S-ar putea să vă placă și
- Perturbari Ale MetabolismuluiDocument2 paginiPerturbari Ale MetabolismuluiLoretta IftinchiÎncă nu există evaluări
- Plan Lectie ProtozoareDocument4 paginiPlan Lectie ProtozoareMadalina Elena DinuÎncă nu există evaluări
- Hormonii S10Document12 paginiHormonii S10AAMC77Încă nu există evaluări
- IndrumarDocument36 paginiIndrumarДобрый МолодецÎncă nu există evaluări
- Microbiologie - VirusuriDocument14 paginiMicrobiologie - VirusuriAlexandra LeahuÎncă nu există evaluări
- IMUNOLOGIEDocument14 paginiIMUNOLOGIEMihaela StavaracheÎncă nu există evaluări
- Metode de Microscopie S.A.Document30 paginiMetode de Microscopie S.A.Mirela Bojoga100% (1)
- Electroforeza Adn UluiDocument2 paginiElectroforeza Adn UluiFlorea RodicaÎncă nu există evaluări
- ADN, ARN - Mecanisme Biochimice În Procesarea Informație La Nivel CelularDocument15 paginiADN, ARN - Mecanisme Biochimice În Procesarea Informație La Nivel CelularABIGAIL KALMARÎncă nu există evaluări
- Biologie 2011Document11 paginiBiologie 2011RoxanaMateiÎncă nu există evaluări
- Genetica MolecularaDocument7 paginiGenetica MolecularaClaudia GrosuÎncă nu există evaluări
- Olimpiada de Biologie 2016Document19 paginiOlimpiada de Biologie 2016GeorgiaStefanÎncă nu există evaluări
- Excitabilitatea Si Legile AcesteiaDocument20 paginiExcitabilitatea Si Legile AcesteiaMarinica UlmeanuÎncă nu există evaluări
- OBSTRETICA SI GINECOLOGIE - Anul 3 E AMGDocument22 paginiOBSTRETICA SI GINECOLOGIE - Anul 3 E AMGAlexandra LeahuÎncă nu există evaluări
- Mecanismele Genetice Ale Diversitatii ImunoglobulinelorDocument7 paginiMecanismele Genetice Ale Diversitatii ImunoglobulinelorGumus MedelaineÎncă nu există evaluări
- Aluminiu ReferatDocument11 paginiAluminiu ReferatAlexandra LeahuÎncă nu există evaluări
- Bacterii. Protiste. Ciuperci - RecapitulareDocument1 paginăBacterii. Protiste. Ciuperci - RecapitulareMadalina UngureanuÎncă nu există evaluări
- Dizertatie Raul FinalDocument59 paginiDizertatie Raul FinalAlexandra LeahuÎncă nu există evaluări
- Nutritia Autotrofa Si Heterotrofa. Digestia Mamiferelor Clasa A X - ADocument6 paginiNutritia Autotrofa Si Heterotrofa. Digestia Mamiferelor Clasa A X - AClaudia Bucur0% (1)
- Structura BacteriilorDocument11 paginiStructura BacteriilorAlexandra G. Cazan0% (1)
- MicrobiologieDocument76 paginiMicrobiologieMirela Florinela RotaruÎncă nu există evaluări
- Studiul Diviziunii MitoticeDocument7 paginiStudiul Diviziunii MitoticeEstephy DeeÎncă nu există evaluări
- Biologie Xi CorigentaDocument3 paginiBiologie Xi CorigentaBlajiu BeatriceÎncă nu există evaluări
- Increngatura Spermatophyta (Antophyta, Phanerogamae)Document85 paginiIncrengatura Spermatophyta (Antophyta, Phanerogamae)Madalina AndradaÎncă nu există evaluări
- Prezentare AnticorpiDocument15 paginiPrezentare AnticorpiOanaDorofteÎncă nu există evaluări
- Caracterele Generale Ale AmfibienilorDocument9 paginiCaracterele Generale Ale AmfibienilorFerma MosneniÎncă nu există evaluări
- Eco Atitudini NR 3Document59 paginiEco Atitudini NR 3Alexandra LeahuÎncă nu există evaluări
- Tema BioDocument6 paginiTema BioClaudia ClaraÎncă nu există evaluări
- Micro CursDocument37 paginiMicro CursMaria JalbaÎncă nu există evaluări
- Histologie - SintezeDocument28 paginiHistologie - SintezeGruia AlexandraÎncă nu există evaluări
- Zone Si Arii Protejate Si Valorificarea Lor in TurismDocument73 paginiZone Si Arii Protejate Si Valorificarea Lor in TurismAlexandra Leahu100% (1)
- Histologie CitoscheletDocument46 paginiHistologie CitoscheletAnamaria ConstantinÎncă nu există evaluări
- Rezumate Paraziti Examen CPZ 1Document7 paginiRezumate Paraziti Examen CPZ 1Nicole AdamcsekÎncă nu există evaluări
- Botanica Notiuni IntroductiveDocument3 paginiBotanica Notiuni IntroductiveMinciuna Alina100% (1)
- Afis Concurs Judetean Caleidoscopul Stiintelor 2023Document4 paginiAfis Concurs Judetean Caleidoscopul Stiintelor 2023꧁Cʜᴇʀʀʏ꧂Încă nu există evaluări
- OvogenezaDocument7 paginiOvogenezaSimonaÎncă nu există evaluări
- Infectii Ale Pielii Si Tesuturilor MoiDocument61 paginiInfectii Ale Pielii Si Tesuturilor Moiucim navzarÎncă nu există evaluări
- 4.3. Cresterea Si Reproducerea MucegaiurilorDocument7 pagini4.3. Cresterea Si Reproducerea MucegaiurilorBrînza VladÎncă nu există evaluări
- Curs 10 BacteriocineDocument26 paginiCurs 10 BacteriocineIoana PopescuÎncă nu există evaluări
- MeristemeDocument4 paginiMeristemeMarcelaMorosanu0% (1)
- CancerDocument5 paginiCancerpisimiau07Încă nu există evaluări
- 058 Conexiunile Hipotalamusului Cu Hipofiza Si RetinaDocument1 pagină058 Conexiunile Hipotalamusului Cu Hipofiza Si RetinaLăcătușu IulianÎncă nu există evaluări
- Coloratia VitalaDocument3 paginiColoratia Vitalalalu15Încă nu există evaluări
- Absorbția IntestinalăDocument7 paginiAbsorbția Intestinalăscoot vlog SMÎncă nu există evaluări
- CCM2 - Colesnic VasileDocument18 paginiCCM2 - Colesnic VasileVasile ColesnicÎncă nu există evaluări
- LenticeleDocument3 paginiLenticeleCris1234450% (2)
- Genetica Si CancerulDocument12 paginiGenetica Si CancerulElena CaranicaÎncă nu există evaluări
- PinocitozaDocument15 paginiPinocitozaIonut Bojor100% (1)
- IS - Rezultate Evaluare Concurs DirectoriDocument8 paginiIS - Rezultate Evaluare Concurs DirectoriDana UrsuleasaÎncă nu există evaluări
- Gjavantajele Hibridarii Somatice-ReferatbunDocument15 paginiGjavantajele Hibridarii Somatice-ReferatbunfabipetÎncă nu există evaluări
- Spermatogeneza OvogenezaDocument4 paginiSpermatogeneza OvogenezaM91tinaDrokiaÎncă nu există evaluări
- Comportamentul Perusilor in CaptivitateDocument2 paginiComportamentul Perusilor in CaptivitatemichelbrestonÎncă nu există evaluări
- Test Sistemul Excretor - 6 Blaj GherghinaDocument3 paginiTest Sistemul Excretor - 6 Blaj GherghinaRadu FloricaÎncă nu există evaluări
- Proiect HG Cu Privire La Aprobarea Programului Activităților de Reintegrare A Ţării Pentru Anul 2023Document5 paginiProiect HG Cu Privire La Aprobarea Programului Activităților de Reintegrare A Ţării Pentru Anul 2023News UngheniÎncă nu există evaluări
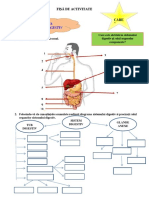
- Fisă Sistem DigestivDocument1 paginăFisă Sistem DigestivMarcela EmanuelaÎncă nu există evaluări
- Genetica TestDocument2 paginiGenetica TestBiancaÎncă nu există evaluări
- Dezvoltarea Embrionară A Sistemului RespiratorDocument1 paginăDezvoltarea Embrionară A Sistemului RespiratorYoana AlecsandraÎncă nu există evaluări
- Arborele CirculatorDocument3 paginiArborele CirculatorAlexandraÎncă nu există evaluări
- Referat SportDocument8 paginiReferat SportDenisa SanduÎncă nu există evaluări
- Embriologie Curs 1 Si 2Document89 paginiEmbriologie Curs 1 Si 2Cristina DobrinÎncă nu există evaluări
- Reproducerea La Gimnosperme Si AngiospermeDocument13 paginiReproducerea La Gimnosperme Si AngiospermeloreÎncă nu există evaluări
- LP 12 - Antivirotice Antimicotice, AntituberculoaseDocument63 paginiLP 12 - Antivirotice Antimicotice, AntituberculoaseRoxana CutasÎncă nu există evaluări
- Lec 3 Structura Celulei BacterieneDocument28 paginiLec 3 Structura Celulei BacterieneMariana GhermanÎncă nu există evaluări
- Catb 4Document3 paginiCatb 4Diana ValentinaÎncă nu există evaluări
- Microbiologie, Bacteriologie, GeneralitatiDocument8 paginiMicrobiologie, Bacteriologie, GeneralitatiRoberta GrosuÎncă nu există evaluări
- Microbiologie Generala Intrebari de Control-35105-1Document23 paginiMicrobiologie Generala Intrebari de Control-35105-1Elena CataragaÎncă nu există evaluări
- OBSTRETICA SI GINECOLOGIE - Alexandra Anul 3 AMGDocument19 paginiOBSTRETICA SI GINECOLOGIE - Alexandra Anul 3 AMGAlexandra LeahuÎncă nu există evaluări
- Referat AluminiuDocument16 paginiReferat AluminiuAlexandra LeahuÎncă nu există evaluări
- RelatiiDocument7 paginiRelatiiAlexandra LeahuÎncă nu există evaluări
- 4 MorfologiaDocument5 pagini4 MorfologiaAlexandra LeahuÎncă nu există evaluări
- Microbiologie - CIUPERCILEDocument14 paginiMicrobiologie - CIUPERCILEAlexandra LeahuÎncă nu există evaluări
- Istoricul Și Importanţa Teledetecției Și FotogrammetrieiDocument3 paginiIstoricul Și Importanţa Teledetecției Și FotogrammetrieiAlexandra LeahuÎncă nu există evaluări
- Indici Muntii ParangDocument16 paginiIndici Muntii ParangAlexandra LeahuÎncă nu există evaluări
- Carpatii Meridionali - Grupa ParangDocument18 paginiCarpatii Meridionali - Grupa ParangAlexandra LeahuÎncă nu există evaluări