S-ar putea să vă placă și
- Particularitatile Anesteziei La Pacienti Cu Teren HepaticDocument7 paginiParticularitatile Anesteziei La Pacienti Cu Teren Hepaticvictor100% (1)
- AritmiileDocument7 paginiAritmiileAngelina BulaiÎncă nu există evaluări
- CapulDocument16 paginiCapulAla Grițcan-BîtcaÎncă nu există evaluări
- Hipnotice N.bacinschiDocument34 paginiHipnotice N.bacinschiDomnita UrsuÎncă nu există evaluări
- ColinergiceDocument3 paginiColinergiceStefan TiganuÎncă nu există evaluări
- Metabolismul GlucidicDocument48 paginiMetabolismul GlucidicBrinza Olga100% (1)
- Sindroame Pleurale CursDocument50 paginiSindroame Pleurale CursMadelaineDobreÎncă nu există evaluări
- Traumotologie RaspunsuriDocument36 paginiTraumotologie RaspunsuriSt StÎncă nu există evaluări
- Semiologie SDR Insuficientei Coronariene Angina Pectorala Bezu MedicinaDocument47 paginiSemiologie SDR Insuficientei Coronariene Angina Pectorala Bezu MedicinaLia ScutaruÎncă nu există evaluări
- Caragat Supurative-5583Document57 paginiCaragat Supurative-5583AnnitaCristalÎncă nu există evaluări
- 1.2 - Apendicita Acuta Si CronicaDocument41 pagini1.2 - Apendicita Acuta Si Cronica22194Încă nu există evaluări
- Subiecte Rezolvate Morfopatologie CantacuzinoDocument14 paginiSubiecte Rezolvate Morfopatologie CantacuzinoLuiza BezneaÎncă nu există evaluări
- Urologie OralDocument79 paginiUrologie OralAnna StolearencoÎncă nu există evaluări
- Curs 5 - FIZIOPATOLOGIA DZDocument16 paginiCurs 5 - FIZIOPATOLOGIA DZcmvioÎncă nu există evaluări
- Farma BileteDocument17 paginiFarma BileteRenata ProsutinskaiaÎncă nu există evaluări
- Tulburari de CoagulareDocument7 paginiTulburari de CoagulareCellaÎncă nu există evaluări
- PeritoniteleDocument46 paginiPeritoniteleElla LenusikÎncă nu există evaluări
- Subiecte Sem 2 MorfopatologieDocument21 paginiSubiecte Sem 2 MorfopatologieAlexandra StuparuÎncă nu există evaluări
- Curs 3 Sindroamele Radiologice PulmonareDocument5 paginiCurs 3 Sindroamele Radiologice PulmonareUzea Cezar-DanÎncă nu există evaluări
- Chirurgie - ApendicitaDocument5 paginiChirurgie - ApendicitaTina AcridahÎncă nu există evaluări
- Colocviu NeuroDocument36 paginiColocviu NeuroMariaBoicuÎncă nu există evaluări
- Subiecte Morfopat An III Sem IDocument61 paginiSubiecte Morfopat An III Sem IElena-Dana Oprea100% (1)
- Sindrom KallmanaDocument19 paginiSindrom KallmanaДарья КопачинскаяÎncă nu există evaluări
- Mituri Opioide1 PDFDocument15 paginiMituri Opioide1 PDFCristina MartincuÎncă nu există evaluări
- Fiziopatologie Curs - Lista Subiecte (An III)Document2 paginiFiziopatologie Curs - Lista Subiecte (An III)Cristina VÎncă nu există evaluări
- Febra Hemoragica Cu Sindrom RenalDocument15 paginiFebra Hemoragica Cu Sindrom RenalАнтонина ПолизоÎncă nu există evaluări
- 1 Leziuni-CelulareDocument76 pagini1 Leziuni-CelulareBistrea VictorÎncă nu există evaluări
- Fisa de ObservatieDocument5 paginiFisa de ObservatieTatiana StanescuÎncă nu există evaluări
- Teste - Rezidenti Ati Cu RaspunsuriDocument150 paginiTeste - Rezidenti Ati Cu RaspunsuriPopescu VioricaÎncă nu există evaluări
- Acizii Nucleici-6084Document157 paginiAcizii Nucleici-6084anabellevanhelsingÎncă nu există evaluări
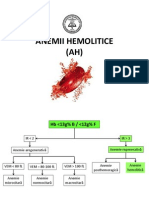
- Anemii HemoliticeDocument56 paginiAnemii HemoliticeViorel ValentinÎncă nu există evaluări
- Adrenomimeticele, AdrenoblocanteleDocument11 paginiAdrenomimeticele, AdrenoblocanteleNastea XdxÎncă nu există evaluări
- Urge ExDocument12 paginiUrge ExCristina LaÎncă nu există evaluări
- Insuficienta Hepatica 2015Document119 paginiInsuficienta Hepatica 2015Andra-Elena ObrejaÎncă nu există evaluări
- Leziuni CelulareDocument25 paginiLeziuni CelulareMelissa MullenÎncă nu există evaluări
- Tema 3Document16 paginiTema 3Bunici MarinelaÎncă nu există evaluări
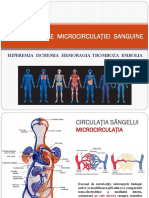
- Tulburări Ale Microcirculaţiei SanguineDocument31 paginiTulburări Ale Microcirculaţiei SanguineOlga-Alina Rada0% (1)
- Marchiafava MicheliDocument16 paginiMarchiafava Micheliheavysky1Încă nu există evaluări
- Indicatii Total.3 FarmaDocument1 paginăIndicatii Total.3 FarmaCristian SîrbuÎncă nu există evaluări
- Sincopa CardiacaDocument7 paginiSincopa CardiacaDanaÎncă nu există evaluări
- Imagini MorfopatDocument49 paginiImagini MorfopatOanaIaconiÎncă nu există evaluări
- ANESTEZIOLOGIADocument11 paginiANESTEZIOLOGIAjeka222100% (1)
- Subiecte Fiziologie Practic Sem 2Document37 paginiSubiecte Fiziologie Practic Sem 2Aura ConstantinÎncă nu există evaluări
- Semiologia Chirurgicala A Sistemului VenosDocument3 paginiSemiologia Chirurgicala A Sistemului VenosRove PatriciaÎncă nu există evaluări
- NeuroDocument68 paginiNeuroAnonymous NqTjEcÎncă nu există evaluări
- Compliment Multiplu La UrologieDocument10 paginiCompliment Multiplu La UrologieMarin DulghieruÎncă nu există evaluări
- Lab 5 AFU 2Document14 paginiLab 5 AFU 2Stina Adrea-ElisaÎncă nu există evaluări
- ExamenDocument86 paginiExamenvasilisanourÎncă nu există evaluări
- Cid in Obstetric ADocument2 paginiCid in Obstetric AClaudia OpreaÎncă nu există evaluări
- Activitatea Electrica A InimiiDocument32 paginiActivitatea Electrica A InimiiOana RaduÎncă nu există evaluări
- Cursuri FizioDocument62 paginiCursuri FizioIuly Manac100% (1)
- FiziologieDocument29 paginiFiziologieLidia PopÎncă nu există evaluări
- Proprietati Fundamentale Ale Muschiului CardiacDocument26 paginiProprietati Fundamentale Ale Muschiului Cardiacema 274Încă nu există evaluări
- BIOFIZICADocument6 paginiBIOFIZICAOana RaduÎncă nu există evaluări
- Curs 1 - InimaDocument43 paginiCurs 1 - InimaRux JitaruÎncă nu există evaluări
- AntiaritmiceDocument17 paginiAntiaritmicebrianed231Încă nu există evaluări
- Fizio Curs Sem 1Document101 paginiFizio Curs Sem 1elenabÎncă nu există evaluări
- Activitatea Electrică A InimiiDocument39 paginiActivitatea Electrică A InimiiYasminaB100% (2)
- Anul 2 Sem 1 Tema 7 Fibre Cardiace Sistemul ExcitoconductorDocument14 paginiAnul 2 Sem 1 Tema 7 Fibre Cardiace Sistemul ExcitoconductorNicoletaÎncă nu există evaluări
- Fiziologie CardiacăDocument30 paginiFiziologie CardiacăaiusvestaÎncă nu există evaluări
- Legea 487 Din 2002 Republicata 2012 ModificataDocument7 paginiLegea 487 Din 2002 Republicata 2012 ModificataMihaela VișanÎncă nu există evaluări
- ADOS Manual RO v0.1Document121 paginiADOS Manual RO v0.1Mihaela Vișan100% (1)
- BenzodiazepineDocument2 paginiBenzodiazepineMihaela VișanÎncă nu există evaluări
- 24.curs PericarditeDocument67 pagini24.curs PericarditeMihaela VișanÎncă nu există evaluări
- Sindromul WolffDocument4 paginiSindromul WolffMihaela VișanÎncă nu există evaluări