmate care sunt inconjurate de un lichid bogat in proteine numit plasma. Ca si alte tipuri de tesut conjunctiv, sangele consta intr-un numar relativ mic de celule intr-un volum mare de material extracelular. ERITROCITELE sau celulele rosii ale sangelui sunt cele mai numeroase, de aprox 1000 de ori mai numeroase decat celulele albe. Eritrocitele normale au un volum mediu constant de 7-8. Microcitele sunt eritrocite mai mici decat 5, iar macrocitele mai mari decat 10. Asemenea eritrocite de volum modificat sunt prezente in anemii specifice micro sau macrocitare. Reticulocitele sunt erotrocite imature eliberate din maduva osoasa in circulatie inainte ca erotropoieza sa fie terminata. In mod normal sunt prezente in sangele periferic in procent de 1-2%. Aceste eritrocite exista ca reticulocite pentru mai putin de o zi inainte de a deveni eritrocite mature. Eritrocitele contin concentratii inalte de hemoglobina, aprox. 1/3din greutate. Rolul eritrocitelor este de a transporta gaze sangvine. Oxigenul este transportat de la tesuturi catre plamani in asociere cu hemoglobina. Este transportat si bioxidul de carbon inplasma ca ion bicarbonat. Forma eritrocitelor este importanta pentru transportul gazelor. Portiunea centrala deprimata a eritrocitelor aduce mai mmulte molecule de hemoglobina aproape de membrana plasmatica decat daca portiunea centrala ar fi sferica. Gazele au o distanta mai mica de parcurs in difuziunea intracelulara catre locul de legare pe hemoglobina in forma biconcava normala a eritrocitului. Hematocritul este procentul de volum sangvin dintr-o monstra ocupat de celulele rosii. Daca se centrigugheaza o monstra de sange se separa trei straturi: plasma supernatanta, stratul clar si stratul de depunere eritrocitara. Fractiunea eritrocitara ocupa 45% din volumul sangvin si reprezinta hematocritul. Un hematocrit mai scazut indica anemie. Stratul clar este subtire 1% si contine celule sangvine albe si plachete sau trmbocite. Aspectul normal biconcav al eritrocitelor poate fi modificat intravascular sau osmotic. Eritrocitele sunt temporar deformate de curgerea sangvina, contactul cu alte celule sangvine si de contactul cu peretele vaselor sangvine. De ex. un eritrocit poate deveni de aspect conic cu varful orientat in directia de curgere. Deformarea eritrocitelor este mai vizibila in capilarele mici cu un diametru capilar mai mic decat diametrul eritrocitului.
22
Un mediu hipoton produce o umflare a eritrocitului care devine sferoidal si palid numit fantoma. Hemoglobina se pierde din eritrocit in conditii hipotone. Procesul se numeste hemoliza. Un mediu hipertonic produce colapsul eritrocitului si conduce la un aspect crenelat prin pierderea apei. Aparenta de celula crenelata sau cu spini rezulta din plicaturarea membranei plasmatice pe scheletul citoplasmatic care ramane rigid. NEUTROFILELE sunt cele mai numeroase laucocite. Granulele citoplasmatice ale acestor celule pot fi acidofile sau bazofile indiferent de originea lor. Neutrofilele sunt mai mari decat eritrocitele. Neutrofilele imature numite celule in banda din cauza nucleului incomplet segmentat se gasesc de obicei in sangele periferic intr-un procent de 4%. Exista doua trasaturi structurale ale nucleului celulei neutrofile care le deosebesc de orice alta celula sangvina. Nucleul neutrofilului este segmentat in 3-5 lobi ceea ce ii ofera si denumirea de leucocite polimorfonucleare. Numarul lobilor nucleari nu creste cu varsta celulei. Cromatina sexuala sau corpusculul Barr apare ca o prelungire in forma de bat de chibrit si este in mod evident mai mica decat un lob nuclear. Corpusculul Barr este vizibil la 3% din neutrofile la femei, la celelalte nefiind vizibil din cauza unghiului de vizibilitate. Granulele citoplasmatic sunt de doua tipuri. Granulele azurofile, sau primare apar in toate granulocitele ca si in limfocite si monocite. Aceste grabule sunt lizozomi care contin si peroxidaza. Granulele specifice sau secundare sunt mai mici si numeroase decat cele azurofile. Au fost identificate substante bacteriostatice si bactericide. Neutrofilele sunt celule fagocitice ce functioneaza extravascular.Sunt capabile sa traverseze peretele endotelial al venelor mici si de asemenea la locul traumatismelor sau infectiilor. Bacteriile de la locul infectiei sunt fagocitate de neutrofile, iar vacuolele ce contin bacteriile se numesc fagozom. Granulele specifice fuzeaza cu fagozomul in cateva secunde dupa formare. Continutul bactericid al granulelor specifice se varsa in fagozomul unde este bocata bacteria. Granulele azurofile se unesc cu complexul fagozom-granule specifice si digera microorganismul. Prezenta unui numar mare de neutrofile in tesut este o dovada a inflamatiei acute in care continutul granulelor pot fi eliberat extracelular. Inflamatia cronica este caracterizata de prezenta macrofagelor si limfocitelor. EOZINOFILELE sunt de aprox. aceeasi dimensiune cu neutrofilele si sunt urmatoarele ca numar dupa granulocite. Le este specific nucleul bilobat si prezenta granulelor specifice acidofile. Segmentul de legatura dintre cei doi lobi nucleari poate sa nu fie prezent pe planul de sectiune si poate da interpretari eronate ca fiind binucleat. Granulele spacifice ale eozinofilelor sunt mai mari, acidofile, iar in microscopia electronica in mod specific prezinta o banda. Cantitatea eozinofilelor creste in sangele periferic in timpul infectiilor parazitice si a reactiilor alergice. Eozinofilele parasesc sangele si se acumuleaza in numar mare la locul reactiilor alergice. Eozinofilele fagociteaza complexele antigen-anticorp. BAZOFILELE au aceeasi dimensiune ca neutrofilele si sunt cel mai putin intaknite granulocite.
23
Granulele bazofile contin histamina care produce contractia muschiului neted bronhiolar si relaxarea musculaturii netede arteriolare cu extravazarea plasmei din capilare si venule. Mastocitele si bazofilele sunt similare in aparenta, ambele contin histamina dar doar mastociltele contin heparina. Mastocitele sunt celule ale tesutului conjunctiv care se concentreaza de-alungul vaselor vangvine, bazofilele sunt celule sangvine destinate spatiului vascular. Bazofilele parasesc sangele pentru a activa si in tesutul conjunctiv. Pot avea rol in hipersensibilitate si anafilaxie. Imunoglobulina E se leaga de bazofile si celulele sunt astfel sensibilizate la antigen. La expunerea secundara la acelasi antigen, se poate declansa degranularea bazofilelor cu eliberarea substantelor active farmacologic. LIMFOCITELE sunt cele mai numeroase leucocite agranulare. Limfocitele mici si medii apar in mod normal in sangele periferic, iar cele mari nu intra in mod normal in sangele periferic. Limfocitele mari sunt celule mitotice gasite in tesutul limfatic. Limfocitele au o citoplasma minima si le lipsesc granulele specifice. Limfocitele din sangele periferic pot fi clasificate in limfocite T, limfocite B si celule ucigase (natural killer). Celulele T si B nu se deosebesc histologic, celulele ucigase nu au markerii celulelor T si B pe suprafata. Ca raspuns la stimularea antigenica, celulele T si B sufera transformare blastica. Celulele T se divid si diferentiaza in celulele citolitice si in celule mediatoare ale raspunsului imun. Celulele B se divid si diferentiaza in celule plasmatice secretoare de imunoglobuline in cadrul raspunsului imun umoral. MONOCITELE sunt cele mai mari celule prezente in mod normal in sangele periferic. Servesc ca sursa de celule ce vor functiona extravascular ca macrofage. Sunt celule precursoare pentru cele mai multe celule fagocitice. PLACHETELE sau TROMBOCITELE sunt fragmente celulare anucleate. Sunt desprinse de pe suprafata megacariocitelor din maduva rosie. Plachetele au rol important in mentinerea sistemului cardiovascular. Previn pierderea de sange din vase prin coagulare. Au de asemenea un rol incomplet inteles in mentinerea integritatii stratului endotelial. Plachetele reactioneaza la suprafetele anormale ca in sangerarile vasculare. Plachetele activate adera la capetele taiate ale vaselor si mai ales la componentele tisulare ca colagenul care au fost expuse la evenimentul traumatic. Pe masura ce masa plachetelor creste, capetele deschise ale vaselor sangvine devin ocluzionate de plachete prin tromb. Granulele plachetelor isi elibereaza continutul in sistemul tubular conectat cu suprafata celulara. De aici continutul granulelor difuzeaza in plasma si spatiul ticular. Factorii eliberati de plachete participa la cascada coagularii si in formarea fibrinei care este princilalul component fibros al trombului.
24
GRUPELE SANGUINE SISTEMUL ABO Eritrocitele contin in membrana substante receptoare de natura mucopolizaharidica, denumite substante de grup, similare unor antigeni (numite aglutinogene), care sub actiunea unor factori corespunzatori lor, aflati in plasma altor persoane ( numite aglutinine) determina aglomerarea eritrocitelor in gramezi, iar procesul se numeste aglutinare. In prezenta complementului apare hemoliza. Exista mai multe sisteme de aglutinogeni. Aglutinogenii cei mai importanti apartin sistemului ABO si sunt denumiti A, B, O si sunt repartizati in asa fel incat la acelasi individ lipsesc din plasma aglutininele corespunzatoare (antiA sau antiB). In functie de repartizarea aglutininelor si aglutinogenelor, la om se intalnesc patru grupe sanguinne in sistemul ABO. Aglutinogenul A nu este o grupa unitara. Exista un aglutinogen A1 foarte activ reprezentand 80% din grupele A si AB si aglutinogenul A2 mai slab evidentiabil cu aglutinina antiA din serul persoanelor O. Caracterele de grup se transmit ereditar, conform legilor genetice, genotipurile fiind reprezentate de OO, OA, AA, OB, BB si AB, O fiind caracter recesiv, iar grupa A si B dominant. Amestecarea unor cantitati egale de sange de grupe diferite produce totdeauna aglutinare, desi practic, sangele O(donator universal) poate fi transfuzat in cantitati care sa nu depaseasca 500ml oricarui individ, iar persoanele din grupa AB pot primi pana la aceiasi cantitate de la orice donator. Faptele mentionate se explica prin aceea ca aglutininele anti-A si anti-B din sangele O al donatorului universal se dilueaza in volumul plasmatic al primitorului si de aceea nu produc hemoliza, decat daca concentratia lor depaseste limita. Hematiile insa nu-si dilueaza aglutinogenii asupra carora aglutininele primitorului concentrate in sangele acestuia, actioneaza direct. De aceea, in transfuzia de sange, de volum mic se tine seama ca aglutinogenii din sangele donatorului sa nu fie omologi aglutininelor din plasma primitorului. In transfuziile masive, pentru a preveni aglutinarea inversa, se administreaza exclusiv sange izogrup izoRh. In afara sistemului OAB exista si alti aglutinogeni slabi, care provoaca reactii de aglutinare, dar fara importanta pentru transfuzii (M, N, S, P). Existenta acestora face ca numarul grupelor sanguine la om sa depaseasca 30. Testarea acestora este importanta in genetica si in medicina legala in demonstrarea paternitatii. SISTEMUL Rh Sistemul Rh este sub raport genetic mai complex decat OAB, Antigenele sunt alcatuite din trei gene cuplate in acelasi loc.Factorul Rh este un aglutinogen, care se intalneste cu frecventa de 85% la populatia alba. Persoanele Rh negative, in proportie de 15% nu dispun in plasma de aglutinine anti-Rh. Ele pot insa apare prin izoimunizarea persoanelor Rh negative cu eritrocite Rh pozitive, ceea ce se poate realiza dupa transfuzii in 40-50% din cazuri, sau prin sarcini la mame Rh negative cu feti Rh pozitivi, dupa prima nastere cand ruperea vilozitatilor placentare elibereaza in circulatia materna hematii fetale Rh pozitive.La a doua sarcina Rh pozitiva, aglutininele materne strabat placenta si determina o intensa hemoliza a eritrocitelor fetale Rh pozitive, care se manifesta clinic prin eritroblastoza fetala. In practica medicala se efectueaza profilaxia
25
izoimunizarii Rh prin administrarea mamelor Rh negative, la 72 de ore dupa prima nastere si urmatoarele nasteri, de anticorpi antiRh care neutralizeaza eritrocitele fetale afunse in circulatia materna. Nou-nascutilor cu eritroblastoza li se practica exanguinotransfuzia cu sange Rh negativ, pentru oprirea procesului de hemoliza. Determinarea grupelor sanguine se face cu seruri test, anti-A, anti-B, la temperatura camerei si anti-Rh, cu titrul de aglutinare ridicat. Pe lama de microscop, o picatura de ser test amestecata cu sangele de analizat reprezentand 1/10 din volumul sau determina reactia de aglutinare in prezenta aglutinogenului corespunzator aglutininei din serul test. In lipsa serurilor test pentru practicarea unei transfuzii de urgenta se executa proba de compatibilitate directa, amestecand in aceleasi proportii ce mai sus ser sau plasma de la receptor cu sange integral de la donator, recoltat pe un anticoagulant, citrat de sodiu. Socul hemolitic care apare la transfuzia de sange incompatibil heterogrup este constituit din totalitatea manifestarilor clinice grave, ce apar la transfuzarea unei cantitati de sange care excede capacitatea de retinere a hemoglobinei rezultate din hemoliza in sistemul reticulo-endotelial ( constituit de spina, ganglioni limfatice, maduva osoasa) si care poate duce la moarte prin anurie in urma blocarii nefronilor cu hemoglobina. HEMOSTAZA este un proces fiziologic, prin care organismul intervine in oprirea hemoragiei produsa prin leziune vasculara (traumatica prin agenti interni sau externi, sau modificarea peretelui vascular). Prin hemostaza se previne pierderea sangelui din vasele lezate. In procesul hemostazei au rol important: vasul (factorul parietal), placutele si procesul coagularii. Hemoastaza are trei faze: 1. FAZA VASCULARA Modificarile vasculare sunt rapide si constau in spasmul musculaturii netede a vasului lezat, cu reducerea consecutiva a lumenului. Inchiderea vasului lezat se realizeaza prin mecanisme reflexe si umorale. Reflex, lezarea vasului stimuleaza terminatiile nervoase libere care conduc impulsurile nociceptive pana la coarnele laterale ale maduvei spinarii in centrii nervosi simpatici. Prin caile eferente simpatice, se produce dupa aprox.1min, constrictia vasului lezat, care dureaza cateva minute. Umoral, se produce constrictia vasului prin serotonina epiberata din trombocitele dezagregate, cu efecte de mai mare durata. In capilare, inchiderea se realizeaza prin retractia elastica a vasului lezat. In vasele mai mari, aceste mecanisme nu sunt eficiente. 2. FAZA PLACHETARA se caracterizeaza prin aderarea trombocitelor la suprafata vasului lezat, in mai multe straturi succesuve, pana la formarea unui dop, cheag trombocitar, care include si alte elemente figurate sanguine. Din placutele care se dezagrega se elibereaza factorul trombocitar care impreuna cu tromboplastina tisulara initiaza coagularea. Aderarea trombocitelor in zona leziunii este favorizata de unele substante din placute si tesuturi. 3. FAZA DE COAGULARE Incepe dupa 15-20 secunde si este totala la 1-3 minute, determinand formarea unui dop care ocluzioneaza vasul dupa 3-6 minute. Este urmata de retractia cheagului dupa 30-60minute, prin scurtarea prelungirilor emise de placute.Acesta esteun proces activ, cu consum de energie. Prin retractia cheagului se separa partea lichida-serul- de cheagul propriu-zis. Prin acest proces de retractie buzele plagii se apropie. Intr-o etapa urmatoare, din sange intra si alte elemente figurate (celule mobile), granulocite, neutrofile, limfocite. Are loc procesul de organizare fibroasa a
26
chagului. In cursul procesului de coagulare, pe filamentele de fibrina se absoarbe plasminogen care sub influenta unor activatori locali se transforma in plasmina, care produce liza fibrinei. In decurs de cateva zile apare o cicatrice fibroasa reparatorie si se reface continuitatea vasului. Procesul de hemostaza este foarte important in reactia de aparare a organismului. El devine nociv pentru peretii vaselor, apar leziuni care ii fac rugosi si umectabili (leziuni de ateroscleroza) xu depuneri de lipide in pereti. Placile ateromatoase, prin caracterul lor rugos, initiaza adeziunea si aglomerarea de trombocite in zona lezata, putand duce la formarea de trombi, cu rol obliterant. Trombii mobilizati in circulatie, reprezinta emboli. Daca ajung la plamani pe calea circulatiei venoase, declanseaza o serie de reflexe si tulburari mecanice circulatorii de ocluzie arteriala pulmonara, care dau tabloul emboliei pulmonare a carei gravitate poate duce la moarte. TESUTUL OSOS Osul este un tesut conjunctiv calcifiat. Osul are o funstie de suport si de protectie. Este de asemenea un loc de stocare a calciului si fosfatului. Ca orice tesut conjunctiv este format din celule si compartiment extracelular numit matrice. Tesutul osos formeaza oasele ca organe ale sistemului scheletal. CELULELE OSOASE sunt speceifice stadiului de dezvoltare si maturizare al osului: celule osteoprogenitoare, osteoblaste, osteocite, osteoclaste. CELULELE OSTEOPROGENITOARE sunt cel mai putin diferentiate celule formatoare de os. Ele nu prezinta depozite de matrice sintetizate. Seamana cu fibroblastele, iar celulele osteoprogenitoare de la suprafata osului vor face parte din periost, iar cele care delimiteaza cavitatea medulara vor face parte din endostiu. OSTEOBLASTELE sunt celule germinative ale osului care se dezvolta din celulele osteoprogenitoare aflate adiacent matricei osoase pe care o sintetizeaza. Osteoblastele formeaza un strat epitelialde celule cuboidale acolo unde apare depunerea osoasa si un strat delicat, incomplet de celule scuamoase pe alte suprafate osoase unde nu apare depunere osoasa. Tesutul osos este produsul de secretie calcifiat al osteoblastelor.Colagenul si componentele matricei sunt secretate de osteoblaste in spatiul extracelular. Osteoblastul secretor devine cuprins in propria secretie. Cand apare acest fenomen celula este consiterata a deveni osteocit. OSTEOCITELE sunt celule osoase mature. Un osteocit are un numar mare de procese care radiaza din regiunea centrala, continatoare a nucleului.Osteocitele sunt complet inconjurate de matricea osoasa pe care au sintetizat-o. Matricea osoasa este strabatuta de un sistem complex si extins de canalicule si lacune. Lacunele sunt cavitati in matricea osoasa care sunt ocupate de corpurile celulare ale osteocitelor. Canaliculele sunt canale fine care radiaza dinspre lacune si contin prelungirile osteocitelor. Foarte multe camalicule radiaza dintr-o singura lacuna. Vasele sanguine nu intra nici in lacune nici in canalicule. Prelungirile unui osteocit contacteaza procesele osteocitelor apropiate la nivelul canaliculilor. Datorita acestei retele se contureaza o cale celulara pentru transportul
27
nutrientilor dinspre spatiul perivascular catre celulele din lacune.Osteocitele sunt celule solitare care nu se gasesc aglomerate, asa cum sunt condrocitele in cartilagiu. Aceasta este o dovada a absentei cresterii interstitiului osului. Osteocitele pot participa la rezorbtia matricei osului printr-un proces numit osteoliza osteocitica, proces ce este mult mai putin importantin rezorbtia osului decat osteoclazia osteoclastelor. OSTEOCLASTELE sunt celulele rezorbtive osoase, adica celule fagocitice multinucleate, gigante, localizate aproape de suprafata osului, unde apare osteoclazia sau rezorbtia matricei osoase. Suprafata osteoclastului aflat cel mai aproape de suprafata osului, dezvolta o margine multiviloasa. Osteoclastul multinucleat este produs prin repetate replicari ADN intr-o singura celula. Osteoclastele sunt membre ale sistemului fagocitic mononuclear deoarece sunt derivate din monocitele singelui periferic. MATRICEA OSOASA formeaza cea mai mare parte a osului si contine componente organice si anorganice. Colagenul este principala componenta organica a osului. Aranjarea colagenului in matricea osoasa determina aspectul lamelar sau spongios. Substanta de baza este componenta nonfibrilara a matricei organice si ocupa spatiul din jurul fibrelor de colagen si cristalelor de hidroxiapatita. Substanta de baza este sintetizata de osteoblaste si osteocite. Matricea anorganica este formata de fosfat de calciu sub forma de hidroxiapatita si ajunge la un procent de 75% din masa osoasa. Mentinerea matricei osoase se face prinactivitatea osteocitelor. Moartea osteocitelor duce la rezorbtia matricei inconjuratoare. Aceasta este o dovada ca osul nu este un suprt inert. Este mai degraba un tesut dinamic in care celulele interactioneaza constant cu matricea. Osteoidul este matricea necalcifiata sintetizata de osteoblaste.Este matricea cel mai nou formata in care cristalele de hidroxiapatita nu s-au dezvoltat inca. Calcificarea apare prin cristalizarea hidroxiapatitei in asociere cu veziculele matricei care se afla in osteoid, pe cale sa inceapa calcifierea. Veziculele sunt probabil derivate din procesele osteoblastelor apropiate si osteocitelor. Veziculele acumuleaza calciu si contin enzimele necesare pentru a initia precipitarea sarurilor de calciu. Cresterea osului apare prinapozitie care este adugarea de noua matrice suprafetei osoase preexistente. Cresterea interstitiala care apare la cartilagiu nu apare la os. Tipuri de os: osul compact si spongios pot fi identificate prin inspectie vizuala a unei sectiuni prin osul decalcifiat si uscat. Osul compact este lama exterioara care defineste forma osului si include compartimentul intern de os cu spiculi. Osul spongios este format din spiculi si ocupa cea mai mare parte din interiorul oaselor lungi formate sau in curs de formare. Periostul si endostiul sunt lame de tesut conjunctiv ale osului. Periostul est lama conjunctiva de la suprafata osului si contine o lama exterioara fibroasa si o lama interioara celulara formata din celule osteoprogenitoare. Endostiul este o lama mai ptin bine definita de tesut conjunctiv si celule osteogenice aplicate peretelui cavitatii osoase. Endostiul se intinde intre tesutul osos si maduva osoasa. Spre deosebire de periost, endostiul este lipsit de o lama fibroasa distincta. Tendoanele si ligamentele se ataseaza osului. Fibrele colagene si manunchiurile de tendoane si ligamente se unesc cu tesutul conjunctiv al periostiului. Curand in
28
dezvoltarea osului, tendonul se ataseaza de suprafata osului. Incursul cresterii osului, colagenul tendoanelor devine inglobat in matricea osului.
Caile respiratorii superioare
Caile respiratorii reprezinta succesiunea de cavitati si conducte prin care areul ajunge in plamini. Dintre acestea, nasul, cavitatea nazala, faringele si laringele fac parte din caile respiratorii superioare iar traheea si branhiile principale din caile respiratorii inferioare. Nasul extern este proeminenta mediana situata in ijlocul fetei, care adaposteste o parte a cavitatii nazale (care se intinde in profunzimea maxilarului superior pana la nivelul portiunii nazale a faringelui). Limitele regiunii nazale: -superior scobitura de sub glabela. -inferior orizontala dusa imediat sub septul nazal -lateral santurile nazo-geniene. Nasul are forma unei piramide triunghiulare cu varful in sus si baza inferior careia i se descriu urmatoarele: -radacina sub glabela, intre cele doua sprincene. -dosul nasului cu doua versante laterale ce se unesc anterior intr-o margine ascutita. -varful nasului terminarea inferioara. -aripile nasului puternic arcuite, delimiteaza narile cu septul membranos intre acestea. Cavitatea nazala se afla in centrul maxilarului superior, deasupra cavitatii bucale, sub neurocraniu, inaintea portiunii nazale a faringelui si intre cele doua orbite. Cavitatea nazala comunica cu sinusurile paranazale. Se compune din doua fose nazale. Fosele nazale sunt doua tunele alungite antero-posterior, inalte si turtite lateral. Fiecare fosa nazala se imparte in doua regiuni: vestibulul nazal si fosa nazala propriuzisa. Fosa nazala propriu-zisa are o regiune respiratorie si o regiune olfactorie. Vestibulul nazal este un conduct cu directia verticala, turtit lateral, inalt de 15mm. Este limitat inferior de nara, iar superior de marginea inferioara arcuita a cartilajului lateral al nasului. Vestibulul este captusit de piele. Acesta reprezinta continuarea tegumentului. Fosele nazale Un sept median separa simetric cele doua fose. Fiecare fosa are patru pereti si comunica cu exteriorul anterior printr-un orificiu oval numit nara, iar cu portiunea nazala a faringelui posterior printr-un alt orificiu dreptunghiular numit choana. Peretele medial sau septul nazal este comun ambelor fose nazale. Are o portiune osoasa numita sept osos posterior, o portiune cartilaginoasa numita septul cartilaginos 29
anterior si septul membranos situat intre cele doua nari in completarea formei generale a nasului. Peretele lateral este alcatuit dinspre posterior spre anterior de o succesiune se oase: palatin, etmoid, cornet nazal inferior, maxila, lacrimal, nazal. Se desprind trei cornete nazale (superior, mijlociu, inferior) dispuse etajat si alungite antero-posterior. Ele prezinta un cap mai voluminos, orientat anterior si o coada posterior. Cornetele sunt fixate pe peretele lateral al fosei nazale numai prin marginea lor superioara si delimiteaza intre ele si peretii laterali ai fosei spatii numite meat bazal superior, mijlociu, inferior. Comunica cu sinusul maxilar, sinusul frontal, canalul nazolacrimal, orificiul faringian al tubei auditive. Peretele inferior sau planseul fosei nazale este format din oasele maxila si palatin. Acest perete se prezinta ca un jgheab antero-posterior de aproximativ 6cm lungime. Peretele superior sau bolta fosei nazale este ca un sant ingust antero-posterior (nazo-fronto-etmoido-sfenoidal). Bolta este neregulata ascendenta la nivelul oaselor nazale, apoi orizontala in portiunea fronto-etmoidala si descendenta sfenoidal. La nivelul frontalului bolta are raporturi cu baza sinusului frontal, iar la nivelul etmoidului tunica mucoasa a cavitatii nazale inchide orificiile lamei ciuruite a etmoidului, lasand sa treaca doar filetele nervoase olfactive si vasele. Fosa nazala este captusita de tunica mucoasa. Linia de separatie dintre regiunea superioara si inferioara a fosei nazale trece prin marginea inferioara libera a cornetului nazal superior. Regiunea inferioara sau respiratorie corespunde cornetului inferior, mijlociu si celor trei meate corespunzatoare, iar regiunea superioara sau olfactorie raspunde cornetului superior, boltii fosei si septului nazal de la acest nivel. Faringele fiind o cale comuna traseului respirator dar si digestiv, va fi studiat la aparatul digestiv. Laringele este un organ tubular cu rol respirator si fonator. Este situat in regiunea antero-mediana a gatului sub osul hioid cu care este solidar si deasupra deasupra traheei cu care se continua. Laringele este asezat inaintea portiunii laringiene a faringelui, in care proemina si inapoia lobilor glandei tiroide si muschilor subhioidieni. Mijloace de fixare: continuitatea cu faringele si traheea, unirea cu osul hioid. Dimensiuni: creste rapid pana la 3-4 ani, apoi lent pana la pubertate si din nou in ritm rapid pana la 20-25 de ani pentru ambele sexe. Caracteristica este cresterea puternica a inaltimii si diametrului sagital al laringelui la barbat. Se descrie o forma de piramida triunghiulara trunchiata, cu baza mare orientata in sus si baza mica in jos (varful), doua fete laterale, o fata posterioara, o margine anterioara si doua margini posterioare. Baza mare este orientata in sus si priveste in endoferinge. Orificiul laringelui se numeste aditus laringian. Aerul trece prin acesta in inspiratie din faringe in laringe si in expiratie din laringe in faringe. Anterior este reprezentat de relieful epiglotei. Fata anterioara a epiglotei, care priveste spre baza limbii poate fi examinata prin cavitatea bucala cand limba este apasata in jos. Se observa o foseta transversala care separa epiglota de limba, strabatuta de trei plice sagitale care leaga fata anterioara a epiglotei de baza limbii. Acestea sunt plica glossoepiglotica mediana si doua plice glossoepiglotice laterale, iar intre ele doua fosete glossoepiglotice sau valecule.
30
Posterior, aditusul este delimitat de relieful celor doua cartilaje aritenoide si cele doua cartilaje corniculate. Cartilajele corniculate proemina formand tuberculul corniculat. Intre cartilajele aritenoide aflate mai posterior se afla scobitura interaritenoidiana. Toate aceste formatiuni sunt acoperite de mucoasa. Lateral, aditusul laringian este delimitat de plicele ariepiglotice in grosimea carora se afla cartilajul cuneiform. Baza mica (varful) este marginea inferioara a cartilajului cricoid ce se continua cu traheea. Fetele anterolaterale de sus in jos sunt formate de o succesiune de lame membranoase si cartilaje: membrana tirohiodiana, lamele dreapta si stanga a cartilajului tiroid, ligamentul cricoidian, arcul cartilajului cricoid si ligamentul cricotraheal. Fata posterioara este rotunjita si proemina in portiunea laringeana a faringelui. Intre ea si fetele laterale ale faringelui se delimiteaza cate un sant laringo-faringian. Marginea anterioara rezulta din unirea celor doua fete anterolaterale la nivelul unghiului determinat de unirea celor doua lame ale cartilajului tiroid.In ea este cuprins unghiul iesind anterior al cartilajului tiroid care proemina sub piele la barbati sub forma marului lui Adam. Marginea posterioara este reprezentata de marginile posterioare ale celor doua lame ale cartilajului tiroid. Cavitatea laringelui este captusita cu mucoasa si se poate compara cu doua palnii ce se unesc prin varfurile lor. Zona varfurilor este mai ingusta datorita prezentei plicelor vocale si plicelor vestibulare care proemina in cavitatea laringelui. Palniile se largesc in sus spre aditusul laringelui si in jos spre trahee. Aceasta zona centrala ingustata, limitata in sus de plicele vestibulare si in jos de plicele vocale este glota. Palnia superioara, dintre aditusul laringelui si glota este vestibulul laringelui, iar cavitatea inferioara, dintre glota si trahee este cavitatea infraglotica.
Caile respiratorii inferioare si plamanii
Traheea este un organ tubular. Se intinde de la a 6-a vertebra cervicala de unde continua laringele pana la a 4-a vertebra toracala, unde se divide in cele doua bronhii principale. Limitele sunt relative deoarece se bucura de o mare elasticitate conferita de structura sa. Se alungeste atunci cand laringele se ridica si se scurteaza in cazul in care laringele coboara. Traheea apartine atat gatului (portiunea cervicala) cat si toracelui (portiunea toracica). Patrunde in mediastin prin orificiul superior al toracelui. Este situata in planul mediosagital al corpului inaintea esofagului. Traheea are forma unui tub cilindric cu perete posterior turtit. Ea prezinta de asemenea doua impresiuni determinate de organele vecine: 31
1. impresiunea tiroidiana situata pe partea stanga, data de lobul stang al glandei tiroide. 2. impresiunea aortica situata la terminarea organului , tot pe partea stanga, determinata de arcul aortic care se sprijina pe trahee. Portiunea cervicala are raporturi: -anterior cu istmul glandei tiroide peste care se dispun lama conjunctiva (fascia) pretraheala, muschii subhioidieni, lama superficiala a fasciei cervicale, extremitatea superioara a timusului la copil, piele.Aceasta zona este locul de electie al traheotomiei de urgenta. -posterior portiunea cervicala a esofagului. -lateral lobii glandei tiroide (hipertrofie glandei poate comprima traheea cu fenomene de dispnee), manunchiul vasculo-nervos al gatului (artera carotida, vena jugulara interna si nervul vag). Portiunea toracica este situata in mediastin, cu traiect median. Anterior este incrucisata chiar deasupra bifurcatiei de arcul aortic, din care se desprinde trunchiul arterial brahiocefalic care ulterior trece in dreapta aortei si carotida comuna stanga care trece mai sus si in stanga traheei. Mai superficial, traheea vine in raport cu vena brahiocefalica stanga, timusul la copil, stern, piele. Posterior se afla portiunea toracala a esofagului. Lateral stanga pleura mediastinala, arcul aortic, artera carotida comuna stanga. Lateral dreaptaa pleura mediastinala, vena cava superioara, arcul venei azygos. Bifurcatia traheei realizeaza raporturi cu numeroase noduri limfatice traheobronsice si : -anterior cu aorta ascendenta, -posterior cu esofagul toracic, -inferior pericardul, inima si bifurcatia trunchiului pulmonar. Traheea este formata dintr-un schelet fibrocartilaginos si captusita de mucoasa. Scheletul fibrocartilaginos este alcatuit din 15-20 de arcuri cartilaginoase elastice, in forma de potcoava incomplete in partea lor posterioara. Cartilajele sunt orizontale. Ultimul are forma literei V si proemina in interiorul traheei, la locul de bifurcare in cele doua bronhii principale. Proeminenta poarta numele de pintene traheal. Bronhiile principale dreapta si stanga rezulta din bifurcarea traheei la nivelul celei de a 4-a vertebre toracale. De la origine, bronhiile se indreapta lateral si in jos, formand intre ele un unghi de 70-75. Dupa ce ajung la hilul plamanilor, bronhiile se ramifica intrapulmonar formand o arborizatie bogata. Bronhia dreapta merge putin inapoi, iar cea stanga inainte. Bronhia dreapta se apropie mai mult de verticala, iar cea stanga se apropie mai mult de orizontala. Intrre ele se formeaza astfel un unghi ce se deschide in jos. Deoarece bronhia dreapta este mai verticala, corpii straini patrund mai usor in aceasta si mai rar icea stanga. Plamanii Organele principale ale respiratiei, plamanii sunt in numar de doi, drept si stang. Plamanii sunt continuti in cele doua seroase pleurale, complet separate intre ele. Seroasele pleurale si plamanii, separati prin mediastin sunt situati la randul lor in cavitatea toracica.
32
Cei doi plamani, suspendati prin pediculii lor pot fi comparati cu doua jumatati ale unui con taiat de la varf la baza. In acest mod, fiecare plaman prezinta o baza, un varf, doua fete si doua margini. Aceste elemente realizeaza raporturi de vecinatate prin intermediul pleurelor. Baza repauzeaza pe diafragm si se numeste fata diafragmatica. Este puternic concava, inclinata, priveste anterior si medial. Diafragma este mai ridicata in dreapta din cauza ficatului. Ca urmare, baza plamanului drept este situata pe un plan mai inalt decat baza plamanului stang. Prin intermediul diafragmului, baza plamanului stang are raporturi cu splina, fundul stomacului si partial lobul stang al ficatului. Baza plamanului drept raspunde tot prin intermediul diafragmei recesurilor subfrenice si fetei diafragmatice a ficatului. Asa se explica posibilitatea ca un abces subfrenic sa se deschida intr-o bronhie. Varful paraseste cavitatea toracica, depasind orificiul superior al acestuia cu 23cm si raspunde fosei supraclaviculare mari de la baza gatului. Varful este acoperit de cupola pleurala si raspunde prin intermediul acesteia primei coaste. Fata costala este convexa, rotunjita, priveste inainte, lateral si inapoi, urmarind curba descrisa de coaste. Coastele imprima pe aceasta fata cateva santuri transversale. Fata mediala prezinta o portiune vertebrala si o portiune mediastinala. Portiunea vertebrala patrunde in santul pulmonar al toracelui osos. Raporturile acestuia se realizeaza cu fetele laterale ale corpurilor vertebrale toracice, extremitatile posterioare ale coastelor, spatiile intercostale, nervi si vase intercostale. Portiunea mediastinala este mai larga, usor concava si anterioara. Aici este situat hilul plamanului, care reprezinta locul pe unde trec elementele pediculului pulmonar. Hilul este usor excavat si situat in treimea superioara a fetei mediale. Marginea anterioara separa fata costala de cea mediala. Este ascutita , la dreapta usor convexa, iar la stanga cu o puternica scobitura, numita scobitura cardiaca determinata de prezenta inimii si pericardului. Sub scobitura cardiaca, marginea anterioara trimite o prelungire mediala, numita lingula. Marginea anterioara raspunde inainte sternului si inapoi pericardului. Marginea inferioara circumscrie baza plamanului. Este subtire si taioasa, separa fetele costala si mediala. Baza fiind inclinata, marginea inferioara urmeaza acelasi plan, fiind mai coborata posterior. Marginea inferioara patrunde in recesul costo-diafragmatic al pleurei parietale. Pe fetele plamanilor se intilnesc fisuri sau arcuri. La plamanul stang se afla o fisura oblica care porneste de pe fata mediala a plamanului, deasupra hilului, se indreapta in sus, trece pe fata costala in jos si inainte, intersecteaza marginea inferioara si baza, apoi ajunge din nou pe fata mediala spre limita inferioara a hilului. Fisura oblica divide plamanul stang in doi lobi: superior si inferior. La plamanul drept fisura oblica are traiect asemanator. apare insa si fisura orizontala, care porneste de pe fata costala a plamanului drept, din portiunea mijlocie a fisurii oblice, se indreapta medial, strabate marginea anterioara a plamanului, ajunge pe fata mediala si se termina la nivelul hilului. Fisurile separa plamanul drept in trei lobi: superior, mijlociu si inferior. Cel mai voluminos este lobul inferior si cel mai mic este cel imjlociu. Radacina sau pediculul pulmonar este formata din totalitatea elementelor anatomice care vin sau pleaca de l aplaman. Acestea trec prin hilul plamanului.
33
Plamanul este alcatuit din bronhia principala, artera pulmonara, doua vene pulmonare, artere bronhice, vene bronhice, limfatice, plex nervos pulmonar. Raporturile contactate in interiorul pediculului, intre elementele sale componente se numesc rapoarte intrapediculare. Rapoartele extrapediculare constau in principal cu pleura. Pleurele se reflecta la nivelul ambilor pediculi pe care ii invelesc, dar numai pe portiunea lor laterala, apropiat de hilul plamanului. Portiunea mediala a pediculilor este lipsita de invelisul pleural. Vena cava superioara coboara inaintea pediculului pulmonar drept. Inapoia acestuia urca vena azygos, iar arcul acesteia depaseste in sus pediculul, se indreapta inainte, incaleca pediculul drept si se varsa in vena cava superioara. Aorta ascendenta urca inaintea traheei si pediculului stang, se continua cu arcul aortei care incaleca pediculul si apoi cu aorta descendenta, care coboara inapoia pediculului stang. Astfel, cele trei portiuni ale aortei: ascendenta, arcul si cea descendenta ocolesc dinainte inapoi pediculul pulmonar stang. Structura plamanului cuprinde: -componenta bronhiala, care se arborizeaza in ramificatii din ce in ce mai mici si conduce aerul la nivelul parenchimului pulmonar. -componenta parenchimatoasa reprezentata de totalitatea acinilor, cu rol in realizarea schimburilor respiratorii pulmonare. -stroma care delimiteaza si uneste celelalte componente ale plamanilor. -vasele si nervii plamanilor. Componenta bronhiala formeaza ramificatii intrapulmonare bogate, provenite din ramificarea bronhiilor principale. Acestea grupeaza in jurul lor celelalte componente, realizand teritoriile bronhopulmonare. Urmarind ramificatiile intrapulmonare ale bronhiilor se observa ca bronhia principala dreapta da trei bronhii lobare: superioara, mijlocie si inferioara, in timp ce bronhia principala stanga da doua bronhii lobare: superioara si inferioara. Numarul bronhiilor lobare corespunde numarului de lobi ai fiecarui plaman, bronhia lobara srevind la aeratia parenchimului lobar corespunzator. Bronhiile lobare se divid in bronhii segmentare, care deservesc teritorii bronhopulmonare mai mici numite segmente. Segmentele bronhopulmonare se caracterizeaza prin: aeratie proprie, data de bronhia segmentara, pedicul arterial propriu, stroma intersegmentara care le delimiteaza de segmentele vecine. Plamanul drept are 10 bronhii segmentare cu tot atatea segmente. Plamanul stang are 8 bronhii segmentare cu tot atatea segmente. Componenta parenchimatoasa. Bronhiolele segmentare continua sa se divida in bronhiole lobulare sau terminale, care deservesc teritorii numite lobuli. Bronhiolele lobulare se ramifica in bronhiole respiratorii ce se ramifica in ducte alveolare, terminate prin dilatatii numite saculeti alveolari. Saculatii se compartimenteaza in mai multe formatiuni veziculare numite alveole pulmonare. Bronhiola respiratorie si ramificatiile ei formeaza acinii pulmonari. Totalitatea acinilor formeaza parenchimul pulmonar unde au loc schimburile de gaze. Acinul reprezinta in consecinta unitatea morfofunctionala a plamanului.
34
Vasele si nervii plamanilor Vascularizatia plamanilor este dubla: functionala si nutritiva. Vascularizatia functionala este asigurata de trunchiul pulmonar cu cele doua ramuri ale alui: artera pulmonara dreapta si atanga si cele patru vene pulmonare: doua drepate si doua stangi. Trunchiul pulmonar porneste din ventriculul drept al inimii si duce la plamani sange neoxigenat, iar venele pulmonare aduc sange oxigenat de la plamani la atriul stang. Trunchiul pulmonar si venele pulmonare alcatuiesc vasele circulatiei mici prin care se asigura permanent schimbul de gaze. Ramurile arterelor segmentare insotesc bronhiile si strabat impreuna segmentele bronhopulmonare. Ramificatiile lor ajung la lobulii pulmonari unde se dezvolta o retea de capilare perialveolare. Venele pulmonare se formeaza din reteaua de capilare perialveolare, e unde transporta sangele bogat in oxigen. Aceste vene realizeaza legaturi cu reteaua capilara bronhica, apoi formeaza vene perilobulare, intrasegmentare, intersegmentare si din unirea tuturar afluentilor. Se formeaza in final cate doua vene pulmonare la dreapta si stanga, una superioara si inferioara ce fac parte din pediculii pulmonari. Vascularizatia nutritiva este asigurata de arterele bronhice din aorta toracica si artera toracica interna, respectiv de venele bronhice. Arterele bronhice insotesc bronhiile si se distribuie peretilor acestora, peretilor vaselor pulmonare si stromei pulmonare. Arterele bronhice ajung numai pana la nivelul bronhiolelor respiratorii unde se termina in reteaua capilara din care pornesc venele bronhice. Arterele bronhice nu asigura nutritia parenchimului respirator, a acinilor pulmonari. Nutritia parenchimului este asigurata de vasele circulatiei functionale, adica de ramurile pulmonarei. Venele bronhice se formeaza din reteaua subpleurala, din stroma pulmonara si reteaua capilara din jurul bronhiolelor intralobulare. Limfaticele se grupeaza subpleural si intrapulmonar. Cele superficiale, subpleurale merg in profunzime si conflueaza cu cele intrapulmonare. Vasele limfatice trec prin nodurile limfatice pulmonare, bronhopulmonare, traheobronsice, mediastinale. De aceea carcinomul pulmonar metastazat de-alungul acestor noduri poate da paralizia nervilor frenici. Nervii formeaza la nivelul pediculului pulmonar un plex anterior si unul posterior. Fibrele sunt parasimpatice (din nervul vag) si simpatice (din ganglionii spinali toracali). Insotesc bronhiile intrapulmonare si arterele. Fibrele parasimpatice sunt destinate musculaturii bronhiale (bronhoconstrictie) si glandelor din mucoasa bronhica (secretia). Fibrele simpatice sunt vasomotoare: regleaza calibrul vaselor, debitul pulmonar si sunt relaxante pe musculatura bronhica.
Pleura
35
Pleurele sunt doua membrane seroase care invelesc plamanii. Pleura dreapta si stanga nu comunica intre ele, fiind separate prin mediastin. Fiecare pleura este formata din doua lame sau foite: -foita viscerala fixata la suprafata plamanului. -foita parietala captuseste peretii toracelui, -intre cele doua foite se formeaza o cavitate pleurala. Cele doua foite se afla in continuitate intre ele la nivelul unei linii de reflexie. Intre cele doua foite se realizeaza o aderenta prin presiune negativa datorita unei pelicule fine de lichid numit lichid pleural. Cavitatea pleurala este o cavitate virtuala, care devine reala cand apare in ea aer sau lichide, sange, puroi. Foita viscerala acopera toate fetele, marginile, varful plamanului. Foita intra la nivelul fisurilor si tapeteaza fetele interlobare, cu exceptia doar a unei portiuni la nivelul hilului, unde se reflecta spre foita parietala. Foita parietala sau pleura parietala este unica, dar in mod didactic se imparte in: mediastinala, costala, diafragmatica si cupola. Pleura mediastinala se plaseaza in plan sagital: -deasupra pediculului pulmonar este neintrerupta, se continua cu pleura costala. -la nivelul pediculului se reflecta medial si se comporta ca un manson care inveleste pediculul dupa care se continua cu pleura viscerala. -dedesuptul pediculului se continua cu pleura viscerala. si in acelasi timp se prelungeste in jos continuand mansonul pedicular pentru a forma ligamentul pulmonar. Pleura costala se intinde de la stern anterior la corpurile vertebrelor toracalaposterior. Anterior si posterior, pleura costala se continua cu pleura mediastinala realizand recese costomediastinale. In jos, pleura costala formeaza cu cea diafragmatica un reces costodiafragmatic. Deasupra varfului plamanilor, pleura costala se continua cu pleura mediastinala, realizand la acest nivel cupola pleurala. Pleura diafragmatica acopera fata superioara a diafragmului, de care este fixata prin intermediul unui tesut conjunctiv. Cupola pleurala sau domul pleural este un reces costomediastinal superior, ce acopera varful plamanului. Spatiul complementar face parte din cavitatea pleurala. Este acea portiune a recesurilor in care doua portiuni ale pleurei parietale se aplica una pe cealalta. Cel mai mare spatiu complementar este la nivelul recesului costodiafragmatic Pleura costala se aplica pe o distanta variabila pe pleura diafragmatica. In inspir, plamanul patrunde in recesul costodiafragmatic, fara sa-l ocupe cu totul. Portiunea neocupata de plamani reprezinta spatiul complementar. El este virtual ca si restul cavitatii pleurale.
Fiziologia respiratiei
Ventilatia pulmonara 36
Mecanica ventilatiei Inspiratia este un proces activ care se datoreaza contractieiuschilor inspiratori, avand ca urmare marirea tuturor diametrelor cutiei toracice. Diametrul longitudinal creste datorita contractiei muschiului diafragm. Cand diafragmul coboara si se aplatizeaza, va presa asupra viscerelor abdominale, le va cobora, exercitand prin intermediul lor o presiune laterala asupra rebordurilor ultimelor coaste.Astfel se mareste si circumferinta bazei diafragmului.Dupa sectionarea (experimentala) sau paralizia nervilor frenici, desi muschii cutiei toracice compenseaza miscarea in inspir, diafragmul va fi aspirat de vidul toracic si se va bolti mai mult. Aceasta leziune va fi letala. Expiratia apare dupa inspiratie (1secunda) fara pauza si la dult dureaza 2 secunde. Expirul este pasiv, energia consumata de fibra musculara in inspir se reface, si cutia toracica revine la forma initiala. In efort fizic sau obstacole pe caile aeriene, expiratia poate deveni activa prin interventia muschilor expiratori accesori. Acestia comprima viscerele abdominale, deplaseaza in sus diafragmul si apropie rebordurile costale. Vidul pleural este rezultatul aspiratiei pe care plamanul elastic o exercita in zona spatiului pleural. In inspir, plamanii sunt aspirati de vidul pleural in crestere si urmaresc cu fidelitate miscarile cutiei toracice. Coborarea diafragmului deschide recesurile costodiafragmatice unde patrund plamanii. Deschizand bilateral pleurele, plamanii se retracta brusc spre hil, in lipsa vidului intrapleural si poate apare moartea prin asfixie. In parenchimul pulmonar se descriu anatomo-functional: -o zona hilara, rigida fara elasticitate in care se afla ramificatiile bronhice, vase si in care expansiunea este absenta. -o zona perihilara in care plamanul are expansiuni reduse. -o zona mobila activa a plamanului, periferica care executa excursiile cele mai mari si ventilatia activa. Deplasarea aerului in arborele respirator. Viteza aerului in caile reapiratorii superioare este mai mare si pe masura ce arborele bronsic se ramifica, iar suprafata totala de sectiune a ramificatiilor creste, viteza s e reduce proportional catre nivelul alveolelor, unde miscarea este o amestecare a aerului proaspat cu cel anterior continut. In zona de expansiune maxima se produce murmurul vezicular care s-ar datora patrunderii in mod brusc a aerului inspirat, din ductele alveolare in sacii alveolari si propagarii suflului tubar de-alungul coloanei de aer. Volume si capacitati respiratorii Volumul respirator curent este cantitatea de aer ce se schimba in respiratia normala. Volumul expirator de rezerva este cantitatea de aer ce poate fi expirata fortat la sfarsitul unui expir normal. Volumul inspirator de rezerva este volumul ce mai poate fi inspirat la sfarsitul unui inspir normal. Volumul rezidual este aerul ramas inca in plamani la sfarsitul unui expir maxim. Capacitatile respiratorii se numesc volumul de aer ce rezulta din sumarea mai multor volume mentionate. Capacitatea pulmonara totala reprezinta toate volumele de aer mentionate fiind cantitatea de aer ce se afla in plaman la sfarsitul unui inspir fortat.
37
Capacitatea vitala reprezinta volumele expirate fortat dupa un inspir ,axi, (volumul inspirator de rexerva + volumul respirator curent + volumul expirator de rezerva). Capacitatea inspiratorie reprezinta volumele inspirate maximal la sfarsitul unui expir normal. Capacitatea functionala reziduala reprezinta suma volumului rezidual si a volumului expirator de rezerva. Capacitatea vitala reprezinta volumele ce pot fi utilizate in veetilatie si a fost propusa ca masura a functiei respiratorii. In cursul efortului fizic in care se intensifica schimburile respioratorii, adaptarea ventilatiei pulmonare, cu cresterea debitului respirator se face nu atat prin marirea amplitudinii respiratiilor (volumelor inspirator si expirator re rezerva) cat mai ales prin cresterea frecventei respiratorii. De aceea debitul respirator ofera indicii mai fidele asupra functiei ventilatorii. Debitul respirator de repaus este de 8l/min (0,5l aer 16 respiratii /min = 70 respiratii/min). Debitul respirator maxim este de 180 200l/min. Ventilatia alveolara Aerul inspirat este distribuit numarului mare de spatii alveolare intr-un mod aproape uniform. Cantitatea de aer inspirat repartizate fiecarei alveole depinde de gradul de expansiune ale acestora. Fiecare spatiu alveolar primeste o cantitate de aer inspirat proportionala cu volumul lui. Expansiunea inegala a spatiilor alveolare depinde la subiectul sanatos in mod predominant de gradientul vertical de presiune pleurala (datorita gravitatiei, spatiile apicale inpozitie ortostatica sunt mai expansionate decat cele bazale). Distributia sangelui venos trimis de ventrivulul drept in capilarele pulmonare depinde de gravitatie. Zonele bazale ale plamanului sunt mai irigate. Raportul ventilatie-perfuzie nu este identic in teritoriile pulmoare nici macar la subiectul normal, la care se observa o neuniformitate in repartitia pe unitati a aerului si sangelui, dar mai ales in conditii patologice.
Schimburile gazoase
La nivelul membranei alveolare, la trecerea din aerul alveolar in sangele din capilarele pulmonare, unde se combina cu hemul din molecula de hemoglobina, iar bioxidul de carbon trece din sange in aerul alveola, de unde este eliminat la exterior cu expiratia. Transferul gazos prin membrana alveolocapilara este determinat de diferenta dintre presiunile partiale pe care la exercita moleculele de gaz de o parte si de alta a membranei. Presiunea partiala a oxigenului este mai mare in aerul alveolar decat in sangele venos amestecat, care patrunde in capilare, si astfel oxigenul trece din aer in sange pana cand presiunea lui partiala venoasa urca la nivelul alveolar. Persiune apartiala a bioxidului de carbon fiind mai mare in sangele venos amestecat, dacat in alveole, transferul se face invers, pana cand presiunea bioxidului de carbon in sange caboara la nivelul alveolar. Volumul de oxigen sau bioxid de carbon ce strabate membrana alveolocapilara in unitatea de timp, depinde de gradientul (diferenta) de presiune partiala a gazului de o 38
parte si de alta a membranei, de suprafata membranei disponibile pentru transfer, in anumite conditii de grosimea ei si de raporturile ventilatie-perfuzie. Transportul de oxigen Oxigenul transferat prin membrana alveolocapilara este transportat de sangele care paraseste plamanii, sub doua forme: -dixolvat inplasma, -si mai ales legat chimic de hemoglobina. Saturatia hemoblobinei cu oxigen (>95% la subiectii normali) depinde de presiunea partiala a oxigenului dizolvat in plasma, ph-ul sangvin, temperatura corpului. Scaderea presiunii partiale a oxigenului la jumatate din valoarea normala (100mm Hg) diminua cu numai 5% saturatia in oxigen.. La presiuni partiale mici, eliberarea oxigenului se face rapid si saturatia oxigenului scade cu 30% cand presiunea partiala a oxigenului ceboara de la 30 la 15 mm Hg. Aceasta favorizeaza fixarea oxigenului in sange la nivelul plamanilor si eliberarea lui in tesuturi. Aceasta este decrierea curbei de disociere a oxigenului. Transportul bioxidului de carbon Cea mai mare parte a bioxidului de carbon transportat de sange este sub forma de combinatie chimica: -bicarbonat -carbhemoglobina. si o mica parte este dizolvat in plasma. Fractiunea dizolvata este aceea care determina (prinpresiunea partiala) schimbul de bioxid de carbon. Eliberarea de bioxid de carbon la nivelul plamanilor si fixarea acestuia la nivelul tesuturilor se desfasoara potrivit unei curbe de disociere a bioxidului de carbon influentata atat de presiunea partiala a bioxidului de carbon cat si de saturatia oxigen. Curba de disociere a bioxidului de carbon este abrupta, ca urmare la variatii mici ale presiunii partiale de bioxid de carbon, se elibereaza volume mari de gaz.
Reglarea respiratiei
Sub denumirea de centru respirator este desemnata formatiunea neuronala bulbara care reprezinta structura nervoasa indispensabila reglarii respiratiei, dotata cu automatism si cu proprietatea de a raspunde la stimuli umorali si reflexi. Centrul respirator bulbar cuprinde doi centri situati simetric, de o parte si de alta a liniei mediane: -centrul inspirator in 1/3 inferioara a bulbului. -centrul expirator cu o pozitie mai ventrala si mediala. Delimitarea celor doi nu este anatomica, ci functionala in raport cu efectele stimlarii electrice. Exista fibre intercentrale homolaterale si incrucisate ce ii pun pe cei doi in legatura. Influentele intercentrale apar frecvent in activitatea centrilor respiratori: -in reflexul de voma se initiaza o faza de pegatire in care miscarile respiratorii sunt frecvente si ample, si apoi faza de expulzie in care are loc oprirea respiratiei la sfarsitul inspiratiei. -in cursul deglutitiei respiratia se opreste. -activitatea respiratiei influenteaza activitatea centului cardioinhibitar, determinand aritmia respiratorie.
39
Automatismul respirator este influentat mai ales de proprietatile chimice ale sangelui. Stimularea umorala a centrului respirator Rolul principal in reglarea respiratiei revine concentratiei de bioxid de carbon, din acest motiv a fost numit si hormon respirator. Ventilatia maxima sub influenta bioxidului de corbon se desfasoara cu o senzatie subiectiva de respiratie dificila numita dispnee. Injectarea unui acid in sangele care iriga bulbul produce activarea respiratiei. S-a considerat ca eliberarea ionilor de hidrogen prin disociere ar fi stimulatoare. Dar in realitate, acizii nevolatili ataca bicarbonatul de sodiu sangvin si elibereaza bioxidul de carbon responsabil de stimularea respiratiei. Daca prin injectarea clorurii de amoniu se produce acidoza (ph-ul scade< 7,35 7,20) stimularea respiratiei se manifesta printr-o crestere a debitului respirator. Acidoza respiratorie poate produce stimulare respiratorie cu toata existenta unei hipocapnii. Hipoxia axtioneaza putin marcat pentru stimularea centrilor respiratori. Efectele se realizeaza prin zona chemoreceptiva sinocarotidiana. Ionii de calciu modereaza excitabilitatea centrilor respiratori. Cand ca sangvin scade determina cresterea sensibilitatea centrilor respiratori pana la oprirea espiratiri.Administrarea iv. al Ca-ului normalizeaza functia centrilor respiratori. Temperatura actioneaza stimulator asupra centrilor respiratori.
Stimularea reflexa a centrilor respiratori Excitarea reflexa a centrilor respiratori se face prin zonele preso si chemo sensibile. Zona reflexogena alveolara. Este bogata in receptori care sunt stimulati de gradul de distensie al peretelui alveolar ce determina reflex expirul. Relaxarea peretilor alveolari in expiratie, deaclanseaza pe calea aferentelor vagale inspiratia. Cele mai frecvente descarcari bioelectrice vagale au loc in inspr. Sectionarea vagilor rareste respiratia. Zonele reflexogene sinocarotidiana si aortica. Sunt alcatuite din formatiuni glomerulare perechi. Corpusculii carotidieni si aortic sunt foarte bogat irigati de capilare sinusoide, cu debit mare. Frecventa impulsurilor creste cu scaderea presiunii partiale a oxigenului in sangele arterial (hipoxemia). Scaderea ph-ului si cresterea bioxidului de carbon au efecte mai putin marcate asupra zonei chemoreceptive. Stimularea presoreceptorilor sinocarotidieni si aortici prin cresterea presiunii arteriale, determina la cresteri moderate ale presiunii, rarirea , iar la valori mari, oprirea respiratiei. Scaderea presiunii produce polipnee prin mecanism reflex. Alte zona reflexogene. Stimularea mucoasei cailor respiratorii subglotice determina reflex accelerarea respiratiei, iar stimularea mucoasei supraglotice inhibitia. Conluzie: in repaus la presiunea atmosferica obisnuita, activitatea ritmica a centrului respirator este mentinuta prin stimulare directa a acestuia de catre presiunea partiala a bioxidului de carbon sangvin, concentratia ionilor de hidrogen, la care se adauga stimulul presiunii partiale a oxigenului arterial in zonele chemoreceptoare.
40
In urma hiperventilatiei voluntare se produce o hiperoxigenare a sangelui, cu disparitia stimulilor presiunii partiale a oxigenului arterila, alcaloza in urma hipocapniei, datorita scaderii presiunii partiale a bioxidului de carbon alveolar. Efectul este de scadere a ritmului respirator.
Aparatul circulator Inima
Inima este cuprinsa in sacul fibroseros format de pericard si este situata in mediastinul anterior. Axul longitudinal al cordului aflat intre vaza si varf este orientat oblic ianinte, la stanga si in jos. Baza cordului priveste opus, la dreapta, in sus si posterior. Varful inimii corespunde spatiului V intercostal stang, putin medial de linia medioclaviculara, Inima are forma unui con turtit, cu varful globulos, baza si doua fete , doua margini. Fata sternocostala sau anterioara corespunde in cele 2/3 inferioarea ventriculilor, iar in 1/3 rupeioara atriilor si vaselor mari. Ventriculii sunt despartiti prin santul interventricular anterior, ce se continua putin la dreapta de varful real al inimii, prin incizura apicala a cordului. Santul atrio-ventricular anterior este mascat de emergenta vaselor mari din ventricul, trunchiul pulmonar din ventriculul drept si aorta din ventriculul stang. Portiunile laterale ale acestui sant sunt mascate de auriculele drept si stang. Fata diafragmatica sau inferioara este formata in cea mai mare parte de ventriculul stang si mai putin de ventriculul drept, iar posterior de aria atriala, unde in dreapta se varsa vena cava inferioara. Intre cei doi ventriculi se afla santul interventricular posterior, mai aproape de marginea ventriculului drept, iar intre atrii si ventriculi santul atrioventricular posterior in care se afla sinusul coronar. Marginea dreapta sau ascutita este cea care se poate observa mai bine. Apartine ventriculului drept si priveste inferior. Marginea stanga sau fata pulmonara este cea mai etalata si priveste spre fata mediala a plamanului stang la nivelul inciziei cardiace. Baza inimii este formata din atrii si vasele mari. Cea mai mare parte a bazei apartine atriului stang, atriul drept fiind mai lateral. In atriul drept se deschid venele cave inferioara si superioara, iar in atriul stang cele patru vene pulmonare, cu trunchi scurt si directie transversala. Varful inimii este mobil, se proiecteaza in spatiul V intercostal stang linia medioclaviculara si este reprezentat de varful ventriculului stang. Cavitatile inimii Atriul drept este cavitatea de receptie a sangelui venos din marea circulatie. Pe peretele superior se deschide vena cava superioara, iar pe cel inferior vena cava inferioara si mai medial colectorul venos coronar numit sinus coronar. De pe peretele anterior se desprinde o prelungire ce traverseaza marginea dreapta a inimii catre santul atrioventricular anterior numita auricul drept.
41
Orificiul atrio-ventricular drept este prevazut cu valva atrio-ventriculara dreapta sau tricuspida. Se afla pe peretele anterior al atriului drept si la baza ventriculului drept. Cele trei valvule cuspide sunt anterioara, posterioara si septala. Prin baza lor se prin de un inel fibros, iar prin cordajele tendinoase de muschii papilari so de peretii ventriculului drept. Fiecare cuspida prezinta o fata axiala neteda si una parietala spre peretele ventriculr pe care se insera cordajele tendinoase. Datorita cordajelor, in sistola valvulele se inchid fara a se putea da peste cap catre cavitatea atriala. Ventriculul drept are peretele muscular mai gros decat atriul drept si prezinta pe suprafata interna reliefuri ridicate sub forma de muschi papilari, cate unul pentru fiecare perete. de la care pleaca cordaje tendinoase spre fiecare din cele trei cuspide valvulare atrio-ventriculare. Muschii papilari, la fel ca peretii ventriculului drept si cuspidele, au aceeasi denumire: anterior, posterior, septal. Alaturi de orificiul atrio-ventricular drept, la nivelul bazei ventriculului, dar mai medial se alfa orificiul trunchiuui pulmonar. In sistola, ventriculul drept se inchide orificiul atrio-ventricular drept si se deschide portiunea ventriculara dinaintea trunchiului pulmonar numita conul pulmonar si apoi valvulele semilunare ale acestuia, sangele fiind expulzat in trunchiul pulmonar si astfel in mica circulatie. Atriul stang are peretele mai subtire decat atriul drept. In el se deschid cele patru vene pulmonare, doua drepte si doua stangi care aduc sangele oxigenat din plamani. Auriculul stang este o prelungire anterioare care inconjoara spre stanga originea trunchiului pulmonar in partea stanga, mascand santul atrio-ventricular anterior in jumatatea stanga. Orificiul atrio-ventricular stang este prevazut cu valva atrio-ventriculara stanga, mitrala sau bicuspida, formata dintr-o cuspida anterioara si posterioara. Orificiul se afla pe peretele anteior al atriului stang si la baza ventriculului stang. Cordajele tendinoase spre cele doua valvule sunt mai puternice, iar muschii papilari anterior si posterior mult mai bine reprezentati. Prin baza lor, cele doua cuspide se prind pe inelul fibros al atriului stang. Ventriculul stang primeste sangele arterial evacuat din atriul stang. Dupa inchiderea valvei atrio-ventriculare stangi in sistola, prin contractia muschilor peretelui ventriculului stang, care este de trei ori mai gros decat al ventriculului drept, sangele este expulzat prin orificiul aortic in marea circulatie. Se deschide astfel portiunea mediala a ventriculului stang numita con arteril si valvele semilunare aortice. De remarcat este ca musculatura ventriculului stang este mult mai groasa, capabila sa lucreze sub un regim presional mult mai mare, pentru a putea sustine miscarea coloanei sangvine prin marea circulatie. Miocardul este tapetat la exterior de pericard, alcatuit din foita viscerala, ce adera de miocard si de foita parietala ce realizeaza sacul sau cavitatea pericardica, care contine lichid pericardic in cantitate mica. Pe fata interna a miocardului, inspre cavitatea cardiaca, se afla stratul endocardic, care se continua prinorificiile vasculare cu endoteliul vaselor, ce asigura netezimea suprafetelor ce vin in contact cu sangele circulant. Miocardul atrial si cel ventricular sunt separate, fiind unite numai prin fascicolul atrioventricular de tesut excitoconductor.
42
Tesutul excitoconductor al inimii este format din celule care conduc excitatia sau stimulul miocardic primitiv in mod specific prin miocard la toate cavitatile, si apoi in mod difuz intercelular, la toate celulele musculare miocardice. Tesutul excitoconductor este reprezentat de nodul sinoatrial, aflat in atriul drept la loculde intrare a venei cave superioare, apoi nodul atrio-ventricular aflat in grosimea septului interatrial, fascicolul His in portiunea superioara a septului interventricular, ramurile dreapta si stanga ale fascicolului His care se distribuie catre varful fiecarei cavitati ventriculare si fibrele Purkinje ce se ramifica in grosimea miocardului. Functional se descriu cai specifice internodale care ar completa continuitatea intregului sistem excitoconductor, dar individualitatea anatomica a acestora nu este admisa. Functia tesutului excitoconductor asigura activitatea ritmica a inimii si prin aceasta adaptarea la conditiile hemodinamice diverse.
Proprietatile miocardului
Excitabilitatea este o functie a membranei fibrelor musculare miocardice, conditionata de polarizarea electrica a acesteia, prin repartitia inegala a ionilor intracelular (unde predomina K si Mg) si extracelular (unde predomina Ca si Na). Potentialul de repaus se masoara su microelectrozi intracelulari, pe fibra musculara izolata si este aprox. -90mV. Sub actiunea stimulilor (electric, mecanic, acid, baze diluate) care depasesc pragul de excitabilitate, se produce o depolarizare a membranei si se genereaza potentialul de actiune. Miocardul reprezinta un sincitiu (celule aflate in legatura una cu cealaltaprintr-o dispozitie de retea) in care excitatia sub forma undei de depolarizare se propaga in totalitate generand contractia intregului muschi miocardic. Unda p precede sistola arteriala mecanica, iar complexul qrs apare inaintea deschiderii valvelor sigmoide arteriale. Stimularea electrica a miocardului in perioada sistolica nu este urmata de contractie, doar daca aceasta are loc spre sfarsitul sistolei si atunci apare o contractie supranumerara numita extrasistola. Inecitabilitatea periodica a miocardului este datorata depolarizarii si repolarizarii membranelor fibrelor miocardice in cursul excitatiei. Automatismul Inima, mentinuta in caonditii fiziologice, in afara organismului isi continua activitatea printr-o functionare spontan, repetitiva, cu caracter ritmic, denumita automatism. suportul morfologic al automatismului este sistemul excitoconductor al inimii format din tesut embrionar specific. Acesta este reprezentat de fibre miocardice ce pastreaza caractere embrionare; diametru mai mare, lungime redusa, nuclei voluminosi, sarace in fibre contractile. Nodul sinoatrial se afla in peretele atrial drept, la locul de varsare al venei cave superioare, ca o formatiune subendocardica de 1-2cm lungime. Nodulatrioventricular este situat posterior, in peretele interatrial, deasupra septului atrioventricular, in apropierea locului de insertie al calvulei tricuspide. 43
Fasciculul His este o continuare a nodului atrioventricular spre ventricule si reprezinta unica structura de comunicare miocardica intre atrii si ventricule. Fasciculul His incepe cu o ramura comuna, care apoi se bifurca intr-o ramura dreapta, mai scurta si una stanga care strabate septul. Ramurile ajung de-a lungul septului la varful ventrivulilor, de unde se rasfrang si se ramifica in evantai in peretii miocardici sub forma retelei Purkinje. Caracteristic pentru activitatea electrica a celulelor nodale sinusale, este depolarizarea lenta a membranei in timpul diastolei, cae ajunsa la un prag, genereaza automat potentialul de actiune. Ritmicitatea este frecventa de descarcare a nodului sinoatrial de 70-80/min. La iesirea sa din functiune, automatismul este preluat de nodul atrioventricular cu o frecventa de 40-60/min si apoi de fasciculul His cu o frecventa de 25-35/min. Conductibilitatea este excitatia care ia nastere in celulele cele mai active, sinoatriale, si apoi prin functia de conducere a impulsurilor se raspandeste in mod relativ lent in musculatura atriala, apoi la nodul atrioventricular, fascicolulHis, ramurile sale, reteaua Purkinje si in mod difuz in intregul miocard ventricular. Ventriculul drept este activat cu putin inaintea celui drept, datorita ramurii drepte a fascicolului His care este mai scurta. Contractilitatea. Unda de depolarizare genereaza o unda de contractie in miocard. Aparatul contractil este complex si consta din filamente de actina si miozina, situate paralele in structura miofibrilelor. In contractie, ionii Ca eliberati, leaga cele doua filamente de actina si miozina prin combinarea ATP-ului si astfel se scurteaza puntea de ATP ce produce scurtarea muschilor in contractie. Legea totul sau numic a fost formulata privitor la activitatea inimii, fata de stimularea electrica. Peste valoarea de prag ce determina o depolarizare, se produce contractie maxima. Amplitudinea contractiei ar fi independente de intensitatea stimulului. Aceasta lege ste valabila pentru generalizarea procesului de excitatie in miocard si nu pentru forta de contractie a miocardului. Tonicitatea este starea de semicontractie ce se pastreaza si in diastola. Persista dupa denervare la inima in vitro, fiind o proprietate intrinseca a miocardului.
Revolutia cardiaca
Fazele activitatii inimii constau in contractii (sistole) prin care se realizeaza evacuarea cavitatilor si relaxari (diastole) in cursul carora are loc umplerea cavitatilor. Rezultatea este deplasarea sangelui intr-o singura directie in inima, datorita rolului de supape ale valvelor. Inima functioneaza ca o pompa, expulzand intermitent cu fiecare contractie (sistola) in sistemul arterial o cantitate de sange (debit sistolic). Arterele mari, datorita elasticitatii lor, transforma curgerea discontinua imprimata sangelui de inima, intr-o curgere continua. Ciclul cardiac la om incepe cu sistola atriala, sub forma unei unde de contractie ce invadeaza atriile, pornind din zona de varsare a venelor cave, unde se gaseste nodulul sinoatrial spre orificiile atrioventriculare. In cursul sistolei atriale o buna parte din sangele atrial este expulzat in ventriculi, o mica parte regurgiteaza in vene, acestea neavand un sistem valvular de inchidere ermetica. 44
Dupa sistola atriala urmeaza sistola ventriculara, in cursu careia sangele este expulzat in arterele aorta si pulmonara. Ventriculele se contracta mai brusc, dezvolta o presiune mai mare, iar volumul ventriculului se reduce, varful inimii apropiindu-se de baza. Exista o perioada in care diastolele atriala si ventriculara coincid, intre sfarsitul sistolei ventriculare si inceputul sistolei atriale. Frevcenta cardiaca. Ritmul cardiac. Valorile normale la adult sunt 60-80/min. Frecventa contractiilor cardiace pe minut variaza cu specia, in functie de talia animalului. Animalele cu talie mare, la care procesele metabolice se desfasoara mai lent au o frecventa mai redusa. Frecventa variaza si cu varsta: de la 140/min la nastere, la un an se reduce la 120110/min si la adult la 70/min. Accelerarea ritmului cardiac se face mai ales pe seama diastolei, sistola scurtandu-se foarte putin. Zgomotele inimii Sunt produse de modificarea vitezei sangelui prin curgerea sa turbulenta si vibratiile consecutive ale valvelor. Zgomotele cardiace se asculta prin aplicarea directa a urechii pe peretele toracic sau cu stetoscopul cu membrana. Clasic, se cunosc doua zgomote audibile. Intre cele doua zgomote exista o pauza, care dureaza cat sistola ventriculara, iar intre zgomotele II si I exista o pauza mare, cu durata diastolei ventriculare. Zgomotul I se percepe in spatiul V intercostal stang pe linia medioclaviculara. Locul coincide cu socul apexian. Ca intensitate, acest zgomot este mai puternic decat zgomotul II. La formarea acestui zgomot contribuie: -deplasarea sangelui dinspre peretele miocardului ventrivular care se contractam inspre valvulele atrioventriculare care erau deschise la sfarsitul diastolei. Aceasta determina oprirea fluxului de sange care din atriu curgea spre ventricul, ce determina si ea zgomotul I. -deschiderea valvelor sigmoide ale atrerelor mari si vibratiei peretilor aortei. Zgomotul II urmeaza zgomotului I dupa pauza mica. Este sincron cu inchiderea valvelor sigmoide ale aortei si arterei pulmonare si se asculta la locul de proiectie al acestor valve pe peretele toracic. Zgomotul este produs de vibratii cu frecventa mai ridicata si are o tonalitate mai ridicata. Pauza mica si pauza mare. Intervalul dintre zgomotul I (la inceputul sistolei ventriculare) si zgomotul II (la sfarsitul sistolei) este pauza mica, si functional constituie sistola hemodinamic activa, pe EKG fiind intervalul p-qrs-t. Intre zgomotul II si I este prezenta pauza mare, de relaxare hemodinamica generala. Zgomotul III si IV nu sunt perceptibile fonic, decat sub forma de inregistrare sub amplificare. Zgomotul III este datorat trecerii sangelui din atrii in ventricule in faza de umplere rapida la inceputul diastolei. Zgomotul IV este prezent in hipertrofia atriala, sincron cu sistola atriala.
45
Socul apexian se palpeaza in spatiul V intercostal stang pe linia medioclaviculara, unde se gaseste varful nimii in contact cu peretele toracic. In sistola, inima sufera o torsiune, prin care ventriculul stang care este usor posterior, devine mai anterior si izbeste peretele toracic in spatiul intercostal, determinand o usoara bombare a acestuia. Debitul cardiac sau circulator este expresia finala a activitatii inimii. Se exprima prin cantitatea de sange expulzat intr-un inut de inima dreapta sau inima stanga, aceste cantitati fiind egale. Cantitatea de sange expulzat cu fiecare sistola reprezinta debitul sistolic. La om in repaus este de 4-6l/min. In efort excesiv poate ajunge la 30-35l. Pulsul arterial Dilatarea aortei la originea sa, cu cresterea presiunii datorita sangelui expulzat de ventriculi, se ransmite sub forma unei vibratii care se propaga de-alungul peretilor vasculari in intreg teritoriul arterial. Este unda pulsului. Ea nu este identica cu viteza de circulatie a sangelui. Fiecare revolutie cardiaca genereaza o unda de presiune care se propaga de-alungul aortei si ramurilor sale si se numeste puls. Unda pulsatila care se palpeaza prin compresiunea arterei pe o suprafata rigida, consta in destinderea peretilor arteriali cu cresterea presiunii si diametrului arterei. Presiunea arteriala In artere sangele circula sub presiunea intretinuta de contractiile ritmice ale inimii, de elasticitatea peretilor arteriali si de rezistenta periferica a arborelui arterial. Presiunea are la inceputul undei pulsatile diastolice valoarea de 80mmHg, iar in sistola atinge un varf maxim de 120mmHg. Diferenta aproximativa dintre cele doua valori constituie amplitudinea presiunii. Pentru masurarea presiunii arteriale se pot utiliza; -metoda palpatorie. -metoda ascultatorie. -metoda oscilometrica. Metoda palpatorie. In timp ce se palpeaza artera radiala, se introduce aer in tensiometru , cu para de cauciuc, pana pulsul devine imperceptibil, artera fiind colabata prin presiunea superioara celei din interiorul arterei. Se decomprima aparatul, la aparitia primei pulsatii, urmarind nivelul manometrului. Prima pulsatie indica o presiune in manseta putin inferioara presiunii sistolice din artera. Singura ce poate fi detectata prin aceasta metoda este presiunea sistolica. Metoda ascultatorie se bazeaza pe principiul Korotkov, dupa care asupra arterelor comprimate se aud zgomote si sufluri proportionale cu gradul compresiunii si debitului arterial. Pe suprafata plicii cotului, in dreptul santului bicipital medial, unde artera brahiala este superficiala se fixeaza stetoscopul cu membrana. Se creste presiunea in manseta pana la disparitia pulsului. La descompunerea progresiva apar fenomene ascultatorii: -zgomot discret la scaderea presiunii din manseta cu putin sub presiunea sistolica, cand artera se deschide si presiunea atinge valoarea sistolica. -cu fiecare pulsatie in stetoscop apar zgomote care cresc in intensitate. -zgomotele se transforma in sufluri mai puternice. -zgomotele devin brusc surde sau dispar presinea diastolica.
46
Primul zgomot la decomprimare este valoarea sistolica, iar ultimul zgomot, cea diastolica, Exactitatea procedeului este foarte mare. Metoda oscilometrica. Amplitudinea oscilatiilor peretilor arteriali este maxima cand presiunea pneumatica exercitata asupra acestor pereti este egala cu presiunea medie a sangelui din artera. Practic, se introduce aer in cele doua compartimente ale capsulei oscilometrice, care comunica intre ele si in manseta, la o presiune superioara celei diastolice. Intrerupand comunicarea, apar oscilatiile supramaximale egale, de amplitudine mica, datorate izbirii marginii superioare a mansetei de unda pulsatila. Decomprimand in trepte din 10 in 10mm Hg, intre 120-140mm Hg apare o oscilatie de amplitudine superioara care corespunde presiunii arteriale diastolice. Oscilatiile cresc in amplitudine cu scaderea presiunii si ating o amplitudine maxima ce corespunde presiunii arteriale medii, cu valoarea se 80-90 mmHg. Aceasta nu este media aritmetica a maximei si minimei. La limita dintre ultima oscilatie de amplitudine mai mare si urmatoarele oscilatii mici, egale intre ele, se noteaza presiunea diastolica. Aparatul este utilizat pentru masurarea presiunii medii si a indicelui oscilometric, care exprima extensibilitatea arterelor dela extremitati.
Reglarea circulatiei
Tonusul vascular bazal reprezinta starea de semicontractie a musculaturii netede din vasele de rezistenta. Ea este mentinuta printr-o activitate intrinseca, miogena. Cresterea presiunii arteriale prin mecanism feedback mareste tonusul acestor vase. Acumularea de metaboliti ca urmare a reducerii debitului local exercita asupra vaselor precapilare o activitate locala vasodilatatoare. Reglarea locala a circulatiei prin mecanism miogen si prin metabolism se numeste autoreglare. Ea mentine tonusul bazal al musculaturii netede vasculare si il acomodeaza cerintelor metabolice ale organului. Peste aceste mecanisme de autoreglare pot interveni mecanisme nervoase si umorale. Reglarea umorala a circulatiei 1. Substante vasoconstrictoare. Mediatorii simpatoadrenergici de structura catecolaminica, noradrenalina si adrenalina (prima are efecte vasculare de 3-4 ori mai mari) produc in sectorul microcirculatiei constrictia arteriolelor si venulelor. Fac exceptie vasele musculaturii si inimii si au efect redus in circulatia pulmonara. Ca urmaare are loc o redistribuire a sangelui din organele depozitare, cu cresterea afluxului venos si debitului circulator, activarea functiei cardiace. 2. Substantele vasopresoare de origine renala. Renina este o enzima secretata de rinichi, care produce hipertensiune de durata, cu vasoconstrictie. 3. Vasopresina sau hormonul antidiuretic este vasoconstrictoare numai in doze mari. 4. Ionii de hidrogen si bioxidul de carbon au rol vasodilatator in toate tesuturile, dar cu deosebire in autoreglarea circulatiei cerebrale. 5. Acidul lactic rezultat din metabolizarea anaeroba a glucidelor, intervine in toate teritoriile vasculare ca vasodilatator. 47
6. Acetilcolina este un mediator parasimpatic ce produce vasodilatatie in teritoriile glandulare si in vasele inervate parasimpatic. Are efect de scurta durata. Reglarea nervoasa Nervii vasoconstrictori apartin sistemului vegetativ simpatic. Nervii splanhnici iau nastere din fibrele preganglionare T6-L3 si sunt responsabili de mentinerea unui tonus vasomotor, asigurat de impulsuri eferente cu frecventa de 1-3/secunda. Cresterea frecventei impulsurilor la 10-20/secunda produce o constrictie extrema, iar rarirea lor o dilatatie. Nervii vasodilatatori sunt in functie de mediatorul eliberat parasimpatice colinergice sau simpatice colinergice. Fibrele parasimpatice colinergice vasodilatatoare sunt anexate nervilor cranieni sau au origine in maduva sacrata. Fibrele simpatice colinergice vasodilatatoare se termina mai ales in vasele muschilor scheletici, fiind active in reactia de solicitare. Receptorii cardiovasculari influenteaza primordial activitatea inimii, dar stimularea lor se reflecta si asupra circulatiei sistemice. Sunt importante zona chemo si baro receptorie sinocarotidiana. Baroreceptorii arteriali sistmici (sinusali si ai arcului aortic) se adapteaza lent. O crestere brusca a presiunii arteriale in zona sinusului produce o marire a frecventei impulsurilor in unitatea receptoare. Baroreceptorii arteriali pulmonari sunt lovalizati in ramurile principale ale arterelor pulmonare si sunt sensibili la modificarile de presiune din sursul ciclului cardiac, probabil din cauza distensibilitatii arterei pulmonare. Chemoreceptorii arteriali sunt soncentrati in glomusul carotidian si aortic si reactioneaza lent. La o scadere a presiunii partiale a oxigenului are loc cresterea descarcarii chemoreceptorilor si vasoconstrictie. Centrii vasomotori Centrii vasomotori medulari. Neuronii din coarnele laterale ale maduvei mentin tonusul vaselor si pot declansa reflexe elementare segmentare de vasoconstrictie si chiar vasodilatatie. Acestea pot fi verificate experimental, cand la animalul denervat spinal, dupa un timp tonusul vaselor se reia datorita intretinerii mecanismului de reglare la nivel spinal. Centrul vasomotor bulbar cuprinde totalitatea neuronilor care intervin in functia sistemului respectiv. Centrul vasomotor bulbar este stimulat direct de bioxidulde carbon sangvin ca si centrul respirator. Hipoxia produce stimularea prin acumularea de acid lactic, care mareste concentratia de ioni de hidrogen in neuronii acestui centru. Activitatea centrului vasomotor este subordonata unor centrii suprabulbari din diencefal, sistemul limbic si scoarta cerebrala.
Aparatul digestiv Faringele si Esofagul
Cuprinde mai multe segmente anatomice tubulare, la unele dintre ele fiind anexate si glande secretorii.
48
Aparatul digestiv este format din: cavitatea bucala cu glandele salivare (parotida, submandibulara, sublinguala), faringele, esofagul, stomacul, intestinul subtire (duoden , jejun, ileon), intestinul gros(cec, apendice vermiform, colon, rect) si glandele anexe: pancreasul si ficatul. Tubul sau tractul digestiv este un conduct lung de 10-13m care comunica cu mediul exterior la cele doua extremitati ale sale. Incepe de la fata, prin cavitatea bucala, strabate gatul, toracele, abdomenul, bazinul si termina la anus. Deoarece cavitatea bucala si faringele indeplinesc si functie respiratorie, Nomenclatura Anatomica intelege prin canal alimentar doar esofagul, stomacul, intestinul subtire si intstinul gros. Peretele tubului digestiv constituie a bariera intre mediul extern si organism. Doar prin procese de absorbtie nutrimentele patrund in interiorul organismului , in mediul sau intern. Anexele tubului digestiv cuprind o serie de glande dispuse in lungul acestui conduct si secreta sucuri digestive necesare transformarii alimentelor. Din punct de vedere functional, tubul digestiv poate fi divizat in trei portiuni: 1. portiunea ingestiva deasupra stomacului destinata transportului alimentelor. 2. portiunea digestiva formata din stamac si intestinul subtire, unde alimentele sunt pregatite pentru a fi absorbite. 3. portiunea ejectiva formata din intestinul gros, pe unde resturile digestive sunt eliminate. Faringele este un conduct musculo-membranos, la nivelul caruia se incruciseaza calea respiratorie si alimentara. Bolul alimentar trece in timpul deglutitiei din cavitatea bucala in faringe, apoi in esofag, stomac. Trecerea alimentelor si aerului se face succesiv. Este situat inaintea coloanei cervicale si inapoia foselor nazale si cavitatii bucale. In sus se intinde pana la baza craniului, iar in jos se continua cu laringele si esofagul. Endofaringele este impartit topografic in trei etaje suprapuse: nazofaringe, orofaringe, laringofaringe. Nazofaringele se afla superior, corespunde foselor nazale si se intinde superior pana la baza craniului, iar inferior pana la valul palatin. Orofaringele sau bucofaringele se intinde de la valul palatin superior panala un plan ce trece prin osul hioid. Se continua anterior cu cavitatea bucala iar inferior cu laringofaringele. Laringofaringele este cuprins intre un plan ce trece superior prin osul hioid si alt plan ce trece inferior prin marginea inferioara a cartilajului cricoid al laringelui. Peretele anterior raspunde laringelui. Inferior, laringofaringele se continua cu esofagul. Esofagul este conductul musculo-membranos prin care trec alimentele din faringe in stomac. Limita superioara este planul ce trece prin marginea inferioara a cartilajului cricoid, iar cea inferioara orificiul cardia, prin care esofagul se deschide in stomac. Esofagul strabate regiunea gatului, toracele, diafragmul si ajunge in abdomen, delimitand astfel o portiune: cervicala, toracica sau mediastinala, diafragmatica si abdominala. In stare de vacuitate are aspectul unei panglici musculare. Pe sectiune transversala, lumenul se prezinta ca o fisura cu peretii mai mult sau putin apropiati. Lumenul este virtual la extremitati si are o forma stelata din cauza plicelor mucoasei, in timp ce in rest forma sa este eliptica.
49
In stare de distensie, (dupa insuflare cu aer) arata ca o neregularitate de calibru, prezentand portiuni dilatate si portiuni ingustate sau atramtori fiziologice: 1. stramtoarea cricoidiana sub nivelul cartilajului cricoid, ce raspunde portiunii initialea esofagului. 2. stramtoarea bronhoaortica cauzata de aorta si bronhia stanga. 3. stramtoarea diafragmatica aflata la orificiul esofagian al diafragmului, produsa de constrictia inelului muscular. Intre stramtori exista dilatatii fusiforme.
Stomacul Duodenul Jejunul - Ileonul
Stomacul este portiunea cea mai dilatata a tubului digestiv. Forma reala se bazeaza pe observatia intraoperatorie, radiologica. In postura verticala, stomacul gol are forma litereiJ sau a unei undite, cu o portiune mai lunga, verticala si o portiune mai scurta, orizontala catre dreapta. Limitele se afla la orificiul cardia spre esofag si la orificiul piloric catre duoden. Se descriu doi pereti, doua margini si doua orificii. Marginea dreapta (curbura mica) este concava, continua marginea dreapta a esofagului si prezinta un versant vertical si unul orizontal intre care se contureaza o incizura unghiulara. Marginea stanga este data de curbura mare. Ea este convexa, porneste de la marginea stanga a esofagului impreuna cu care formeaza un unghi ascutit numit incizura cardiala, apoi ocoleste portiunea verticala a stomacului , fundul , corpul si isi schimba directia orientandu-se spre dreapta. Curburile separa peretele anterior de cel posterior. Portiunea verticala a stomacului este mai voluminoasa, dilatata, saculara si cuprinde 2/3 din stomac. Ea se subdivide in portiunea cardica, fund si corp. Portiunea cardica raspunde orificiului cardia pe care il cuprinde. Fundul sau fornixul este portiunea superioara, dilatata a stomacului situata deasupra orizontalei ce trece prin unghiul cardic. Corpul se afla sub nivelul planului amintit anterior si tine pana la un plan vertical ce trece prin incizura unghiulara. Portiunea orizontala este numita si pilorica, se afla inte incizura unghiulara si santul duodeno-piloric. Pilorul este portiunea terminala, stramta a stomacului. Portiunea verticala este cea digestiva, iar cea orizontala evacuatorie. Duodenul este prima portiune a intestinului subtire si se deosebeste de jejun-ileon prin faptul ca este fixat la peretele abdominal posterior. Forma seamana cu un cerc sau potcoava cu deschiderea la stanga. Limita dintre duoden si stomac este santul duodeno-piloric. Limita dintre duoden si jejun este flexura duodeno-jejunala. De la pilor, duodenul se indreapta in sus si spre dreapta spre colul vezicii biliare, unde coteste si formeaza flexura duodenala superioara. Apoi coboara la dreapta capului pancreatic, pana se recurbeaza din nou si formeaza flexura duodenala inferioara. De aici
50
se indreapta transversal peste coloana vertebrala si apoi devine usor ascendent pe flancul stang al coloanei. Se formeaza astfel patru portiuni: I portiunea superioara de la santul duodeno-piloric la flexura duodenala superioara. II portiunea descendenta intre cele doua flexuri duodenale superioara si inferioara. III portiunea orizontala sau inferioara de la flexura duodenala inferioara pana la flancul stang alcoloanei vertebrale. IV portiunea ascendenta pe flancul stang al coloanei vertebrale pana la flexura duodeno-jejunala. Jejun-ileonul este portiunea intestinului subtire cuprinsa intre flexura duodenojejunala si valva ileocecala. Este legat de peretele abdominal posterior prin mezenter. Se deosebeste de duoden prin lungime, numarul mare al curburilor, mobilitate. Jejun-ileonul are o forma cilindroida, cu doua margini, una libera si alta aderenta la mezenter. Cele doua fata difera dupa orientare si situatia anselor. Jejun-ileonul are 14-16 anse, primele fiind orizontale, iar ultimele vertivale. De la flexura duodeno-jejunala intestinul se indreapta inainte si spre stanga. Fiecare ansa este formata din doua ramuri sensibil egale, paralele, incadrand o portiune de mezenter. Un segment al ansei se numeste aferent (orientat spre flexura duodenojejunala) si altul eferent (spre valvula ileocecala). Lungimea jejun-ileonului este de aproximativ 5-6m. Limita dintre jejun si ileon nu este neta. Se admite ca jejunul cuprinde 2/5 din lungimea jejun-ileonului. Elementele de diferentiere sunt de ordin histologic si functional. Jejunul are plice circulre mai numeroase, dar ii lipsesc placile Peyer la nivelul mucoasei. Ileonul are plice circulare mai reduse dar placile Peyer sunt prezente. Placile Peyer sunt o grupare de foliculi limfatici agregati la nivelul mucoasei intestinale. Structurile limfoide intestinale au rol in mecanismele bariere de aparare ale organismului. Intestinul subtire ocupa cea mai mare parte a etajului submezocolic si pelvisului.
Intestinul gros
Inconjoara ca o rama intestinul subtire. Prin dispozitia sa formeaza trei laturi inegale ale unui patrulater. Intestinul gros gros cuprinde: cecul, colonul ascendent, transvers, descendent, sigmoid si rectul. Aceasta impartire se face dupa criteriul asezarii topografice si a schimbarii de directie a diferitelor segmente, precum si dupa modul de comportare a peritoneului. Cecul are o forma saculara, inchis in partea inferioara si se continua superior cu colonul ascendent. Pe fata mediala se insera apendicele vermiform. De la radacina apendicelui pleaca trei tenii musculare care diverg si urca pe cec si apoi pe colon pierzandu-si individualitatea la nivelul rectului. Cecul se afla in fosa iliaca dreapta, pe care nu o ocupa in intregime fiind considerata ca o situatie normala. Exista si cazuri in care acesta poate avea o pozitie inalta (pana in regiunea lombara sau sub ficat) pozitie joasa sau pelvina (coboara in bazin). Poate exista si pozitie ectopica in alte regiuni decat ale abdomenului.
51
Apendicele vermiform este un segment rudimentar al intestinului gros, transformat in organ limfoid. El este implantat in cec la 2-3cm sub deschiderea ileonului si are aspect de tub cilindroid fusiform. De cele mai multe ori descrie o ansa cu concavitatea mediala, este spiralat sau flexuos. In afara acestei situatii poate lua pozitii variate si fata de cec. Oricare ar fi directia apendicelui, punctul de insertie este intotdeauna acelasi. Colonul. Plecat de la valva ileocecala, din fosa iliaca dreapta, colonul urca pana sub fata viscerala a ficatului formand colonul ascendent. Angularea brusca dintre colonul ascendent si cel transvers formeaza flexura colica dreapta. Colonul transvers strabate transversal cavitatea abdominala pana in vecinatatea splinei, unde se creaza o noua angulare, flexura colica stanga. Urmeaza colonul descendent care coboara pana la nivelul crestei iliace. Ultima portiune, colonul sigmoid descrie un arc care strabate fosa iliaca stanga si coboara in bazin, unde se va continua cu rectul. Colonul este fixat de peretele abdominal posterior prin intermediul peritoneului care formeaza un mezou pentru colonul transvers si sigmoid, numite mezocolon si mezosigma, sau direct prin acolare la peretele abdominal posterior la nivelul colonului ascendent si descendent. Rectul este ultima portiune a intestinului gros. Limita superioara este data de punctul de terminare al mezosigmei si prezinta o usoara strangulare, jonctiunea recto-sigmoidiana. Limita inferioara este linia circulara ano-cutanata, situata la jonctiunea dintre tegumentul anal si pielea perineului. Rectul are doua portiuni: -una pelvina, lunga, larga numita ampula rectala. -alta perineala, ingusta, scurta numita canal anal. Ampula indeplineste functia de rezervor in care sunt acumulate materiile fecale, iar canalul anal este un conduct de evacuare al portiunii precedente. Anusul este orificiul prin care rectul se deschide la exterior. Este situat i perineul posterior, in profunzimea santului interfesier. Are forma unei fisuri sagitale, cu doua margini laterale si doua comisuri, anterioara si posterioara. Sta inchis in mod obisnuit si nu se deschide decat la trecerea materiilor fecale.
Pancreasul si Ficatul
Pancreasul este o glanda voluminoasa, cu dubla secretie externa si interna, anexata duodenului. Prin structura, prezinta asemanari cu glandele salivare, motive pentru care i s-a dat si numele de glanda salivara abdominala. Pancreasul este format ditr-o portiune dreapta, orientata vertical, mai voluminoasa numita cap si o portiune orizontala, oblica in sus si la stanga, numita corp. Se termina printr-o coada, iar intre cap si corp segmentul mai ingustat se numeste col. Capul pancreasului se afla asezat in potcoava duodenala, fiind ca si acesta din urma atasat direct la peretele abdominal posterior, peritoneul acoperindu-l doar anterior.
52
Structura glandei este specializata pentru doua functii. Masa principala a glandei are functie exocrina si secreta sucul pancreatic pe care il varsa in duoden. Sistemul canalicular prin care acest suc este transportat ia nastere la nivelul acinilor, apoi aceste canalicule se unesc in ducte colectoare, ce se termina in ductele principal Wirsung si accesor Santorini de unde continutul acestora se varsa in duoden. Partea endocrina a glandei, este dispersata printre tesutul exocrin, nu are canale excretoare si este reprezentata de insulele de celule Langerhans. Acestea secreta doi hormoni: insulina si glucagonul, cu rol in metabolismul glucidelor. Ficatul este cel mai voluminos viscer al abdomenului. Prin caile biliare extrahepatice isi varsa produsul de secretie externa (bila) in duoden. Bila intervine in procesul de digestie prin emulsionarea grasimilor. Sistemul de canale prin care bila este condusa in duoden constituie caile biliare externe. Aflat in hipocondrul drept, ficatul prezinta cele mai importante rapoarte la nivelul fetei viscerale (inferioare). Aici se afla hilul ficatului si o serie de trei santuri dispuse in forma literei H. Doua santuri au directie antero-posterioara, santul sagital drept si stang. Santul sagital stang separa lobul stang de cel drept si prezinta in adancimea lui doua cordoane fibroase rezultate din obliterarea a doua vene din viata intrauterina (ligamentul rotund al ficatului anterior, si ligamentul venos al lui Arantius). Santul sagital drept este larg si format din fosa vezicii biliare anterior si santul venei cave posterior. Al treilea sant, transversal, care le uneste pe cele sagitale poarta numele de hilul hepatic.Acesta contine: vena porta, artera hepatica, ductul hepatic. Vena porta colecteaza si transporta la ficat sangele venos de la organele digestive abdominale. Ea prezinta la cele doua extremitatile cate o retea de capilare. Artera hepatica se distribuie vezicii biliare, parenchimului hepatic. Ductul hepatic ia nestere la iesirea din ficat ca un colector comun biliar, dupa care in grosimea pediculului hepatic primeste ductul cistic, iar rezultatul unirii celor doua, va fi ductul coledoc. Acesta se va deschide in duoden, in imediata vecinatate a canalului principal pancreatic. Motilitatea intestinala Intestinul subtire prezinta mai multe tipuri de miscari: segmentare, pendulare, modificari de tonus, contractii peristaltice, miscari ale vilozitatilor intestinale. Miscarile segmentare sunt contractii inelare, care impart intestinul in segmente. Acestea servesc la amestecarea continutului intestinal si facilitarea absorbtiei. Miscarile pendulare sunt scurtari si alungiri ritmice ale intestinului care se deplaseaza atat in directie caudala cat si orala, asigurand alunecarea anselor una peste alta si amestecarea continutului. Modificarile de tonus. Tonusul este capacitatea musculaturii netede de a-si modifica lungimea fara a modifica tensiunea. Propulsarea continutului intestinal este asigurata de miscarea peristaltica. Undele se transmit intr-o singura directie, dinspre stomac spre anus. Transmiterea unidirectionala a fost explicata prin aceea ca stimularea mecanica a unui segment intestinal determina intotdeauna contractia portiunii suprajacente si relaxarea portiunii subjacente. Miscarile vilozitatilor intestinale apar prin scurtarea si alungirea acestora, ca raspuns la contractiile musculaturii submucoase. Aceasta se contracta independent de
53
stratul muscular circular si longitudinal. Miscarile vilozitatilor imping substantele absorbite in vasele limfatice si produc o agitatie a continutului intestinal.
Digestia
Digestia in cavitatea bucala incepe cu prehensiunea activa, constienta, dupa care urmeaza masticatia. Secretia salivara contribuie la formarea bolului alimentar si degradarea enzimatica a polizaharidelor. Secretia salivara este asigurata de glandele parotida, submandibulara, sublinguala si glandele accesorii ale mucoasei bucale. Acestea secreta o saliva fluida. Saliva indeplineste urmatoarele: -prin amilaza salivara degradeaza amidonul si glicogenul. -prin apa si mucina faciliteaza masticatia si deglutitia. -solubiilizeaza alimentele si stimuleaza receptorii gustativi. -indeparteaza resturile alimentare interdentare. -rol bactericid prin lizozim, Ig A de tip secretor. -rol excretor pentru Pb, Hg, uree (in uremie), in contaminarea virala (eliminarea virala in rabie, poliomielita). -mentine mucoasa bucala elastica. Digestia gastrica Dupa amestecarea alimentelor cu sucul gastric se formeaza chimul gastric ce este eliminat in duoden ca urmare a miscarilor active ale peretilor gastrici. Sucul gastric este elaborat de glandele gastrice care secreta pepsina, acid clorhidric, gastrina si altele. Acirul clorhidric este secretat de celulele parietale care contin mici canalicule care comunica cu lumenul glandelor gastrice. Rolul acidului clorhidric este de a activa pepsinogenul, de a facilita absorbtia Fe si de a exercita o activitate bactericida. Cea mai importanta enzima gastrica este pepsina, secretata in forma inactiva ca pepsinogen si care se activeaza sub actiunea acidului clorhidric. Pepsinele sunt endopeptidaze ce hidrolizeaza legaturile peptidice incare se afla grupari aminice. Rezulta polipeptide cu 6 resturi de aminoacizi. Nucleoproteinele sunt separate in acizi nucleici si proteine. Lipaza gastrica este prezenta doar la copil si hidrolizeaza grasimile fin emulsionate din lapte, galbenus de ou. Lizozimul se afla si in sucul gastric. Factorul intrinsec este o mucoproteina care fixeaza vitamina B12 ce astfel poate fi absorbita. Mucina formeaza un invelis de 1-1,5mm ce adera de peretii stomacului si il protejeaza impotriva factorilor mecanici, chimici. Digestia in intestinul subtire Digestia alimentelor inceputa in cavitatea bucala, continuata in stomac, este finisata in intestinul subtire. Intrucat ph-ul optim de actiune a enzimelor intestinului este neutru, este necesar sa fie redusa aciditatea chimului gastric. Aciditatea chimului se reduce prin schimbul intre ionii de hidrogen si Na, prin peretele intestinal si prin actiunea
54
neutralizanta a sucului pancreatic, bilei si sucului intestinal propriu. La nivelul primei portiuni a intestinului subtire se varsa sucul pancreatic si bila. Secretia pancreatica Pancreasul exocrin este alcatuit din acini glandulari similari cu cei din glandele salivare. Sucul pancreatic se varsa in duoden prin canalul Wirsung si Santorini. In functie de stimulul aplicat, pancreasul secreta doua tipuri de sucuri: un suc foarte alcalin, sarac in enzime si un suc bogat in enzime, dar de volum redus. Secretia pancreatica normala este un amestec intre cele doua tipuri. Contine: -substante anorganice:Na, K, Ca, Mg. -anhidraza carbonica joaca rol in formarea bicarbonatului de sodiu. -substante organice: -tripsina secretata sub forma de tripsinogen, este activata de o enzima eliberata de mucoasa duodenala si intervine in digestia substantelor proteice. -chimiotripsinogenul este convertit in chimiotripsina, proteaza cea mai concentrata din sucul pencreatic ce coaguleaza laptele. -carboxipeptidaza -ribonucleaza care descimpune acizii nucleici in nucleotide, -inhibitorul tripsinei, -lipaza pancreatica activa numai asupra grasimilor emulsionate fin in prealabil de sarurile biliare. Grasimile sunt scindate in acizi grasi si glicerina, mono si digliceride. -amilaza pancreatica actioneaza asupra amidonului sau glicogenului. Secretia biliara. Functiile ficatului: -elimina pe cale biliara pigmentii biliari, colesteralul, acidul colic, metale grele, fosfataza alcalina. -intervine in metabolismull proteinelor, glucidelor, lipidelor, vitamine, substante minerale. -rol in termoreglare, sangele din venele suprahepatice au temperatura cea mai ridicata din organism. -functie antitoxica prin celulele Kupffer care capteaza particule straine patrunse din intestin in circulatie. -in perioada embrionara ficatul are si functie hematopoietica prin formarea elementelor sangvine rosii si albe. -la adult sintetizeaza factorii coagularii si fibrinolizei. Bila este produsul de excretie si secretie hepatica. Nu este un suc digestiv deoarece nu contine enzime (cu exceptia fosfatazei alcaline). Datorita marii puteri de absorbtie a peretilor veziculei biliare, bila veziculara se concentreaza. Bila contine: saruri biliare, pigmenti biliari, colesteral, lecitina. Sarurile biliare se formeaza pe seama colesterolului. Ele au functie; -de detergent, adica de reducere a tensiunii superficiale favorizand fractionarea lipidelor unpicaturi foarte fine, adica emulsionarea. -de activare a lipazelor din intestin. -de favorizare a absorbtiei acizilor grasi prin mucoasa intestinului.
55
Pigmentii biliari constituie produsul de secretie hepatica ce rezulta din descompunerea hemoglobinei. Iau nastere biliverdina, din aceasta bilirubina care este insolubila in apa. Ea circula in sange absorbita pe proteine. La ficat, este detasata si conjugata, forma sub care ajunge in intestin. Acumularea pigmentilor biliari in tesuturi da nastere la culoarea galbena a tegumentelor si mucoaselor, numita icter. Colesterolul este mentinut in suspensie datorita lecitinei si sarurilor biliare. Cresterea raportului colesteral/saruri biliare precipita colesterolul si da nastere la calculi biliari. Administrarea e saruri biliare persoanelor predispuse sa faca calculi ii previne. Desi bila este secretata in mod continuu, se evacueaza discontinuu in perioada de digestie. Intre mese se acumuleaza in vezica biliara, deoarece coledocul se mentine inchis. In momentul contractiei veziculei biliare, se deschide sfincterul coledocian si evacueaza bila, aceste miscari fiind sincronizate cu peristaltica intestinala. Evacuarea biliara este posibila in perioada de ralaxare duodenala. Sucul intestinal contine: -enterokinaza, enzima proteolitica specifica, -fosfataze, -amilaze, maltoza, lactaza, -mucus vascos alcalin ce protejeaza mucoasa. Secretia intestinului gros. Sucul secretat de glandele intestinlui gros este foarte vascoasa, alcalina. In colon nu se secreta enzime digestive. Rolul este de a facilita trecerea materiilor fecale de a proteja mucoasa de iritatiile mecanice si chimice prin neutralizarea acizilor iritanti, formati in urma intensei activitati bacteriene din interiorul colonului. Capacitatea de absorbtie a mucoasei intestinului gros este apreciabila, iar apa este principala substanta absorbita, si pe langa aceasta electrolitii, vitaminele, aminoacizii.
Aparatul urinar
Rinichii sunt organele indispensabile vietii care produc urina. In numar de doi, unul dintre ei poate lipsi, caz i care functia celuilalt devine compensatorie. Au forma de bob de fasole, cu o fata anterioara convexa, o fata posterioara (plana), extremitate superioara rotunjita, extremitate inferioara rotunjita, o margine laterala convexa, o margine mediala usor concava, pe care se fala hilul renal. Hilul este o despicatura pe marginea mediala, marginit de doua buze ( anterioara si posterioara) si este strabatut de elementele pediculului renal. Rinichii se afla retroperitoneal, in profunzimea abdomenului , in fosa lombodiafragmatica. Separarea celor doua portiuni. lombara si diafragmatica este data de insertia diafragmei. Rinichii sunt mentinuti in pozitia lor de mai multi factori: fascia renala. masa adipoasa din jurul acestora, presa abdominala, vasele pediculului renal, peritoneul. Mobilitatea lor normala este in relatie cu miscarilor respiratorii. Rinichiul stang este mai bine fixat decat cel drept. La aceasta mai contribuie presiunea ficatului care in dreapta apasa mai mult rinichiul si corpul pancreasului situat anterior in stanga.
56
Structura rinichiului cuprinde a capsula fibroasa si parenchimul renal alcatuit la randul sau de o substanta periferica numita corticala si profunda, medulara. Parenchimul delimiteaza o cavitate numita sinus renal care contine elementele pediculului renal. Medulara rinichiului este discontinua si formata din mai multe fragmente piramidale inconjurate din toate partile de corticala. Unitatea morfologica si functionala a rinichiului este nefronul, formatiunea secretoare a rinichiului. Nefronul cuprinde: -corpusculi renali situati exclusiv in corticala atat suprapiramidala cat si juxtamedulara. Aceasta contine un glomerul care este un ghem de anse capilare arteriolare care realizeaza filtrarea sangelui la capatul opus glomerulului numit pol urinar. -tubul renal incepe la polul urinar al corpusculului si el produce o prelucrare calitativa si cantitativa a urinei primare. Aceasta prezinta trei segmente: -segmentul proximal, incolacit numit tub contort proximal -segmentul intermediar drept sub forma de ansa ca acul de par, care coboara catre medulara si apoi revine in corticala. -segmentul distal cu o portiune dreapta ascendenta si una incolacita numita tub contort distal. Tubii colectori nu mai apartin nefronilor si sunt situati in medulara. Au rol de colectare si excretie catre papilele urinare si apoi caile excretoare propriu-zise. Caile excretoare urinare sunt alcatuite din: calice mici, calice mari, pelvis renal, ureter, vezica urinara, uretra. Calicele mici sunt tuburi musculomembranoase in sinusul renal care aduna urina de la papilele renale (dinspre medulara) in conductte mai mari numite calice mari. Calicele mari sunt trei palnii musculomembranoase aflate in sinusul renal, care se unesc si formeaza pelvisul renal. Pelvisul renal sau bazinetul este o formatiune musculomembranoasa dilatata, in forma de palnie turtita, care colecteaza urina din calicele mari, iar prin varf se continua cu ureterul. Ureterul este un conduct urinar lung, intins de la pelvisul renal la vezica urinara, care strabate portiunea abdominala si pelvina fiind retroperitoneal. Portiunea abdominala a ureterului coboara aproape vertical retroperitoneal, pentru ca in vecinatatea coxalului sa se incurbeze medial si spre anterior, formand portiunea pelvina. Vezica urinara este un rezervor membranos in care urina secretata continuu de rinichi, este adusa prin uretere. Urina se acumuleaza in intervalul dintre mictiuni si apoi este eliminata la exterior prin uretra. Vezica este un organ pelvin, extraperitoneal, inapoia oaselor pelvine. Are o forma piramidala, cu un varf, fund, col si corp. Vezica goala este turtita si greu de demonstrat. Vezica plina ia forma unui ovoid cu baza in jos si varful in sus, depasind astfel stramtoarea superioara a pelvisului pentru a patrunde in abdomen. Prezinta o fata anterioara, posteroara, separate prin margini laterale si o fata inferioara sau fundul. Vezica plina imbraca forma unui ovoid, cu baza in jos si inapoi cu varful in sus si inainte. Pe masura ce se umple, axul sau tinde spre verticalitate. Spre deosebire de cea goala, marginile laterale ale vezicii pline devin fete laterale.
57
Suprafata interioara a vezicii urinare prezinta aceleasi segmente ca si suprafata externa. Varful este reprezentat printr-o depresiune. Fundul este segmentul cel mai important. Cuprinde trigonul vezical si fosa retroureterica. Trigonul vezical este o regiune neteda, plana, lipsita de plice, cu baza intre cele doua orificii ureterale si varful la orificiul intern al uretrei. Fosa retroureterica este o depresiune transversale situata inapoia plicei interureterice (dintre cle doua orificii ureterale). Orificiul intern al uretrei constituie in mod normal punctul cel mai decliv al vezicii. Corpul vezical are un aspect interior neregulat, cu coloane sau plice anastomozate. Uretra este conductul care face legatura dintre vezica urinara si exterior. La barbat are trei portiuni(prostatica, membranoasa sau diafragmatica si spongioasa) iar la femeie o portiune pelvina si una perineala. In uretra se deschid la barbat orificiile glandelor bulbouretrale, iar la femeie orificiile glandelor uretrale. Vascularizatia rinichiului Artera renala se divie in interiorul rinichiului in artere interlobare apoi artere arciforme care merg intre corticala si medulara, paralal cu suprafata organului. Din ele pleaca ramuri perpendiculare spre suprafata organului numite artere interlobulare. Acestea la randul lor emit artere glomerulare aferente. La iesirea din glomerul, artera eferenta re un calibru mult mai mic si se divide intr-o noua retea capilara care hraneste tubii contorti, dupa care se varsa in venele renale. Arteriola eferenta se ramifica intr-un sistem de vase subtiri (vasa recta) care urmeaza traiectul anselor (segmentelor intermediare0 inainte de a conflua in venele renale. Acest aspect este legat de unul din mecanismul de concentrare a urinii. Rinichiul prezinta astfel circulatie nutritiva si functionala. Limfaticele sunt absente.
Formarea urinii
Functia glomerulara. Punctia capsulei glomerulare si analiza urinii primare, rezultata imediat dupa filtrarea sangelui arterial prin membrana glomerulara, demonstreaza ca ea contine doar urme de proteine. Pana la greutatea moleculara de 55000 (inulina) coeficientul de concentrare filtrat/plasma este de 1. Peste aceasta valoare are loc procesul de cernere moleculara, in sensul de scadere progresiva a permeabilitatii spre albumina cu masa moleculara de 70 000. Filtrarea glomarulara depinde de: -structura si suprafetele membranei filtrante. -presiunea efectiva de filtrare. -debitul circulator renal. Functiile tubulare Urina primara, in scurgerea ei prin tubii renali sufera procese de resorbtie si secretie. De la volumul de 180l, filtrat glomerular? 24 de ore, volumul urinei definitive devine de peste 100 de ori mai concentrat, adica 1,5l urina/ 24 de ore. Reducerea de volum se face pe seama resorbtiei active de Na urmata pasiv de resorbtia apei. Transportul apei si ionilor in diferite segmente ale nefronului. La sfarsitul primei portiuni a tubului contort proximal, coeficientul de concentrare urina/plasma este de trei, 58
ceea ce inseamna ca in aceasta zona se resorb 2/3 din apa filtratului glomerular, in aceeasi proportie cu sarurile, lichidul ramanand izoton. La inceputul tubului contort distal urina tubului este puternic hipotona, pentru ca in urma pierderii apei devine izotona spre sfarsitul tubului contort distal si hipertona in tubii colectori. -Na este resorbit obligator in tubul contort proximal. -Cl se comporta la fel. -Ca neproteic se absoarbe ca si sodiul. -K este resorbit in tubul proximal in cea mai mare parte, iar apoi in tubul contort distal. Transportul ionilor prin epiteliul tubular nu poate fi explicat numai fizico-chimic intrucat se face impotriva unei diferente de potential. Glucoza in conditii normale nu apare in urina. Peste 180mg% incepe eliminarea urinara, care creste progresiv, dar mai lent decat concentratia sangvina. Cantitatea de glucoza resorbita maximal constituie transportul tubular maximal. Fosfatii sunt resorbiti in conditii normale in tubul contort proximal. Fosfatii eliminati prin urina au rol de tampon ph in tubul contort distal contribuind la retinerea sodiului in sange. Sulfatii se comporta ca si fosfatii. Macromoleculele. Epiteliul tubului contort proximal, in afara de resorbtia unor micromolecule, are proprietatea de a capta si retine macromolecule filtrate glomerular cum ar fi proteine, coloranti macromoleculari. Eliminarea de proteine prin urina peste 200mg/24 de ore se nmeste proteinurie. Ea se datoreaza cresterii permeabilitatii filtrului glomerular, cu depasirea capacitatii de retinere a epiteliului tubular. Substante care se filtreaza si apoi se excreta cuprind in acet grup multe medicamente care se leaga partial de proteine, forma sub care nu se filtreaza sau intra in competitie de secretie cu substante inrudite ca structura. Mecanismul de transport activ in acest caz consta in formarea la suprafata celulei a unui compus care este transportat in interiorul celulei si de aici spre polul opus , de unde prin descompunere substanta de transportat este eliberata in urina. Procesul necesita energie. Mecanismul de concentrare si diluare a urinii. Masurand presiunea osmotica in adancimea piramidelor renale se observa cresterea progresiva dinspre baza spre varf sau in sens corticomedular. In ramura ascendenta a ansei are loc resorbtiaa activa a sodiului urmata de clor. Ansa este impermeabila pentru apa. Ca urmare, urina la capatul ramurei ascendente va deveni hipotona iar spre medulara hipertona. Fluxul urinar antreneaza spre varful piramidei ionii astfel difuzati, marind concentratia osmotica in aceasta zona. Anselor le revine rolul de a realiza un gradient osmotic intre baza piramidei si varf prin resorbtia activa a sodiului in ansa ascendenta. La sfarsitul ansei ascendente urina devine hipotona. La sfarsitul tubului contort distal redevine izotona. De-alungul tubilor colectori permeabili pentru apa, urina deplasandu-se spre un gradient osmotic marit, apa se resoarbe, concentrand urina finala. Mictiunea Din canalele colectoare urina trece in calice, bazinet si prin intermediul ureterelor in vezica urinara.
59
Deplasarea in uretere este posibila datorita contractiilor ritmice, printr-un automatism similar celui cardiac. La locul de varsare al ureterelor in vezica se afla cate o mica valva, care permite trecerea urinei intr-un singur sens si nu refluxul. Anatomic, vezica urinara prezinta trei straturi musculare: extern longitudinal, mijlociu circular, intern reticular longitudinal. La nivelul colului se formeaza un sfincter intern, neted. Pe masura ce urina se acumuleaza in vezica, volumul ei creste. Presiunea exercitata asupra peretilor, excita receptorii din mucoasa si determina reflex, relaxarea de adaptare. Treptat apare senzatia de mictiune. Daca individul nu consimte actul reflex, vezica se relaxeaza din ou in oarecare masura. Cand cantitatea de lichid acumulata atinge maximum admisibil, apare senzatia imperioasa de mictiune insotita de durere. Sfincterul extern al vezicii este format din fibre musculare striate. Este inervat de sistemul nervos cerebrospinal supus vointei. Sfincterul extern intervine cand individul nu consimte sa urineze.
Aparatul genital feminin
Ovarele sunt glandele sexuale ale femeii, care determina caracterele sexuale si produc ovule. Acestea au functie endocrina prin secretia de hormoni sexuali. Ovarele impreuna cu tubele si ligamentele alrgi sunt numite de clinicieni anexe. Ovarul are forma de ovoid putin turtit, dispus cu axul mare aproape vertical, cu o fata mediala si laterala, doua margini (libera si mezovariana) si doua extremitati (tubara si uterina). La nou-nascuta, ovarul este albicios, roz palid la fetita, rosiatic la femeia adulta, iar dupa menopauza redevine albicios-cenusiu. Ovarul se afla in cavum retrouterin, adica compartimentul cavitatii pelvine aflat inapoia ligamentelor largi, alipit de peretele lateral al excavatiei, sub bifurcatia arterei iliace comune. Ovarul este suspendat de ligamentul alrg si fixat in mod relativ prin: ligamantul suspensor, ligamentul proriu al ovarului, ligamentul tubo-ovarian. Ligamentul suspensor numit si infundibulo-pelvic este cel mai puternic si pleaca din fosa iliaca, strabate stramtoarea superioara a pelvisului si patrunde in unghiul superolateral al ligamentului larg, unde se fixeaza pe extremitatea tubara a ovarului. Ligamentul propriu sau uteroovarian este un cordon fibromuscular, care se intinde de la extremitatea uterina a ovarului la unghiul uterului, atasat de ligamentul larg. Ligamentul tuboovarian leaga extremitatea tubara a ovarului de infundibulul tubei. De el se ataseaza un ciucure tubar numit fimbria ovarica. Mezovarul este o plica a ligamentului larg prin care ovarul este suspendat la fata posterioara a acestuia. Prin marginea mezovariana a ovarului patrund elemente pediculare prin hil in organ. Structura. Pe o sectiune, ovarul prezinta de la suprafata in profunzime, un epiteliu sub care se afla un invelis conjunctiv, o zona periferica corticala si o zona centrala medulara. Epiteliul este simplu si se opreste brusc la nivelul hilului.
60
Invelisul conjunctiv se numeste albuginee, se continua fara delimitare cu stroma corticalei. Substanta corticala contine elemente caracteristice; foliculi ovarieni in diferite grade de evolutie. Ovocitele a caror prezenta este obligatorie, vor induce in cursul vietii sexuale active a femeii diferentierea si maturarea elementelor celulare, care culmineaza cu eliberarea unuia sau mai multor ovule mature, proces numit ovulatie, lunar. Substanta medulara prezinta o structura intens vascularizata. Functia endocrina a ovarului se instaleaza la pubertate si este sustinuta de celule corticale secretoare de hormoni specifici: estrogen si progesteron, sub controlul hormonilor gonadotropi, stimulatori si inhibitori.
Tubele uterine sunt doua conducte musculo-membranoase care se intind de la coarnele uterine pana la ovare. Au rolul de a capta ovulul, de a-l transporta si a oferi mediul necesar fecundarii atunci cand este cazul. Tuba prezinta o lungime de 10-13 cm si are patru portiuni care dinspre lateral spre medial sunt: infundibulul, ampula, istmul si portiunea uterina. Infundibulul sau pavilionul este segmentul lateral cu forma de palnie cu baza evazata, foarte festonata, cu mai multe fimbrii sau ciucuri. Unul dintre ei este mai alungit si numit fimbria ovarica, si insoteste ligamentul tubo-ovarian avand rol de dirijare a ovulului. Ampula este segmentul cel mai lung, ocupa 2/3 din lungimea totala si se ingusteaza progresiv spre medial avand un traiect ansiform in jurul ovarului. Istmul este portiunea ingusta, rectilinie, care patrunde in cornul uterului. Portiunea uterina sau interstitiala strabate grosimea peretelui uterin si este scurta. Orificiile tubare sunt doua: -ostiul abdominal in centrul infundibulului. -ostiul uterin care se deschide in unghiul cavitatii uterine. Datorita acestor comunicari o endometrita se poate complica cu o salpingita si apoi pelviperitonita. Tubele uterine sunt in continuitate cu uterul, situate in marginea superioara a ligamentelor largi, in mezosalpinge (mezoul tubei sau salpingelui). Tuba impreuna cu mezosalpingele cad in cavum retrouterin, prezentand o mobilitate mai mare in portiunea ei laterala.
Aparatul genital feminin - Uterul
Uteruleste un organ muscular, cavitar, mepereche in care se desfasoara gestatia. Are forma unui trunchi de con turtit antero-posterior cu baza orientata in sus si varful trunchiat in jos. Istmul uterin este un sant semicircular, doar pe fata anterioara. Corpul uterului prezinta doua fete ( una viscerala si intestinala), doua margini (dreapta si stanga), fundul si doua unghiuri tubare.
61
Colul uterului are forma cilindrica, usor bombat. extremitatea superioar a vaginului se insera pe col de-alungul unei linii circulare oblice, situata mai sus posterior si mai jos anterior. In acest fel, aceasta insertie imparte colul intr-o portiune supravaginala (cilindrica) si o portiune intravaginala (tronconica). Varful este perforat de un orificiu numit ostiu uterin care conduce prin canalul cervical in cavitatea uterina. Uterul este situat in centrul cavitatii pelvine, inapoia vezicii urinare, inaintea rectului , deasupra vaginei, iar corpul nu depaseste planul stramtorii superioare a bazinului. In mod normal, uterul se afla in pozitie de anteversie-anteflexie, luand de fapt la nivelul corpului o inclinatie aproape orizontala. Datorita mobilitatii organului, poate fi deplasat in conditii fiziologice de organele vecine in functie de starea lor de plenitudine sau in conditii patologice de procese inplamatorii, prin aderente, caz in care pozitia anormala devine fixata, constanta. Mijloace de fixare: peritoneul, ligamentele largi, ligamentele rotunde, aderentele la vazica urinara si rect, perineul. Ligamentele largi sunt doua cute peritoneale patrulatere, aflate intre marginile uterului si peretii laterali ai excavatiei pelvine. In marginea superioara a ligamentelor largi se afla tuba uterina. In partea inferioara, cele doua fooite diverg, iar directia in totalitate urmeaza directia corpului uterin. Inportiunea superioara a ligamentului larg, sub tuba se formeaza o zona mai subtire si mobila a acestui ligament, numita mezoul salpingelui (tuba) numit mezosalpinge. Pe fata posterioara a ligamentului larg, sub mezosalpinge, se ataseaza marginea mezovariana a ovarului, loc de unde ligamentul larg trimite o scurta prelungire numita mezoul ovarului sau mezovar. Sub acest nivel, ligamentul devine mai gros, foitele anterioara si posterioara se indeparteaza si formeaza mezoul uterului (mitra) numit si mezometru. Ligamentul rotund este un cordon conjunctiv pereche, care pleaca de la unghiul tubar al uterului , strabate ligamentul larg, parcurge peretele abdominal anterior la nivelul canalului inghinal si apoi se ancoreaza prin travee conjunctive in regiunea prepubiana si labiile mari. Aceasta participa la realizarea pozitiei uterului de anteversie-anteflexie. Cavitatea uterina are o forma triunghiulara, cu baza la fundul organului si varful spre canalul cervical. Cele trei unghiuri sunt marcate de trei orificii: superioare sau laterale corespunzatoare deschiderii tubelor si inferior , orificiul cervical intern.
Canalul cervical se afla in interiorulcolului si prezinta o regiune mijlocie fusiforma si doua orificii: unul superior orificiul cervical intern spre cavitatea uterina, si orificiul cervical extern care este chiar ostiul uterin si se deschide in vagin.
Aparatul genital masculin si glanda mamara
62
Este reprezentat de glandele genitala: testiculele, conductele genitale si glandele seminale. Testiculul este glanda genitala masculina care indeplineste functia de spermatogeneza si o functie de glanda endocrina prin secretia de hormoni ce imprima caracterele sexuale secundare masculine. Testiculul este situat in scrot, la nivelul perineului anterior si prezinta o forma ovoida, cu o fata laterala, mediala, extremitate superioara si inferioara, margine anterioara si poaterioara. Marginea posterioara vine in raport cu epididimul si da trecere vaselor si nervilor proprii, motiv pentru care se considera a avea rol de hil. Epididimul este o formatiune alungita ce se compune din cap, corp, coada cu dispunere descendenta fata de marginea posterioara a testiculului si care se continua apoi cu ductul deferent fara nici o limita. Structura testiculului prezinta o lobulatie conjunctiva ce contine formatiuni tubulare incolacite numite tubi seminiferi contorti, care se aduna apoi treptat catre hilul organului in tubi seminiferi drepti ce apoi se deschid in epididim si ductul deferent. Dealungul acestor formatiuni tubulare are loc spermatogeneza si maturarea elementelor specifice. Ductul deferent este un conduct lung, care pleaca de la coada epididimului, strabate canalul inghinal si grosimea peretelui abdominal anterior, apoi se incurbeaza catre pelvis si se termina la baza prostatei unde se uneste cu ductul excretor al veziculei seminale. La unirea celor doua se formeaza ductul ejaculator, care se deschide in uretra prostatica. Veziculele seminale sunt organe pereche, cu rol secretor, produsul adaugandu-se lichidului seminal. Prostata este o glanda situata inferior de vezica urinara, dezvoltate in jurul portiunii initiale a uretrei. Forma clinica, rezultata la tuseul rectal este de as de pica ale carui extremitati superioare, coarnele se continua cu veziculele seminale, iar varful cu uretra. Un sant median, usor de perceput, situat pe ata posterioara, marcheaza limita dintre cei doi lobi laterali ai prostatei. Glanda mamara Mamelele sau sanii sunt organe glndulare de origibe cutanata, asezate simetric in regiunea toracica anterioara la om. Este posibila existenta si de mamele supranumerare de-alungul unei linii care pleaca din axila si pana in regiunea inghinala. La om, frecvent se dezvolta doar o singura pereche de glande care ia proportii diferite dupa sex si functie. Areola este o prelungire circulara de culoare variabila, roz-maronie in portiunea cea mai proeminenta, cu diametru de 2,5-3cm. La suprafata sa se afla 10-15 proeminenta neregulate, numite tuberculi Morgagni, determinati de glande sebacee subjacente. Papila mamara sau mamelonul este o proeminenta cilindrica sau conica, cu varful rotunjit, baza mai larga in centrul areolei. Pe varful ei se deschid 15-25 ducte lactifere prin orificii mici. Stratul celulo-adipos premamar sau anterior este mai gros la periferie si se subtiaza spre centru.
63
Corpul mamelei este o formatiune discoidala, care prezinta o portiune periferica moale, vascularizata, friabila, formata din parenchim glandular si o portiune centrala albicioasa, rezistenta, formata predominant din ducte excretoare. Stratul celulo-grasos retromamar este subtire intre fata posterioara a corpului mamelei si fascia muschilor peretelui toracicanterior, fata de care in mod normal exista un grad de mobilitate.
Fiziologia receptorilor feminini
Toate structurile organismului feminin sufera modificari sub influenta hormonilor steroizi sexuali, efecte generale metabolice si efecte specifice in relatie directa cu concentratia receptorilor hormonali continuti de celulele din diferite organe si tesuturi. Organele tinta prin concentratia crescuta de astfel de receptori, prezinta modificari specifice sub influenta hormonilor ovarieni. Organele genitale feminine se matureaza sub influenta hormonilor sexuali, iar la femeia adulta se constata modificari ciclice care realizeaza ciclul uterin (cervical, endometrial, miometrial) vaginal, tubar, mamar in raport cu ritmul secretor de estrogeni, progesteron, androgen. Explorarea acestor receptori permite o evaluare a functiilor normale ovariene. Ciclul endometrial Endometrul este format dintr-un invelis epitelial, glande si corionul conjunctiv cu vase si nervi la vaza. Fazele ciclului endometrial sunt: 1. Faza proliferativa (estrogenica) care incepe la sfarsitul singerarii menstruale (aprox. ziua 5 a ciclului menstrual) si se termina in ziua ovulatiei aprox. ziua 14 a unui ciclu de 28 de zile. Se caracterizeaza prin fenomene proliferative determinate de hormonii estrogeni ovarieni, care intereseaza toate structurile endometriale. 2. Faza secretorie (estroprogesteronica) incepe dupa ovulatie si este determinata de stimularea endometrului prin estrogeni si progesteronul produs de corpul galben si se caracterizeaza prin fenomene de maturizare celulara si activitate secretorie. 3. Faza menstruala indusa de scaderea rapida a nivelurilor estro-progesteronice in lipsa fecundatiei si ovoimplantatiei, care constituie prabusirea unui edificiu destinat instalarii unei sarcini. Incepe cu un stadiu premenstrual ischemic in zilele 27-28 si continua cu perioada de descuamare si regenerare in zilele 1-4 ale ciclului endometrial. Ovogeneza Ovarela iindeplinesc doua functii esentiale: -ovogenetica (gametogenaza) prin care furnizeaza in fiecare ciclu menstrual cate un ovul matur eliberat pentru a fi fecundat, -hormonala (glanda endocrina) prin care secreta hormoni steroizi ce pregatesc receptorii genitali pentru fecundatie. Ovarul se diferentiaza inca din primala 10-12 saptamani ale vietii intrauterine. Inca din saptamana 10-12 din gonocitele primordiale se constitnuie aglomerari celulare formate dintr-o celula centrala derivata din ganacitul primordial numita ovogonie, si celule turtite la periferie numite granuloase sau foliculare. Gonocitele primordiale se
64
inmultesc si ajung la 5-7 milioane in luna a V-a intrauterina, dupa care o mare parte sufera procese de atrezie, incat la nastere numarul final este de 1-2 milioane. Concomitent cu multiplicarea, ovogoniile sufera si un proces de maturizare transformandu-se in ovocite care isi incep diviziunea reductionala. La nastere, ovocitele se afla oprite in aceasta diviziune reductionala. In jurul ovocitelor se aduna unul sau mai mmulte straturi de celule granuloase sau foliculare, formand foliculul primar. La pubertate ajung 300 000 foliculi din 1-2 milioane de la nastere. Dezvoltarea foliculilor in cursul ciclului ovarian. Foliclul primordial este format ditr-un ovocit inconjurat de un sungur strat de celule foliculare. Foliculul primar este un ovocit inconjurat de mai multe straturi de celule foliculare si aparitia unui spatiu cavitar perivitelin, Foliculul cavitar mic mijlociu mare, in cursul maturizarii, in mijlocul celulelor granuloase se acumuleaza un produs de secretie al acestora, numit lichid folicular intr-o cavitate centrala ce creste trptat in dimensiuni. In foliculii mari, prin cresterea cavitatii foliculare, ovulul este impins excentric si intreg foliculul spre suprafata ovarului. Celulele foliculare care captusesc peretele folicular constituie celulele prietale. Acumulare de celule foliculare intre ovul si peretele foliculului formeaza cumulus proliger sau discul proliger. Stratul de celule care inconjoara ovulul in afara membranei pellucida se numeste coroana radiata. Foliculul matur de Graaf are dimensiuni de 15-25mm si este format di exterior spre interior din: -teaca externa formata din tesut conjunctiv densificat, cu vase sangvine, limfatice, fibre musculare netede. -teaca interna formata din celule mari, -membrana Kolliker Slawjansky care desparte invelisul tecal vascularizat de mambrana granuloasa. -membrana granuloasa formata din celule granuloase sau foliculare aranjate in strat parietal, cumulus proliger, coroana radiata. -lichid folicular. Ovulatia este dehiscenta foliculului matur, cu eliminarea ovulului inconjurat de 68 straturi de celule granuloase care constituie coroana radiata si perfectarea meiozei (prima diviziune reductionala. Ovulul inconjurat de celulele granuloase este captat de fimbriile trompei prin proiectiuni delicate si miscari peristaltice estrogen-induse ale ampulei tubare in asociere cu miscarile cililor vibratili tubari ce ating in aceasta faza dezvoltarea si activitatea motorie maxima. Meioza initiata in viata intrauterina (saptamana 12) trece intr-o perioada de inghetare pana la instalarea ciclurilor ovulatorii. La ovulatie, in perioada de activitate sexuala se perfectioneaza diviziunea reductionala din cadrul meiozei si se formeaza doua celule fiecare nu numar haploid de cromozomi (22X) dar cu citoplasma inegala: ovocitulde ordinul II care pastreaza aproape toata citoplasma celulei mame si primul globul polar care este expulzat in spatiul perivitelin.
65
A doua diviziune de maturare incepe imadiat dupa perfectarea meiozei, este de tip diploid si are loc abia in momentul fecundatiei. Temporal, meioza precede cu 2-4 ore dehiscenta foliculului, iar ovulele eliminate spontan sau extrase cu laparoscopul sau prin punctia foliculilor si car sunt fecundabile in vitro, se gasesc in cursul celei de a II-a diviziuni, de maturatie. In urma celei de a doua diviziuni de maturatie, ia nastere ovulul fecundant si cel de al doilea globul polar care se pierde.
66
S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Anatomie 2 - CelulaDocument7 paginiAnatomie 2 - CelulaDariusNastaÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Anatomie Oculara CursDocument82 paginiAnatomie Oculara Cursmihaitoma72Încă nu există evaluări
- NEUROPSIHOLOGIE: elementele de bază ale problemeiDe la EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiEvaluare: 5 din 5 stele5/5 (1)
- Curs 2 Anatomie - Celula, Tesuturi, Sisteme-APDocument21 paginiCurs 2 Anatomie - Celula, Tesuturi, Sisteme-APMinerva TutoveanuÎncă nu există evaluări
- Celula Animală ContinuareDocument33 paginiCelula Animală ContinuareNicoleta CobanÎncă nu există evaluări
- Celula Tipuri TesuturiDocument8 paginiCelula Tipuri Tesuturiandreea barosanÎncă nu există evaluări
- Celula Si TesuturiDocument10 paginiCelula Si TesuturiNastea SilivestruÎncă nu există evaluări
- Suport de Curs FiziologieDocument20 paginiSuport de Curs FiziologiechivumihaelaÎncă nu există evaluări
- Curs - ANATOMIA SI FIZIOLOGIA ANIMALELOR OdtDocument42 paginiCurs - ANATOMIA SI FIZIOLOGIA ANIMALELOR OdtAlexandra PenciuÎncă nu există evaluări
- Curicullum BFTDocument7 paginiCuricullum BFTVash da StampidoÎncă nu există evaluări
- Celula Si Organitele CelulareDocument4 paginiCelula Si Organitele CelulareMonica AoaneiÎncă nu există evaluări
- Celula (Structură, Proprietăţi)Document37 paginiCelula (Structură, Proprietăţi)Sorina DobreÎncă nu există evaluări
- Anatomie CondensatDocument7 paginiAnatomie CondensatAlex IacobÎncă nu există evaluări
- CelulaDocument7 paginiCelulaAndrada AndrutzaÎncă nu există evaluări
- Curs 2, 3 Anatomie AmgDocument15 paginiCurs 2, 3 Anatomie AmgAlexandru GiubegaÎncă nu există evaluări
- Organizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternDocument4 paginiOrganizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternVata LuminitaÎncă nu există evaluări
- Celula Și Fiziologia CelularăDocument16 paginiCelula Și Fiziologia Celularăzilahivanessa799Încă nu există evaluări
- Ce Este CelulaDocument4 paginiCe Este CelularalucaÎncă nu există evaluări
- Tema - ,,celula - Unitate Structural - Funcțională A Organismului UmanDocument12 paginiTema - ,,celula - Unitate Structural - Funcțională A Organismului UmanIon MosnoiÎncă nu există evaluări
- Celula Și Fiziologia CelularăDocument26 paginiCelula Și Fiziologia CelularăMARIA-CASANDRA VOICUÎncă nu există evaluări
- Anatomie - Curs1 - CelulaDocument27 paginiAnatomie - Curs1 - Celulaalfa5017Încă nu există evaluări
- BiologieDocument23 paginiBiologieCostÎncă nu există evaluări
- Celula AnimalaDocument10 paginiCelula AnimalaRaluka Siliște80% (5)
- Anatomia Si Fiziologia OmuluiDocument6 paginiAnatomia Si Fiziologia Omuluimirabela100% (2)
- Anatomia Si Fiziologia Omului 191746Document3 paginiAnatomia Si Fiziologia Omului 191746murguÎncă nu există evaluări
- Fundamente BiologiceDocument17 paginiFundamente BiologiceElena MoiseÎncă nu există evaluări
- Organite Celulare ComuneDocument13 paginiOrganite Celulare ComuneDennisÎncă nu există evaluări
- Bio - Cap I - Alcatuirea Corpului UmanDocument8 paginiBio - Cap I - Alcatuirea Corpului UmanAngie Monoranu100% (1)
- Proiect BiologieDocument13 paginiProiect BiologieFlorinAndrabuleaÎncă nu există evaluări
- CelulaDocument14 paginiCelulaFirma GSCÎncă nu există evaluări
- Cito HistoDocument10 paginiCito HistoGabriela TanasescuÎncă nu există evaluări
- Celula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Document1 paginăCelula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Gabor IzabellaÎncă nu există evaluări
- Celula Animala Si Celula VegetalaDocument4 paginiCelula Animala Si Celula Vegetalaalinutza_doltenitzaÎncă nu există evaluări
- Anatomia Si Fiziologia Omului - 240109 - 155131Document20 paginiAnatomia Si Fiziologia Omului - 240109 - 155131Alex IacobÎncă nu există evaluări
- Material MedicinaDocument143 paginiMaterial MedicinaAna Matei100% (1)
- Celula Unitatea Morfofuncţionala A Organismelor VIIDocument7 paginiCelula Unitatea Morfofuncţionala A Organismelor VIIlaklod100% (1)
- Scoala Postliceala Sanitara Arad: Unitatea de InvatamantDocument5 paginiScoala Postliceala Sanitara Arad: Unitatea de InvatamantDannyela BuntaÎncă nu există evaluări
- Curs 2 FegDocument4 paginiCurs 2 FegFLORIN STEFAN BARBUÎncă nu există evaluări
- Carte Fizio Farasemnatura-1-1Document274 paginiCarte Fizio Farasemnatura-1-1CIPSITÎncă nu există evaluări
- Adrian's BiologyDocument139 paginiAdrian's Biologycoso1994Încă nu există evaluări
- Curs 2 CelulaDocument12 paginiCurs 2 Celulaaminalhassan963Încă nu există evaluări
- Celula Animala vs. VegetalaDocument14 paginiCelula Animala vs. VegetalaCristina Gabriela100% (1)
- Tesuturile Si Celula AnimalaDocument8 paginiTesuturile Si Celula AnimalaAndreea Ștefura0% (1)
- Celula Animala vs. Celula VegetalaDocument13 paginiCelula Animala vs. Celula VegetalaCristina GabrielaÎncă nu există evaluări
- Cap 3Document8 paginiCap 3Marfescu CristianÎncă nu există evaluări
- Tema 2Document10 paginiTema 2bella17Încă nu există evaluări
- WWW - Referat.ro-Tesuturi Si Celula30268e3fDocument8 paginiWWW - Referat.ro-Tesuturi Si Celula30268e3fAaron Wolfe100% (1)
- Celula AnimalaDocument13 paginiCelula AnimalaMina YoungÎncă nu există evaluări
- Anatomia Si Fiziologia OmuluiDocument38 paginiAnatomia Si Fiziologia OmuluiAlfredDoruÎncă nu există evaluări
- Anatomie Si Fiziologi1Document5 paginiAnatomie Si Fiziologi1Ion PahomiÎncă nu există evaluări
- Organizarea Corpului UmanDocument4 paginiOrganizarea Corpului UmanIuliaNaTuFaruÎncă nu există evaluări
- Cursuri Biocel CDDocument112 paginiCursuri Biocel CDKovacs AnettÎncă nu există evaluări
- CelulaDocument3 paginiCelulaIacob Paula-IzabelaÎncă nu există evaluări
- C2-Niveluri de Organizare Generală Ale Corpului UmanDocument7 paginiC2-Niveluri de Organizare Generală Ale Corpului UmanlauraÎncă nu există evaluări
- Anatomie CelulaDocument8 paginiAnatomie CelulaLavinia DobrescuÎncă nu există evaluări
- Celula AnimalăDocument1 paginăCelula Animalădiana scoalaÎncă nu există evaluări
- Cito PlasmaDocument4 paginiCito PlasmaAndreea ValcuÎncă nu există evaluări
- Patologie Celulara - 2 - Celula - Organizare Si FunctiiDocument24 paginiPatologie Celulara - 2 - Celula - Organizare Si FunctiiSimina AnamariaÎncă nu există evaluări
- Curs de Neonatologie CarteDocument218 paginiCurs de Neonatologie CarteIvan DoruÎncă nu există evaluări
- 53 Laxativele Si PurgativeleDocument3 pagini53 Laxativele Si PurgativeleRoxana ClimescuÎncă nu există evaluări
- Curs Valvulopatii AorticeDocument28 paginiCurs Valvulopatii AorticeIoana MuntianuÎncă nu există evaluări
- Diagnostic DiferentialDocument3 paginiDiagnostic DiferentialIoana MuntianuÎncă nu există evaluări
- Grile La Materia ToxicologieDocument2 paginiGrile La Materia ToxicologieIoana MuntianuÎncă nu există evaluări
- FEBRADocument63 paginiFEBRAIoana Muntianu100% (1)
- Grile Toxi VDocument3 paginiGrile Toxi VAlla HussÎncă nu există evaluări
- Arsuri Nou 3Document170 paginiArsuri Nou 3Ioana MuntianuÎncă nu există evaluări