S-ar putea să vă placă și
- Analizatorul Somatosenzorial-25327Document107 paginiAnalizatorul Somatosenzorial-25327Cătălina HadjiogloÎncă nu există evaluări
- C8 - SomesteziaDocument26 paginiC8 - Somesteziablacklovex100% (1)
- Tema 13 C13, LP13 PDFDocument13 paginiTema 13 C13, LP13 PDFVioleta Rozalia HosuÎncă nu există evaluări
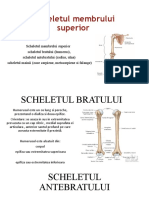
- Scheletul Membrului SuperiorDocument7 paginiScheletul Membrului SuperiorTanticaStoianÎncă nu există evaluări
- CEREBELDocument48 paginiCEREBELRaluca Cernat100% (1)
- PERICARDULDocument2 paginiPERICARDULAdriana IuliaÎncă nu există evaluări
- Vase Membru SuperiorDocument52 paginiVase Membru SuperiorOana Nedelcu100% (1)
- Curs Col Cervicala 2Document42 paginiCurs Col Cervicala 2Raluca DausÎncă nu există evaluări
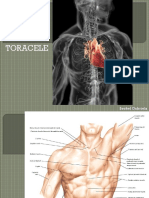
- OSTEOTORACELEDocument45 paginiOSTEOTORACELEVlaicu Tudor100% (1)
- Regiuni Anatomo-Clinice Ale GâtuluiDocument39 paginiRegiuni Anatomo-Clinice Ale GâtuluiPricop NicolaÎncă nu există evaluări
- Sistemul Nervos VegetativDocument42 paginiSistemul Nervos VegetativLore Boboc100% (1)
- MediastinDocument11 paginiMediastinhedstrongÎncă nu există evaluări
- Anatomia Sistemului Circulator.Document12 paginiAnatomia Sistemului Circulator.Cristian MariusÎncă nu există evaluări
- Col. Vert PDFDocument54 paginiCol. Vert PDFNicoli RoscaÎncă nu există evaluări
- 1 Topografia Organelor Și A Sistemelor de OrganeDocument2 pagini1 Topografia Organelor Și A Sistemelor de OrganeRusu Maria BiancaÎncă nu există evaluări
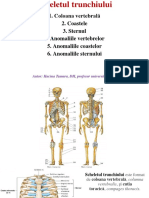
- Scheletul Axial - Anatomia Omului, LP 1, Biologie, Anul IDocument6 paginiScheletul Axial - Anatomia Omului, LP 1, Biologie, Anul IMaria GorganÎncă nu există evaluări
- REGIUNILE MEMBRULUI INFERIOR-presentationDocument20 paginiREGIUNILE MEMBRULUI INFERIOR-presentationIon Popescu100% (1)
- NeurocraniulDocument18 paginiNeurocraniulGeorgiana StateÎncă nu există evaluări
- Organele de SimtDocument15 paginiOrganele de SimtMadalina Ungureanu100% (1)
- Coloana, Peretele ToracicDocument49 paginiColoana, Peretele ToracicIwannaÎncă nu există evaluări
- Analizatorul CutanatDocument16 paginiAnalizatorul Cutanatpaun_alexandra89Încă nu există evaluări
- Articulatiile CostoDocument1 paginăArticulatiile CostoCebotari IvanÎncă nu există evaluări
- Reglarea Cordului Si VaselorDocument14 paginiReglarea Cordului Si VaselorgraceinÎncă nu există evaluări
- Reflex eDocument10 paginiReflex eMadalina MarinÎncă nu există evaluări
- Analizatorul KinestezicDocument7 paginiAnalizatorul Kinestezicstefan manoleÎncă nu există evaluări
- Sistemul Nervos Vegetativ AutonomDocument10 paginiSistemul Nervos Vegetativ AutonomAncaVaduvaÎncă nu există evaluări
- 3.trunchiul CerebralDocument8 pagini3.trunchiul CerebralCristina DorofteÎncă nu există evaluări
- Sub EmbriologieDocument16 paginiSub EmbriologieSandu Emanuela RamonaÎncă nu există evaluări
- Muschii PrezentareDocument12 paginiMuschii Prezentarerugina simona100% (1)
- Lista Nervilor CranieniDocument3 paginiLista Nervilor CranienidianamgggÎncă nu există evaluări
- Analizatorul CutanatDocument14 paginiAnalizatorul CutanatMalina Irimia100% (1)
- L.P. 5 PDFDocument153 paginiL.P. 5 PDFMarius CodrescuÎncă nu există evaluări
- MERSULDocument12 paginiMERSULSima Denisa Laura100% (1)
- Coloana Lombara Curs 8Document6 paginiColoana Lombara Curs 8Gligan Alin-CristianÎncă nu există evaluări
- Fiziologia Obiect de StudiuDocument16 paginiFiziologia Obiect de StudiudimaÎncă nu există evaluări
- Subiectul II - Examen Anatomie, An 2, Semestrul 1Document63 paginiSubiectul II - Examen Anatomie, An 2, Semestrul 1StoicaAlfredÎncă nu există evaluări
- Nervul XDocument3 paginiNervul XNicoleta TurcuÎncă nu există evaluări
- Anatomia Sistemului MuscularDocument17 paginiAnatomia Sistemului MuscularNicoleta NastaseÎncă nu există evaluări
- Semio 10Document19 paginiSemio 10Lauren GrayÎncă nu există evaluări
- AbdomenDocument32 paginiAbdomenValentina CojocaruÎncă nu există evaluări
- Curs 5Document4 paginiCurs 5Alexandra TițaÎncă nu există evaluări
- Subiecte Examen Anatomie An I Sem II 2014-2015 FinalDocument3 paginiSubiecte Examen Anatomie An I Sem II 2014-2015 FinalhospitalbucharestÎncă nu există evaluări
- Coloana Cervicala Curs 6Document4 paginiColoana Cervicala Curs 6Gligan Alin-CristianÎncă nu există evaluări
- Nervii SpinaliDocument3 paginiNervii SpinaliDanutz MariusÎncă nu există evaluări
- 2.sistemul Nervos CentralDocument66 pagini2.sistemul Nervos CentralAky AkyyÎncă nu există evaluări
- Fiziopatologia DureriiDocument17 paginiFiziopatologia DureriiGligor Andrada100% (1)
- Oasele Şi Articulaţiile MâiniiDocument10 paginiOasele Şi Articulaţiile MâiniiPatricia DrăganÎncă nu există evaluări
- Sistem NervosDocument61 paginiSistem NervosIonuț BuburuzanÎncă nu există evaluări
- Maduva Spinarii Subst CenusieDocument24 paginiMaduva Spinarii Subst CenusieIoanna Bianca HÎncă nu există evaluări
- Tabel Muschii DisertatieDocument5 paginiTabel Muschii DisertatieMihai CimpocaÎncă nu există evaluări
- Anatomie Cursurile 1-11-2022Document102 paginiAnatomie Cursurile 1-11-2022ivan ionelaÎncă nu există evaluări
- Teme - Anatomia Analizatorului VizualDocument34 paginiTeme - Anatomia Analizatorului Vizualcomanflavius2262Încă nu există evaluări
- Anatomia Corpului Uman-Functiile OrganismuluiDocument43 paginiAnatomia Corpului Uman-Functiile OrganismuluiMihaela StoicaÎncă nu există evaluări
- Sdr. WhiplashDocument9 paginiSdr. WhiplashAndra MonicaÎncă nu există evaluări
- Analizator KinestezicDocument2 paginiAnalizator KinestezicRemus Coman100% (1)
- Curs 3miologieDocument37 paginiCurs 3miologieVlaicu Tudor100% (1)
- Receptia ExcitariDocument23 paginiReceptia ExcitariElena KuhaskÎncă nu există evaluări
- Functia Somestezica PDFDocument12 paginiFunctia Somestezica PDFNehoianu LilianaÎncă nu există evaluări
- Capitolul VII Analizatori1Document19 paginiCapitolul VII Analizatori1Jora Ion100% (1)
- 6-Sensibilitatea Si Durerea Var 2Document65 pagini6-Sensibilitatea Si Durerea Var 2TheBlackChinkÎncă nu există evaluări