S-ar putea să vă placă și
- Cromozomii 22Document18 paginiCromozomii 22Ama PredaÎncă nu există evaluări
- 8 - Genetica DezvoltariiDocument6 pagini8 - Genetica DezvoltariiVlad PerpeleaÎncă nu există evaluări
- LP MeiozaDocument14 paginiLP MeiozaSimionescu AlinÎncă nu există evaluări
- Histologie Curs 1) Tesuturi EpitelialeDocument6 paginiHistologie Curs 1) Tesuturi EpitelialeMariaMaryaÎncă nu există evaluări
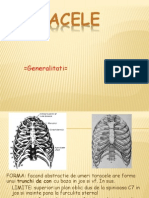
- Toracele - Anatomie Regionala Si TopograficaDocument14 paginiToracele - Anatomie Regionala Si TopograficaMara ChiricaÎncă nu există evaluări
- Teste Tot 2Document8 paginiTeste Tot 2Ana BunescuÎncă nu există evaluări
- Nomenclatura MutatiiDocument31 paginiNomenclatura MutatiiMerlan IulianÎncă nu există evaluări
- Totalizarea 2Document43 paginiTotalizarea 2Bunici MarinelaÎncă nu există evaluări
- Vitaminele La CopiiDocument15 paginiVitaminele La CopiiMalvina GhalayiniÎncă nu există evaluări
- Reglarea Expresiei GenelorDocument13 paginiReglarea Expresiei GenelorDanciu LoredanaÎncă nu există evaluări
- Teste Tot 1 2016Document16 paginiTeste Tot 1 2016Oleg BalanÎncă nu există evaluări
- An Ii LP6Document40 paginiAn Ii LP6mirelaÎncă nu există evaluări
- Diviziunea CelularăDocument7 paginiDiviziunea CelularăGeorge BrinzeaÎncă nu există evaluări
- Schema ProtozoareDocument2 paginiSchema ProtozoareAlexandra ZarzuÎncă nu există evaluări
- Diagnosticul de Laborator in AmibiazaDocument3 paginiDiagnosticul de Laborator in AmibiazaSimonaÎncă nu există evaluări
- ROM 10. BacteriofagiiDocument42 paginiROM 10. BacteriofagiiGabi SaharneanÎncă nu există evaluări
- Anemiile MacrocitareDocument7 paginiAnemiile Macrocitaretzorella_89Încă nu există evaluări
- 25.sistemul ExcitoconductorDocument1 pagină25.sistemul ExcitoconductorLumyÎncă nu există evaluări
- Indicatii - Metodice - Medicamente - Cu - Influenta - Asupra - Singelui - Si - Organelor - Hematopoetice (1) - 1588597664Document8 paginiIndicatii - Metodice - Medicamente - Cu - Influenta - Asupra - Singelui - Si - Organelor - Hematopoetice (1) - 1588597664MariaÎncă nu există evaluări
- Capitol 9 Transcriptia ROMDocument10 paginiCapitol 9 Transcriptia ROMAndrei Rusu100% (1)
- teste la fiziologie.conspecte.md копия4333301357759539466Document47 paginiteste la fiziologie.conspecte.md копия4333301357759539466Irina RaileanuÎncă nu există evaluări
- Determinarea Grupei Sangvine - Vasilica AriadnaDocument2 paginiDeterminarea Grupei Sangvine - Vasilica AriadnaArina VasilicaÎncă nu există evaluări
- Subiecte Fizio LPDocument27 paginiSubiecte Fizio LPhubertsÎncă nu există evaluări
- Sub Rez Sem2practic FizioDocument39 paginiSub Rez Sem2practic FizioGabrielPaduretÎncă nu există evaluări
- Cultivarea VirusurilorDocument13 paginiCultivarea VirusurilorMihaela VladutÎncă nu există evaluări
- Biochimie Amg IDocument3 paginiBiochimie Amg IClaudiu GateaÎncă nu există evaluări
- Lucrarea Practica Nr. 3 Darii Corina M1630Document25 paginiLucrarea Practica Nr. 3 Darii Corina M1630Cătălin Condrea100% (1)
- Patologie Chirurgicala ToraceDocument10 paginiPatologie Chirurgicala ToraceSebastian Stoica100% (1)
- DistrofiileDocument187 paginiDistrofiileGheorghe BiancaÎncă nu există evaluări
- Histologie Curs 05Document9 paginiHistologie Curs 05Carmen-Elena ManeaÎncă nu există evaluări
- Ciclul CelularDocument8 paginiCiclul CelularAna TuchilaÎncă nu există evaluări
- TraheostomiaDocument2 paginiTraheostomiaMorcov Andreea-NicoletaÎncă nu există evaluări
- Curs 8 FiziologieDocument7 paginiCurs 8 FiziologieAlexa MacoveiÎncă nu există evaluări
- FiziologieDocument18 paginiFiziologieGicu Butuc100% (1)
- Valenţa Antigenului S-A Definit Convenţional Prin Numărul de Epitopi Ai Unui AntigenDocument1 paginăValenţa Antigenului S-A Definit Convenţional Prin Numărul de Epitopi Ai Unui AntigenDana CopoiuÎncă nu există evaluări
- Colectarea de DateDocument1 paginăColectarea de DateCristina LupascoÎncă nu există evaluări
- Fiziologie Animala Generala - Suport de CursDocument66 paginiFiziologie Animala Generala - Suport de CursvarcolacÎncă nu există evaluări
- Generarea Si Regenerarea A Fibrelor NervoaseDocument11 paginiGenerarea Si Regenerarea A Fibrelor NervoaseNastiu6a100% (1)
- LP 1 - Microscopie Optica +Document30 paginiLP 1 - Microscopie Optica +Daniela DiaconescuÎncă nu există evaluări
- Totalizare NR 3Document14 paginiTotalizare NR 3Mihaela CecanÎncă nu există evaluări
- Arbovirusuri 222CURSDocument73 paginiArbovirusuri 222CURSMaria MirabelaÎncă nu există evaluări
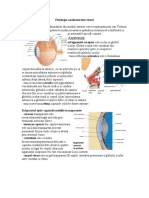
- Fiziologia Analizatorului VizualDocument2 paginiFiziologia Analizatorului VizualPopa CarmenÎncă nu există evaluări
- Curs 01 - 01 - Curs Tesut Epitelial1Document75 paginiCurs 01 - 01 - Curs Tesut Epitelial1Bubulac AngelaÎncă nu există evaluări
- Biofizica PT EleviDocument91 paginiBiofizica PT EleviCojocaru ConstantinÎncă nu există evaluări
- Fiziologie - Curs 5 Neuronul + MiocardulDocument8 paginiFiziologie - Curs 5 Neuronul + Miocardulionut_lupul100% (1)
- Aparatul Genetic Al Celulelor Umane Este Format Din Structuri Celulare Ce Conţin ADNDocument1 paginăAparatul Genetic Al Celulelor Umane Este Format Din Structuri Celulare Ce Conţin ADNDaniela DedovÎncă nu există evaluări
- Toate TesteleDocument108 paginiToate TesteleTimon AgarÎncă nu există evaluări
- 02 - Amiloidoza - Hialinoza - Pigmentațiile-EndogeneDocument12 pagini02 - Amiloidoza - Hialinoza - Pigmentațiile-Endogenemarinacretu1Încă nu există evaluări
- Curs 5 Medicina Dezv UmaneDocument75 paginiCurs 5 Medicina Dezv Umanemiruna_cÎncă nu există evaluări
- Curs 3Document23 paginiCurs 3avmd_roÎncă nu există evaluări
- Tesuturi Excitabile II SMLDocument3 paginiTesuturi Excitabile II SMLMihai TÎncă nu există evaluări
- Informatii Generale GeneticaDocument22 paginiInformatii Generale GeneticaAnonymous ck0aGAqZlÎncă nu există evaluări
- ABERATII CROMOZOMIALE - PTDocument29 paginiABERATII CROMOZOMIALE - PTAndreea LorelaiÎncă nu există evaluări
- Genotipul Uman PatologicDocument8 paginiGenotipul Uman PatologicRuxi CiaglicÎncă nu există evaluări
- Genetică Curs 6 13Document38 paginiGenetică Curs 6 13Andreea CătălinaÎncă nu există evaluări
- Curs Genetica 6,7,8,9,10,11,12,13Document38 paginiCurs Genetica 6,7,8,9,10,11,12,13Maria JalbaÎncă nu există evaluări
- Referat GeneticaDocument6 paginiReferat GeneticaEmilian CretuÎncă nu există evaluări
- LP 5 Studiul Aberatiilor CromozomialeDocument5 paginiLP 5 Studiul Aberatiilor CromozomialeDamian CojocaruÎncă nu există evaluări
- 7.5. Proiect Didactic (Sablon)Document6 pagini7.5. Proiect Didactic (Sablon)Valrtina CalinÎncă nu există evaluări
- Aberatii Cromozomiale - PTDocument29 paginiAberatii Cromozomiale - PTDragos AlexandruÎncă nu există evaluări