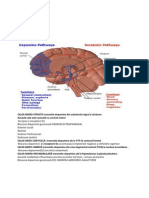
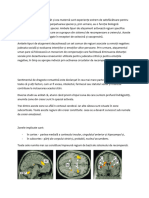
S-ar putea să vă placă și
- S04eu. F - Bazele Biologice Ale Psihiatriei, Neurotransmitatorii, Neuroanatomie FinalDocument12 paginiS04eu. F - Bazele Biologice Ale Psihiatriei, Neurotransmitatorii, Neuroanatomie FinalDiana Elena100% (1)
- Calea NigroDocument14 paginiCalea Nigrodrimaedi9513Încă nu există evaluări
- S04 F - Bazele Biologice Ale Psihiatriei, Neurotransmitatorii, Neuroanatomie FinalDocument12 paginiS04 F - Bazele Biologice Ale Psihiatriei, Neurotransmitatorii, Neuroanatomie FinalMakkai Popa Mihai VladimirÎncă nu există evaluări
- NeurotransmitatoriDocument3 paginiNeurotransmitatorigia_me20003Încă nu există evaluări
- Boala Parkinson Si Receptorii DOPADocument14 paginiBoala Parkinson Si Receptorii DOPATudor Daniel Cucos100% (3)
- Hormonii FericiriiDocument16 paginiHormonii FericiriiIonidia DilimoţÎncă nu există evaluări
- Dopamina WordDocument11 paginiDopamina WordRaluca IvanÎncă nu există evaluări
- NEUROPSIHOLOGIEDocument24 paginiNEUROPSIHOLOGIEGeorgiana CiucăÎncă nu există evaluări
- Ipoteze Neurobiologice Implicate in SchizofrenieDocument13 paginiIpoteze Neurobiologice Implicate in Schizofreniedianacirjaliu100% (1)
- Chimia Creierului 1Document3 paginiChimia Creierului 1Tasy BvÎncă nu există evaluări
- Curs 11 - AntiparkinsonieneleDocument5 paginiCurs 11 - AntiparkinsonieneleCristian RusuÎncă nu există evaluări
- AntiparkinsonieneDocument5 paginiAntiparkinsonieneXDCVXÎncă nu există evaluări
- Antiparkinsoniene An IIIDocument4 paginiAntiparkinsoniene An IIIPintilieNicoletaÎncă nu există evaluări
- S04 - Bazele Biologice Ale Psihiatriei, Neurotransmitatorii, NeuroanatomieDocument10 paginiS04 - Bazele Biologice Ale Psihiatriei, Neurotransmitatorii, NeuroanatomieMCÎncă nu există evaluări
- Rolul Dopaminei in Crearea Dependentei de Cocaina Si AmfetaminaDocument8 paginiRolul Dopaminei in Crearea Dependentei de Cocaina Si AmfetaminamaiasauleaÎncă nu există evaluări
- Farmacologie - AntidepresiveleDocument20 paginiFarmacologie - Antidepresiveledandda9393Încă nu există evaluări
- 4.rolul Dopaminei in ComportamentDocument1 pagină4.rolul Dopaminei in ComportamentDeea NiculaeÎncă nu există evaluări
- Dopamina 2Document5 paginiDopamina 2nitu roxanaÎncă nu există evaluări
- Neuro Curs 3 Psihologie FacultateDocument5 paginiNeuro Curs 3 Psihologie FacultateMaiden DiÎncă nu există evaluări
- Neurotransmitatorii Implicati in Depresie Si AnxietateDocument6 paginiNeurotransmitatorii Implicati in Depresie Si AnxietateAlina MaselutaÎncă nu există evaluări
- AdictiiDocument71 paginiAdictiiAnda Madalina Zaharia100% (1)
- PSIHONEUROFIZIOLOGIEDocument7 paginiPSIHONEUROFIZIOLOGIEmihai2lungÎncă nu există evaluări
- Serotonina Sau 5Document5 paginiSerotonina Sau 5Ioana FlorinaÎncă nu există evaluări
- NeurotransmitatoriDocument4 paginiNeurotransmitatoriAnaXor100% (1)
- Neurotransmitatori ScuDocument3 paginiNeurotransmitatori ScuweikunÎncă nu există evaluări
- 04bazele Biol Ale Psih - Neurotransmitatorii, Neuroanatomie FunctionalaDocument10 pagini04bazele Biol Ale Psih - Neurotransmitatorii, Neuroanatomie FunctionalarrrrraluÎncă nu există evaluări
- 5-A-Luca Amalia Gabriela-Boala Parkinson Și Receptorii DOPADocument10 pagini5-A-Luca Amalia Gabriela-Boala Parkinson Și Receptorii DOPAAmyLucaÎncă nu există evaluări
- NeurolepticeDocument9 paginiNeurolepticeSpiridon Madalina100% (1)
- Clasificarea Neurotransmitatorilor UpdateDocument7 paginiClasificarea Neurotransmitatorilor UpdateNadia DragomirÎncă nu există evaluări
- Neurotransmitori Si NeuromodulatoriDocument6 paginiNeurotransmitori Si NeuromodulatoriMadalina Ioana BratuÎncă nu există evaluări
- Curs 5 - Sistemul Ganglionilor Bazali (Continuare)Document7 paginiCurs 5 - Sistemul Ganglionilor Bazali (Continuare)Laurentiu AndreiÎncă nu există evaluări
- Dopamina Si Dependenta de DroguriDocument6 paginiDopamina Si Dependenta de DroguriSabina DiacÎncă nu există evaluări
- Neuroanatomia Si Neurofiziologia Comportamentului.Document3 paginiNeuroanatomia Si Neurofiziologia Comportamentului.Luiza BezneaÎncă nu există evaluări
- Dependenta Incepe in Creier IFDocument11 paginiDependenta Incepe in Creier IFadelaÎncă nu există evaluări
- OTILIA MITITELU - Relatia Dintre Mecanismele Nervoase Si Cele HormonaleDocument11 paginiOTILIA MITITELU - Relatia Dintre Mecanismele Nervoase Si Cele HormonaleElena LupuÎncă nu există evaluări
- NeurotransmitatoriiDocument20 paginiNeurotransmitatoriipentrucanva1Încă nu există evaluări
- NEUROTRANSMIŢĂTORII Curs 4Document14 paginiNEUROTRANSMIŢĂTORII Curs 4vasilica888100% (3)
- Neurobiologia IubiriiDocument3 paginiNeurobiologia IubiriiSorin PantelimonÎncă nu există evaluări
- Psihofarmacologie Ptr. TCCDocument36 paginiPsihofarmacologie Ptr. TCCMălina-Ionela Corlătianu100% (1)
- Substanțe Toxice Cu Acțiune DislepticăDocument28 paginiSubstanțe Toxice Cu Acțiune DislepticăCorina PopescuÎncă nu există evaluări
- Neurobiologia DepresieiDocument3 paginiNeurobiologia DepresieiDodoaxl AndreeaÎncă nu există evaluări
- Adrenalina Si NoradrenalinaDocument5 paginiAdrenalina Si NoradrenalinaZuZuÎncă nu există evaluări
- Transmisia Dopaminergică 2Document18 paginiTransmisia Dopaminergică 2Daniela GligorÎncă nu există evaluări
- Efectele Consumului de Amfetamine Si BenzodiazepineDocument13 paginiEfectele Consumului de Amfetamine Si BenzodiazepineMarinel Belciug100% (1)
- Particularitati Clinice Si Psihofarmacologice Ale Tratamentului Cu Antispihotice in SchizofrenieDocument40 paginiParticularitati Clinice Si Psihofarmacologice Ale Tratamentului Cu Antispihotice in SchizofrenieAlex VimanÎncă nu există evaluări
- MegaDocument603 paginiMegaAndra Alina NeculițăÎncă nu există evaluări
- SerotoninaDocument2 paginiSerotoninaDaniela VladÎncă nu există evaluări
- AdictiaDocument8 paginiAdictiaAlina FloreaÎncă nu există evaluări
- NeurotransmitatoriDocument8 paginiNeurotransmitatoriciolanescuÎncă nu există evaluări
- Psihotrope IIDocument60 paginiPsihotrope IIMaximciuc Corneliu100% (1)
- CURSUL II-Bazele Biologice Ale Comportamentului IIDocument18 paginiCURSUL II-Bazele Biologice Ale Comportamentului IIannamaria balintÎncă nu există evaluări
- Sindromul ExtrapiramdalDocument6 paginiSindromul ExtrapiramdalAndreea GÎncă nu există evaluări
- SchizofrenieDocument9 paginiSchizofrenieDaniela GreereÎncă nu există evaluări
- NeurolepticeDocument9 paginiNeurolepticeElena ChitoiuÎncă nu există evaluări
- Economia de dopamină: Cum ne influențează neurotransmițătorul plăcerii deciziile și comportamentul nostru în viața de zi cu ziDe la EverandEconomia de dopamină: Cum ne influențează neurotransmițătorul plăcerii deciziile și comportamentul nostru în viața de zi cu ziÎncă nu există evaluări
- PSICOLOGIE, DEPRESIUNE ȘI DISTURBARE UMORALĂ: înțelegerea mecanismelor de bazăDe la EverandPSICOLOGIE, DEPRESIUNE ȘI DISTURBARE UMORALĂ: înțelegerea mecanismelor de bazăEvaluare: 5 din 5 stele5/5 (1)
- Abordarea minte-corp pentru tratarea depresiei: Cum să folosiți meditația, yoga și ayurveda pentru a vă îmbunătăți fericireaDe la EverandAbordarea minte-corp pentru tratarea depresiei: Cum să folosiți meditația, yoga și ayurveda pentru a vă îmbunătăți fericireaEvaluare: 4 din 5 stele4/5 (4)
- Abordari psihodinamice ale schimbarii comportamentaleDe la EverandAbordari psihodinamice ale schimbarii comportamentaleÎncă nu există evaluări
- Past Perfect and ContinuousDocument3 paginiPast Perfect and ContinuousIonita LauraÎncă nu există evaluări
- Depresia Postpartum - Test ProfesionalDocument3 paginiDepresia Postpartum - Test ProfesionalIonita LauraÎncă nu există evaluări
- ASQ AnxietateDocument2 paginiASQ AnxietateIonita LauraÎncă nu există evaluări
- Chestiionarul de Diagnostic E2 RimlandDocument12 paginiChestiionarul de Diagnostic E2 RimlandIonita Laura100% (2)
- Calculul QI - Populaţie 35-60aniDocument1 paginăCalculul QI - Populaţie 35-60aniIonita LauraÎncă nu există evaluări
- CHESTIONAR DE EVALUARE A INTERESELOR CognitromDocument5 paginiCHESTIONAR DE EVALUARE A INTERESELOR CognitromIonita LauraÎncă nu există evaluări
- Anxietatea SocialăDocument4 paginiAnxietatea SocialăIonita LauraÎncă nu există evaluări
- Lecția 7 - Adjectivul - The AdjectiveDocument7 paginiLecția 7 - Adjectivul - The AdjectiveIonita LauraÎncă nu există evaluări
- Present Perfect Simple - Prezentul Perfect SimpluDocument11 paginiPresent Perfect Simple - Prezentul Perfect SimpluIonita LauraÎncă nu există evaluări
- EnglezaDocument204 paginiEnglezaMihaela VadanÎncă nu există evaluări
- Conditionalul If Lecția 10Document4 paginiConditionalul If Lecția 10Ionita LauraÎncă nu există evaluări
- Adrenalina ReferatDocument6 paginiAdrenalina ReferatIonita Laura100% (2)
- Lecția 1Document20 paginiLecția 1Ionita LauraÎncă nu există evaluări
- Personalitatile Dificile II-1Document52 paginiPersonalitatile Dificile II-1Irina CebucÎncă nu există evaluări
- Tulburarea de Personalitate HistrionicaDocument4 paginiTulburarea de Personalitate HistrionicaIonita LauraÎncă nu există evaluări
- Proiect de Seminar Disfunctii Cognitive Si de LimbajDocument5 paginiProiect de Seminar Disfunctii Cognitive Si de LimbajIonita LauraÎncă nu există evaluări
- Tulburarea AspergerDocument3 paginiTulburarea AspergerIonita LauraÎncă nu există evaluări
- "Prevenţie Şi Tratament Cu Ajutorul Terapiilor de Relaxare".Document5 pagini"Prevenţie Şi Tratament Cu Ajutorul Terapiilor de Relaxare".Ionita LauraÎncă nu există evaluări
- Proiect GerontopsihiatrieDocument13 paginiProiect GerontopsihiatrieIonita LauraÎncă nu există evaluări
- Ioniţă Laura CătălinaDocument12 paginiIoniţă Laura CătălinaIonita LauraÎncă nu există evaluări
- Proiect Laura IrimiaDocument7 paginiProiect Laura IrimiaIonita LauraÎncă nu există evaluări
- ADHD Tratament Și IntervențieDocument16 paginiADHD Tratament Și IntervențieRizon IoanaÎncă nu există evaluări
- Psihoterapie Rational Emotiva Si Comportamentala Pentru Depresie PDFDocument89 paginiPsihoterapie Rational Emotiva Si Comportamentala Pentru Depresie PDFjeni chiriac100% (10)
- Psihoterapie NarcisaDocument4 paginiPsihoterapie NarcisaIonita LauraÎncă nu există evaluări
- Workshop Inteligenta SocialaDocument7 paginiWorkshop Inteligenta SocialaIonita LauraÎncă nu există evaluări
- Proiect - Consiliere Si Orientare Scolara Si ProfesionalaDocument5 paginiProiect - Consiliere Si Orientare Scolara Si ProfesionalaIonita LauraÎncă nu există evaluări
- Bazele Neurobiologice Si Psihofarmacologice in DepresieDocument7 paginiBazele Neurobiologice Si Psihofarmacologice in DepresieIonita LauraÎncă nu există evaluări
- Proiect - Psihodiagnoza AptitudinilorDocument6 paginiProiect - Psihodiagnoza AptitudinilorIonita LauraÎncă nu există evaluări
- Caracterizare PsihopedagogicaDocument1 paginăCaracterizare PsihopedagogicaIonita LauraÎncă nu există evaluări
- Proiect Psihologie OrganizationalaDocument7 paginiProiect Psihologie OrganizationalaIonita LauraÎncă nu există evaluări