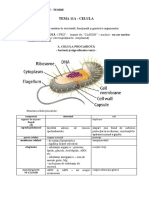
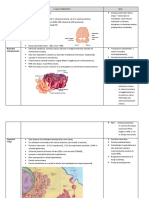
S-ar putea să vă placă și
- Diversitatea Lumii ViiDocument4 paginiDiversitatea Lumii ViiLavinia MihaelaÎncă nu există evaluări
- Material Olimpiada Cls X - Histologie Si HranireDocument61 paginiMaterial Olimpiada Cls X - Histologie Si HranireIrina Borotea100% (2)
- Celula Procariota Si EucariotaDocument4 paginiCelula Procariota Si Eucariotacapota100% (2)
- Morfologia Plantelor - Botanica Semestrul 1Document12 paginiMorfologia Plantelor - Botanica Semestrul 1David StoianÎncă nu există evaluări
- Abateri de La SegregareDocument2 paginiAbateri de La SegregarePopescu ElenaÎncă nu există evaluări
- Tesuturi VegetaleDocument10 paginiTesuturi VegetaleAncuța Rãileanu100% (2)
- Biologie Clasa A 10 ADocument18 paginiBiologie Clasa A 10 ACarmenModoranÎncă nu există evaluări
- Organizarea Materialului Genetic La Virusuri Si ProcarioteDocument19 paginiOrganizarea Materialului Genetic La Virusuri Si ProcarioteAlex Cioanca100% (1)
- Sistem Circulator Pesti - DescriereDocument2 paginiSistem Circulator Pesti - DescriereAndreea ArseneÎncă nu există evaluări
- Referat Tesuturi VegetaleDocument2 paginiReferat Tesuturi VegetaleBianca-Ioana Popescu0% (1)
- LCRDocument46 paginiLCRSpinu Lilia100% (1)
- MeristemeDocument4 paginiMeristemeMarcelaMorosanu0% (1)
- Țesuturi VegetaleDocument4 paginiȚesuturi VegetaleLuminita LuminitaÎncă nu există evaluări
- Ţesuturi Vegetale Şi AnimaleDocument15 paginiŢesuturi Vegetale Şi AnimaleDana Pluhovici100% (2)
- Tesuturile VegetaleDocument1 paginăTesuturile VegetaleAlex MaxÎncă nu există evaluări
- Tesuturi VegetaleDocument1 paginăTesuturi VegetaleVeronica MotocÎncă nu există evaluări
- Celula Diviziunea CelularaDocument26 paginiCelula Diviziunea CelularaGabØrÎncă nu există evaluări
- Biologie TesuturiDocument5 paginiBiologie TesuturiBogdyÎncă nu există evaluări
- Ţesuturi VegetaleDocument20 paginiŢesuturi VegetaleOvidiu CîmpanÎncă nu există evaluări
- Influenta Mediului Asupra EreditatiiDocument2 paginiInfluenta Mediului Asupra EreditatiiBunea Ispas Aurora100% (2)
- Exercitii TesuturiDocument3 paginiExercitii TesuturiXenna MiÎncă nu există evaluări
- Tesuturi Meristematice Si DefinitiveDocument26 paginiTesuturi Meristematice Si Definitivecrisene63Încă nu există evaluări
- Test Recapitulativ Regnul PlanteDocument2 paginiTest Recapitulativ Regnul PlantenadeaÎncă nu există evaluări
- Constituienți ProtoplasmaticiDocument5 paginiConstituienți ProtoplasmaticiAlex TurcÎncă nu există evaluări
- Subiecte Oli Loc 2019 IxDocument11 paginiSubiecte Oli Loc 2019 IxStefan GhergheÎncă nu există evaluări
- Ţesuturi VegetaleDocument4 paginiŢesuturi VegetaleTimmyRso100% (3)
- LenticeleDocument3 paginiLenticeleCris1234450% (2)
- Variante DihibridareDocument4 paginiVariante Dihibridareconstantin_razvan100% (1)
- Bac Bio 9-10Document78 paginiBac Bio 9-10Denisa Badoi100% (1)
- Diviziunea Celulara A Celulei Normale EucarioteDocument3 paginiDiviziunea Celulara A Celulei Normale EucarioteSirb AlexandruÎncă nu există evaluări
- Ţesuturi VegetaleDocument4 paginiŢesuturi VegetaleEusebiu Tarca100% (1)
- Structura Si Ultra Structura CeluleiDocument7 paginiStructura Si Ultra Structura CeluleiLucas Sandor100% (2)
- Test Celula Procariota Si EucariotaDocument2 paginiTest Celula Procariota Si EucariotaElena Stan100% (1)
- Curs 5Document7 paginiCurs 5marissaxz100% (1)
- Variante Legile Mendeliene Ale Ereditatii 2hDocument3 paginiVariante Legile Mendeliene Ale Ereditatii 2hCorban MihaiÎncă nu există evaluări
- 2004 Biologie Locala Subiecte Clasa A X-ADocument10 pagini2004 Biologie Locala Subiecte Clasa A X-ACristina PeleaÎncă nu există evaluări
- Organite CelulareDocument6 paginiOrganite CelulareBobo bobo100% (1)
- Test de Evaluare A 9aDocument2 paginiTest de Evaluare A 9aAlexandre Martin PineÎncă nu există evaluări
- Abaterea de La Segregarea MendelianaDocument1 paginăAbaterea de La Segregarea MendelianaBianca OlteanuÎncă nu există evaluări
- 2013 Biologie Locala Bucuresti Clasa A Xa Subiecte Si BaremeDocument12 pagini2013 Biologie Locala Bucuresti Clasa A Xa Subiecte Si BaremeChiritoiu AndreiÎncă nu există evaluări
- Comparatie Celula Ek Si PKDocument8 paginiComparatie Celula Ek Si PKVoyageR_IKÎncă nu există evaluări
- Celula - Lecție Completă.Document11 paginiCelula - Lecție Completă.Ștefan Lefter100% (1)
- Celula EucariotaDocument13 paginiCelula EucariotaDenisa MariaÎncă nu există evaluări
- Teste BiologieDocument25 paginiTeste BiologieWilly LucyÎncă nu există evaluări
- SINTEZE ȘI TESTE BAC - Clasa A IX-aDocument16 paginiSINTEZE ȘI TESTE BAC - Clasa A IX-alazarÎncă nu există evaluări
- Genetica Umana Clasa A IX-aDocument5 paginiGenetica Umana Clasa A IX-atudoracheconstantaÎncă nu există evaluări
- Histologie Vegetală - Test Grilă Var. 2019Document3 paginiHistologie Vegetală - Test Grilă Var. 2019anca_gheorma100% (1)
- 2009 Biologie Etapa Judeteana Subiecte Clasa A IX-A 0Document11 pagini2009 Biologie Etapa Judeteana Subiecte Clasa A IX-A 0Godja LydiaÎncă nu există evaluări
- Test TesuturiDocument3 paginiTest TesuturiElena Eu Eu ElenaÎncă nu există evaluări
- Caracterizarea Radacinii Si TulpiniiDocument8 paginiCaracterizarea Radacinii Si TulpiniiBURCEA MARIANAÎncă nu există evaluări
- Subiecte Proba Practica IxDocument7 paginiSubiecte Proba Practica IxLily LilonÎncă nu există evaluări
- PlastideDocument11 paginiPlastideRizeaDianaÎncă nu există evaluări
- Lecția - Gimnosperme, Clasa A 9-ADocument4 paginiLecția - Gimnosperme, Clasa A 9-ANicoleta Ginghina100% (1)
- BioDocument23 paginiBioSilvia Silviutza100% (1)
- Circulaţia La PlanteDocument17 paginiCirculaţia La Plantedianamihaela30Încă nu există evaluări
- Tesuturi Vegetale SimpleDocument24 paginiTesuturi Vegetale SimpleAnnaÎncă nu există evaluări
- HistologieDocument19 paginiHistologieAlexa CărpuşorÎncă nu există evaluări
- Botanica Curs 4Document86 paginiBotanica Curs 4Georgiana SimionÎncă nu există evaluări
- Ţesuturi VegetaleDocument265 paginiŢesuturi VegetaleTimothy Benson100% (4)
- Histologie Vegetală UsamvDocument10 paginiHistologie Vegetală UsamvAdi Postelnicu100% (1)
- Tesuturi CLSF, Origine, DefinDocument36 paginiTesuturi CLSF, Origine, DefinAlexandra CismasÎncă nu există evaluări
- Anatomia Omului - CelulaDocument4 paginiAnatomia Omului - CelulaSpinu LiliaÎncă nu există evaluări
- PARAZITOZEDocument67 paginiPARAZITOZESpinu Lilia100% (2)
- Reptile 2Document2 paginiReptile 2Spinu LiliaÎncă nu există evaluări
- PasariDocument3 paginiPasariSpinu LiliaÎncă nu există evaluări
- PREZENTARE ZoologieDocument19 paginiPREZENTARE ZoologieSpinu LiliaÎncă nu există evaluări
- 7patol Pigmenţi Biliari Compet 2018Document21 pagini7patol Pigmenţi Biliari Compet 2018Spinu LiliaÎncă nu există evaluări
- Proteinele Plasmatice 2020Document18 paginiProteinele Plasmatice 2020Spinu LiliaÎncă nu există evaluări
- AmfibieniDocument2 paginiAmfibieniSpinu LiliaÎncă nu există evaluări
- Referat EnzimologieDocument18 paginiReferat EnzimologieSpinu LiliaÎncă nu există evaluări
- Enzime GeneralitatiDocument60 paginiEnzime GeneralitatiSpinu LiliaÎncă nu există evaluări
- 3 Etapele Unui Proces BiotehnologicDocument2 pagini3 Etapele Unui Proces BiotehnologicSpinu LiliaÎncă nu există evaluări
- Proprietatatile UrineiDocument37 paginiProprietatatile UrineiSpinu LiliaÎncă nu există evaluări
- Enzimele PlasmaticeDocument33 paginiEnzimele PlasmaticeSpinu LiliaÎncă nu există evaluări
- 3chimia, Patochimia ProteinelorDocument61 pagini3chimia, Patochimia ProteinelorSpinu LiliaÎncă nu există evaluări
- Secret VaginalDocument41 paginiSecret VaginalSpinu LiliaÎncă nu există evaluări
- Microorganisme Utilizate În BiotehnologiiDocument11 paginiMicroorganisme Utilizate În BiotehnologiiSpinu LiliaÎncă nu există evaluări
- Laborator Chimie CULEGERE FinalDocument65 paginiLaborator Chimie CULEGERE FinalSpinu LiliaÎncă nu există evaluări
- Tesutul Muscular PDFDocument26 paginiTesutul Muscular PDFSpinu LiliaÎncă nu există evaluări