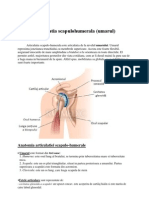
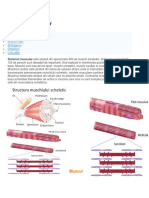
S-ar putea să vă placă și
- Sistemul Osos Sistemul Muscular La OmDocument8 paginiSistemul Osos Sistemul Muscular La OmannymarigÎncă nu există evaluări
- Clavicula Si OmoplatulDocument10 paginiClavicula Si OmoplatulAlina CamerzanÎncă nu există evaluări
- Artrologie, Articulatii, Tipuri de Miscari.Document37 paginiArtrologie, Articulatii, Tipuri de Miscari.Marina CarmenÎncă nu există evaluări
- Nervii SpinaliDocument3 paginiNervii SpinaliDanutz MariusÎncă nu există evaluări
- Oasele Carpiene PresentationDocument32 paginiOasele Carpiene PresentationBogdan Alexandru CristeiÎncă nu există evaluări
- Articulatii - LPDocument7 paginiArticulatii - LPclaudia_gheneaÎncă nu există evaluări
- Biobecanica ArticulatiilorDocument53 paginiBiobecanica ArticulatiilorionÎncă nu există evaluări
- Anatomie Si Biomecanica UmarDocument28 paginiAnatomie Si Biomecanica UmarHristodoulakis Stefanos0% (1)
- Muschi Si Locuri SlabeDocument23 paginiMuschi Si Locuri SlabeDaniela MoraruÎncă nu există evaluări
- Corpul CalosDocument1 paginăCorpul Calosauras27mateiÎncă nu există evaluări
- Articulatiile TrunchiuluiDocument2 paginiArticulatiile TrunchiuluiAdelina SimonÎncă nu există evaluări
- Coloana, Peretele ToracicDocument49 paginiColoana, Peretele ToracicIwannaÎncă nu există evaluări
- Kinesiologie 02Document8 paginiKinesiologie 02Contratov Lucian AndreiÎncă nu există evaluări
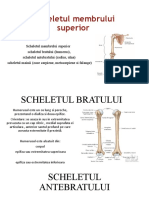
- Scheletul Membrului SuperiorDocument7 paginiScheletul Membrului SuperiorTanticaStoianÎncă nu există evaluări
- Muschii Membrelor SuperioareDocument5 paginiMuschii Membrelor SuperioarealexanadraÎncă nu există evaluări
- Curs 1. Osul Ca OrganDocument36 paginiCurs 1. Osul Ca OrganMolnár CsillaÎncă nu există evaluări
- Osteologia MembrelorDocument50 paginiOsteologia MembrelorandreiÎncă nu există evaluări
- Muschii PrezentareDocument12 paginiMuschii Prezentarerugina simona100% (1)
- Tabele EvaluareDocument12 paginiTabele EvaluareDuta Daniela100% (1)
- Extensia CoapseiDocument9 paginiExtensia CoapseiCarolina HîncuÎncă nu există evaluări
- Notiuni de Artrologie - Bugiuianu GabrielDocument9 paginiNotiuni de Artrologie - Bugiuianu GabrielBugiuianu Gabi100% (1)
- Proiect Facultate KinetoterapieDocument65 paginiProiect Facultate KinetoterapieCTS GiurgiuÎncă nu există evaluări
- BIOLOGIE Suport de CursDocument34 paginiBIOLOGIE Suport de CursIrina Florentina100% (1)
- Functiile Fundamentale Ale Organismului UmanDocument3 paginiFunctiile Fundamentale Ale Organismului UmanPocola100% (1)
- Muschii Extremitatii CefaliceDocument4 paginiMuschii Extremitatii CefalicemadalinaghÎncă nu există evaluări
- OsteologiaDocument9 paginiOsteologiaOleg RussuÎncă nu există evaluări
- Curs 3miologieDocument37 paginiCurs 3miologieVlaicu Tudor100% (1)
- MUSCHIIDocument24 paginiMUSCHIISandy BcÎncă nu există evaluări
- Adducţia Omoplatului: Muşchi Origine InserţieDocument3 paginiAdducţia Omoplatului: Muşchi Origine InserţieCramer IonelaÎncă nu există evaluări
- Muschi GatuluiDocument10 paginiMuschi GatuluiPansyMaryCreataÎncă nu există evaluări
- Dobre Amelia KINETOTERAPIA DEFICIENTELOR FIZICE SI SENZORIALEDocument4 paginiDobre Amelia KINETOTERAPIA DEFICIENTELOR FIZICE SI SENZORIALEAmelia F. DobreÎncă nu există evaluări
- Alcătuirea Generală A Corpului Uman - Sistemul NervosDocument32 paginiAlcătuirea Generală A Corpului Uman - Sistemul NervosEllyzaEllyÎncă nu există evaluări
- SOLDULDocument2 paginiSOLDULjane.dÎncă nu există evaluări
- Anatomie 2Document10 paginiAnatomie 2Kerekes CsengeÎncă nu există evaluări
- Anatomia Omului Articulatiile Prezentare InteractivaDocument25 paginiAnatomia Omului Articulatiile Prezentare InteractivaNeagu George100% (1)
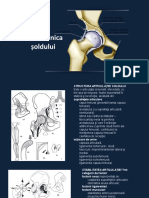
- Biomecanica SolduluiDocument47 paginiBiomecanica SolduluiCatalinaHadaragÎncă nu există evaluări
- Bisorca Oana-Cristina Articulatia CotuluiDocument12 paginiBisorca Oana-Cristina Articulatia CotuluiOana Cristina100% (1)
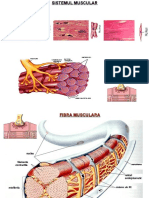
- Tesutul MuscularDocument8 paginiTesutul MusculardanÎncă nu există evaluări
- Generalitati Sist OsosDocument59 paginiGeneralitati Sist OsosNoémi ShepherdÎncă nu există evaluări
- Muschii AbdomenuluiDocument1 paginăMuschii AbdomenuluiAlexei RobuÎncă nu există evaluări
- Seminar 2 Tehnici de Manevrare A BolnavuluiDocument2 paginiSeminar 2 Tehnici de Manevrare A BolnavuluiJst DanezuÎncă nu există evaluări
- Coloana Lombara Curs 8Document6 paginiColoana Lombara Curs 8Gligan Alin-CristianÎncă nu există evaluări
- Oasele Membrelor Superioare Și Articulațiile Lor PDFDocument2 paginiOasele Membrelor Superioare Și Articulațiile Lor PDFGuritanu Alexandrina100% (1)
- Muschii Laterali Ai AntebratuluiDocument2 paginiMuschii Laterali Ai Antebratuluiblueberry03Încă nu există evaluări
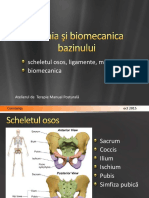
- Anatomia Si Biomecanica BazinuluiDocument21 paginiAnatomia Si Biomecanica BazinuluiAlexandru NechiforÎncă nu există evaluări
- Anatomia Sistemului MuscularDocument17 paginiAnatomia Sistemului MuscularNicoleta NastaseÎncă nu există evaluări
- Anatomie Curs 1 MuschiDocument2 paginiAnatomie Curs 1 MuschiRobert IonutÎncă nu există evaluări
- ARTICULAȚIIDocument5 paginiARTICULAȚIIatezorÎncă nu există evaluări
- Articulatiile Cutiei CranieneDocument5 paginiArticulatiile Cutiei CranienedomnfaneÎncă nu există evaluări
- ArtrologiaDocument24 paginiArtrologiaAlexandra IgnatÎncă nu există evaluări
- Miologie GeneralaDocument4 paginiMiologie GeneralaIoana Ruxandra AnghelÎncă nu există evaluări
- Articulatia ScapulohumeralaDocument3 paginiArticulatia ScapulohumeralaMaria GrigoreÎncă nu există evaluări
- Biomecanica Centurii ScapulareDocument4 paginiBiomecanica Centurii Scapulaready1266Încă nu există evaluări
- Vascularizatie Inervatie Anexe Mb. Sup.Document29 paginiVascularizatie Inervatie Anexe Mb. Sup.Maria NastaseÎncă nu există evaluări
- Curs AnatomieDocument109 paginiCurs AnatomieAlice Balean100% (1)
- Sistemul MuscularDocument30 paginiSistemul Muscularmidnightpoison1Încă nu există evaluări
- Celula Animală ContinuareDocument33 paginiCelula Animală ContinuareNicoleta CobanÎncă nu există evaluări
- CelulaDocument14 paginiCelulaFirma GSCÎncă nu există evaluări
- Fundamente BiologiceDocument17 paginiFundamente BiologiceElena MoiseÎncă nu există evaluări
- Organizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternDocument4 paginiOrganizarea Funcţională A Corpului Uman Şi Homeostazia Mediului InternVata LuminitaÎncă nu există evaluări
- Model de Cerere OUG 147Document2 paginiModel de Cerere OUG 147chivumihaelaÎncă nu există evaluări
- 26MODEL Declaratie Proprie Raspundere 2503Document1 pagină26MODEL Declaratie Proprie Raspundere 2503chivumihaelaÎncă nu există evaluări
- Modulul 2Document25 paginiModulul 2chivumihaelaÎncă nu există evaluări
- Cerere Concediu Fara PlataDocument1 paginăCerere Concediu Fara PlatachivumihaelaÎncă nu există evaluări
- Model Declarație Venituri (Expert Contabil)Document2 paginiModel Declarație Venituri (Expert Contabil)chivumihaelaÎncă nu există evaluări
- Model Declaratie OUG 147Document1 paginăModel Declaratie OUG 147Lasko AmelieÎncă nu există evaluări
- Orar C.P. G 2021-2022Document1 paginăOrar C.P. G 2021-2022chivumihaelaÎncă nu există evaluări
- Modulul 3 - RomanaDocument19 paginiModulul 3 - RomanachivumihaelaÎncă nu există evaluări
- Note de Curs - Masaj de Relaxare, Masaj AntistresDocument31 paginiNote de Curs - Masaj de Relaxare, Masaj Antistreschivumihaela50% (2)
- Curs 6Document23 paginiCurs 6chivumihaelaÎncă nu există evaluări
- Modul 4 Curs ManagementDocument15 paginiModul 4 Curs ManagementchivumihaelaÎncă nu există evaluări
- Note de Curs - Exerci - II Acvatice Pentru Pro Movarea Starii de SanatateDocument15 paginiNote de Curs - Exerci - II Acvatice Pentru Pro Movarea Starii de SanatatechivumihaelaÎncă nu există evaluări
- Note de Curs Aplicatii Ale Hidrokinetoterapiei in PediatrieDocument77 paginiNote de Curs Aplicatii Ale Hidrokinetoterapiei in PediatriechivumihaelaÎncă nu există evaluări
- Curs 5Document20 paginiCurs 5chivumihaelaÎncă nu există evaluări
- ElectroterapiaDocument50 paginiElectroterapiachivumihaelaÎncă nu există evaluări
- Curs AnatomieDocument33 paginiCurs AnatomiechivumihaelaÎncă nu există evaluări
- Curs Hidrokineto PitestiDocument1 paginăCurs Hidrokineto PitestichivumihaelaÎncă nu există evaluări
- Curs 3Document21 paginiCurs 3chivumihaelaÎncă nu există evaluări
- Note de Curs Bazele Masajului in Statiuni BCTDocument45 paginiNote de Curs Bazele Masajului in Statiuni BCTchivumihaelaÎncă nu există evaluări
- Curs 6Document23 paginiCurs 6chivumihaelaÎncă nu există evaluări
- Curs 2Document11 paginiCurs 2chivumihaelaÎncă nu există evaluări
- Curs 1Document12 paginiCurs 1chivumihaelaÎncă nu există evaluări
- Curs 4Document16 paginiCurs 4chivumihaelaÎncă nu există evaluări
- Note de Curs Metode Si Tehnici de Body Wellnes Si SPADocument68 paginiNote de Curs Metode Si Tehnici de Body Wellnes Si SPAchivumihaelaÎncă nu există evaluări
- Note de Curs Si LP Prim AjutorDocument41 paginiNote de Curs Si LP Prim AjutorchivumihaelaÎncă nu există evaluări
- Notiuni de Prim AjutorDocument57 paginiNotiuni de Prim AjutorchivumihaelaÎncă nu există evaluări
- Note de Curs Masaj SomaticDocument35 paginiNote de Curs Masaj SomaticchivumihaelaÎncă nu există evaluări
- Note de Curs Metode Si Tehnici de Body Wellnes Si SPADocument68 paginiNote de Curs Metode Si Tehnici de Body Wellnes Si SPAchivumihaelaÎncă nu există evaluări
- Note de Curs Bazele Teoretice Ale MasajuluiDocument29 paginiNote de Curs Bazele Teoretice Ale MasajuluichivumihaelaÎncă nu există evaluări
- Ordinul MDRT NR 1051-03 03 2011 - Norme MetodologiceDocument93 paginiOrdinul MDRT NR 1051-03 03 2011 - Norme MetodologiceSorina GeorgianaÎncă nu există evaluări