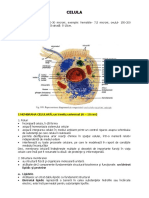
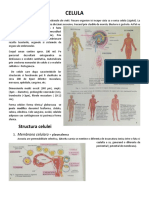
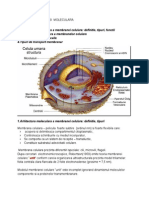
S-ar putea să vă placă și
- Homeostazia Celulara - Proiect BiologieDocument23 paginiHomeostazia Celulara - Proiect BiologieDaiana RotaruÎncă nu există evaluări
- Niveluri de OrganizareDocument12 paginiNiveluri de Organizareiulia buhaiÎncă nu există evaluări
- Referat Membrana CelularaDocument15 paginiReferat Membrana CelularaRazvan_GabrielÎncă nu există evaluări
- Consideratii Generale Ale Permeabilitatii Si Transportului Prin MembraneDocument51 paginiConsideratii Generale Ale Permeabilitatii Si Transportului Prin MembraneAdriana VillarrealÎncă nu există evaluări
- Celula Si SCH Prin MBDocument10 paginiCelula Si SCH Prin MBTeodora VasileÎncă nu există evaluări
- Proprietățile CeluleiDocument10 paginiProprietățile CeluleiTurlea FloareaÎncă nu există evaluări
- Fenomene de TransferDocument23 paginiFenomene de TransferStancioiuBalaciLauraÎncă nu există evaluări
- OsmozaDocument4 paginiOsmozaviorica serbanÎncă nu există evaluări
- Raspunsuri La Intrebarile de La Prelegeri Biofizica ExamenDocument4 paginiRaspunsuri La Intrebarile de La Prelegeri Biofizica ExamenpzdtskblÎncă nu există evaluări
- Membrana BiologiceDocument47 paginiMembrana BiologiceNastiu6aÎncă nu există evaluări
- Transportul SubstantelorDocument52 paginiTransportul SubstantelorAna MadinaÎncă nu există evaluări
- Biofizica CelularaDocument17 paginiBiofizica CelularaAlexandra ArdeleanÎncă nu există evaluări
- Subiecte Biofizica Rezolvate 2012Document18 paginiSubiecte Biofizica Rezolvate 2012gaby90009Încă nu există evaluări
- Curs 2 - Transporturi MembranareDocument59 paginiCurs 2 - Transporturi MembranareAdrian UrsuÎncă nu există evaluări
- Transportul Prin Membrana CelularaDocument7 paginiTransportul Prin Membrana CelularaTeodora Elena100% (1)
- Bio Cel Curs 6 Transportul TransmembranarDocument54 paginiBio Cel Curs 6 Transportul TransmembranarMarin TheodorÎncă nu există evaluări
- FiziologieDocument28 paginiFiziologieCatalina UrsacheÎncă nu există evaluări
- Celula UmanaDocument6 paginiCelula UmanaAndreea F92% (12)
- Schimbul de Substanțe Dintre Celulă Și Mediul AmbiantDocument5 paginiSchimbul de Substanțe Dintre Celulă Și Mediul AmbiantRadu MarianÎncă nu există evaluări
- Biofizica LPDocument12 paginiBiofizica LPIulya BraghesÎncă nu există evaluări
- ANATOMIEDocument29 paginiANATOMIEAlexandra Ana-Maria IancuÎncă nu există evaluări
- Subiecte PT Exam Oral Biofarmacie (Varianta Adaugita)Document42 paginiSubiecte PT Exam Oral Biofarmacie (Varianta Adaugita)Cinaru Catalina MariaÎncă nu există evaluări
- Celula UmanaDocument8 paginiCelula UmanaCatalina CiocanÎncă nu există evaluări
- Fizica IntrebariDocument20 paginiFizica IntrebariStefania PatroleaÎncă nu există evaluări
- CelulaDocument7 paginiCelulaAlexÎncă nu există evaluări
- Biomecanica CeluleiDocument24 paginiBiomecanica CeluleiCristina ElenaÎncă nu există evaluări
- Curs 4 BIOFIZDocument72 paginiCurs 4 BIOFIZfloricapopovici168Încă nu există evaluări
- Organizare Celulare IDocument14 paginiOrganizare Celulare ISmadu AdrianaÎncă nu există evaluări
- Totalizare Fiziologie1 1Document58 paginiTotalizare Fiziologie1 1Ana DubceacÎncă nu există evaluări
- Bilant de MaterialeDocument27 paginiBilant de Materialelyly1601Încă nu există evaluări
- Cursul IIIDocument8 paginiCursul IIIana-maria100% (1)
- Electro DializaDocument15 paginiElectro DializaPatruta Cristina AndreeaÎncă nu există evaluări
- CELULADocument7 paginiCELULAMadalina FoteaÎncă nu există evaluări
- Cursul IiiDocument7 paginiCursul IiiMarianÎncă nu există evaluări
- Barrons 03Document28 paginiBarrons 03Enache Andreea100% (2)
- Fiziologie 1Document37 paginiFiziologie 1Bianca GherghelucaÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- Structura Şi Ultrastructura CeluleiDocument34 paginiStructura Şi Ultrastructura CeluleiNadeaÎncă nu există evaluări
- L6-Fenomene Moleculare de Transport-88753Document37 paginiL6-Fenomene Moleculare de Transport-88753Adi PorcescuÎncă nu există evaluări
- CELULADocument18 paginiCELULASabina PostolacheÎncă nu există evaluări
- Biofizica 7Document34 paginiBiofizica 7Lile George100% (1)
- Celula Eucariota Structura Si UltrastructuraDocument19 paginiCelula Eucariota Structura Si UltrastructuraFlorentina Claudia Calusaru100% (2)
- Fiziologie SubiecteDocument22 paginiFiziologie SubiecteMareș Ion-AlexandruÎncă nu există evaluări
- Transportul Prin Membranele CelulareDocument11 paginiTransportul Prin Membranele CelulareSandru ValiÎncă nu există evaluări
- Celula, Proprietatile CeluleiDocument12 paginiCelula, Proprietatile CeluleiAlexÎncă nu există evaluări
- Structura Membranei CelulareDocument11 paginiStructura Membranei CelulareOana Palade100% (2)
- BiofizicaDocument74 paginiBiofizicaMonica RăileanuÎncă nu există evaluări
- Referat BiofizicaDocument10 paginiReferat Biofizicaraduvelescu100% (1)
- BiocelDocument24 paginiBiocelficioruÎncă nu există evaluări
- Membrana CelularaDocument77 paginiMembrana CelularaCătălina RaicuÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- Transportul Activ Și Transportul PasivDocument5 paginiTransportul Activ Și Transportul PasivRareș RusÎncă nu există evaluări
- Excitabilitatea Si Legile AcesteiaDocument20 paginiExcitabilitatea Si Legile AcesteiaMarinica UlmeanuÎncă nu există evaluări
- Curs 6Document28 paginiCurs 6Ioana ChindrisÎncă nu există evaluări
- Cursul VIDocument45 paginiCursul VIionutbranza19Încă nu există evaluări
- Celula EucariotaDocument5 paginiCelula EucariotaStefan GhergheÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Examen Bacteriologic Secretii, Plagi, Fistule, AbceseDocument2 paginiExamen Bacteriologic Secretii, Plagi, Fistule, AbceseDiana AlexandraÎncă nu există evaluări
- 13.1 Curs BI MD TrichinelozaDocument3 pagini13.1 Curs BI MD TrichinelozaDiana AlexandraÎncă nu există evaluări
- 9.3 Curs BI MD LeptospirozeDocument2 pagini9.3 Curs BI MD LeptospirozeDiana AlexandraÎncă nu există evaluări
- Caz Clinic - DemențăDocument1 paginăCaz Clinic - DemențăDiana AlexandraÎncă nu există evaluări
- LEISHMANIOZEDocument3 paginiLEISHMANIOZEDiana AlexandraÎncă nu există evaluări
- MALARIADocument5 paginiMALARIADiana AlexandraÎncă nu există evaluări
- GLOMERULONEFRITELEDocument28 paginiGLOMERULONEFRITELEDiana AlexandraÎncă nu există evaluări
- Obiective Operationale - Rusu Diana-AlexandraDocument1 paginăObiective Operationale - Rusu Diana-AlexandraDiana AlexandraÎncă nu există evaluări
- Curs 3+4 TiroidaDocument16 paginiCurs 3+4 TiroidaDiana AlexandraÎncă nu există evaluări
- StafilocociDocument9 paginiStafilocociDiana AlexandraÎncă nu există evaluări
- Colocviu HistoDocument1 paginăColocviu HistoDiana AlexandraÎncă nu există evaluări