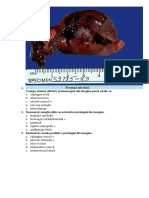
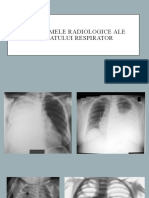
S-ar putea să vă placă și
- Întrebari Pentru Examen La Disciplina Endocrinologie Pentru Studenţii Anului 4Document103 paginiÎntrebari Pentru Examen La Disciplina Endocrinologie Pentru Studenţii Anului 4Radu GherbÎncă nu există evaluări
- Preparatele Cu Acţiune Asupra Inervaţiei PerifericeDocument12 paginiPreparatele Cu Acţiune Asupra Inervaţiei PerifericeCarolina TișcencoÎncă nu există evaluări
- Medicamente ObligatoriiDocument57 paginiMedicamente ObligatoriiJora Ion100% (1)
- VIDocument5 paginiVIBunici MarinelaÎncă nu există evaluări
- cursII PDFDocument12 paginicursII PDFPetru DariiÎncă nu există evaluări
- TesteDocument25 paginiTesteCristina NeghinaÎncă nu există evaluări
- Exam NEFRO 1Document27 paginiExam NEFRO 1Olga GuțanÎncă nu există evaluări
- 1568490488504825 (2) (1)Document28 pagini1568490488504825 (2) (1)Renata ProsutinskaiaÎncă nu există evaluări
- Lab 5 AFU 2Document14 paginiLab 5 AFU 2Stina Adrea-ElisaÎncă nu există evaluări
- Mihai FisaDocument13 paginiMihai FisaAnastasia VlasÎncă nu există evaluări
- Lectia 1Document53 paginiLectia 1Elena PavlicÎncă nu există evaluări
- Bilete 1 56 FinalDocument320 paginiBilete 1 56 FinalMusic playÎncă nu există evaluări
- Teste - Rezidenti Ati Cu RaspunsuriDocument150 paginiTeste - Rezidenti Ati Cu RaspunsuriPopescu VioricaÎncă nu există evaluări
- Echilibrul Acido-Bazic (Magaleas Serghei, F1804)Document13 paginiEchilibrul Acido-Bazic (Magaleas Serghei, F1804)Alexandra MagaleasÎncă nu există evaluări
- Fisa de ObservatieDocument5 paginiFisa de ObservatieTatiana StanescuÎncă nu există evaluări
- Raspunsurile La Farma ClinicaDocument33 paginiRaspunsurile La Farma ClinicaPasha StupelimanÎncă nu există evaluări
- Preparate IulianDocument61 paginiPreparate IulianCimpoi AureliaÎncă nu există evaluări
- Master 1Document63 paginiMaster 1Corina GrosuÎncă nu există evaluări
- RecipeDocument6 paginiRecipeBenzari MarinaÎncă nu există evaluări
- Intrebări Atestarea II Medicina 1Document20 paginiIntrebări Atestarea II Medicina 1M SÎncă nu există evaluări
- 2 5370827380690525915Document49 pagini2 5370827380690525915Vlad PopaÎncă nu există evaluări
- Tema 4Document11 paginiTema 4marinacretu1Încă nu există evaluări
- Fizpat 1Document52 paginiFizpat 1Danu PostolacheÎncă nu există evaluări
- 139-59 Intreb Noi!!!Document19 pagini139-59 Intreb Noi!!!Valeria SmatocÎncă nu există evaluări
- Doze Si Indicatii IVDocument1 paginăDoze Si Indicatii IVСергей МалайÎncă nu există evaluări
- Totalizarea 5 FizpatDocument11 paginiTotalizarea 5 FizpatMocanuIlieÎncă nu există evaluări
- Practic MorfopatDocument149 paginiPractic MorfopatGigi Och100% (1)
- Totalizarea 2Document9 paginiTotalizarea 2Biatricia GuţuÎncă nu există evaluări
- Fiziologia Sistemul NervosDocument27 paginiFiziologia Sistemul NervosAurelia HayateÎncă nu există evaluări
- Tot 3Document127 paginiTot 3dvgÎncă nu există evaluări
- Indicatii - Metodice - Antispirochetoase - Antituberculoase.Antileproase - Antiprotozoice.Antifungice - SpinosuDocument9 paginiIndicatii - Metodice - Antispirochetoase - Antituberculoase.Antileproase - Antiprotozoice.Antifungice - SpinosuJora IonÎncă nu există evaluări
- Vir MicroDocument11 paginiVir MicroCristina HaritonÎncă nu există evaluări
- Tema 2 DigestivDocument10 paginiTema 2 DigestivMihaela Botnari-GuţuÎncă nu există evaluări
- Excitatia Ritmica A CorduluiDocument9 paginiExcitatia Ritmica A CorduluiMihaela VișanÎncă nu există evaluări
- Curs 1 BFK PDFDocument35 paginiCurs 1 BFK PDFOctaviaCosminaÎncă nu există evaluări
- Anemii HemoliticeDocument56 paginiAnemii HemoliticeViorel ValentinÎncă nu există evaluări
- Cardiotonice Chisinau Craiova 2017 Stratu FinalDocument60 paginiCardiotonice Chisinau Craiova 2017 Stratu FinalАня ПрикопÎncă nu există evaluări
- TestefizDocument50 paginiTestefizGorodetchi PetruÎncă nu există evaluări
- Tabele FarmaDocument31 paginiTabele FarmaDorina Efros100% (1)
- DigestivdfgdsDocument11 paginiDigestivdfgdsvictor barbaÎncă nu există evaluări
- Metode de Examinare A TGIDocument43 paginiMetode de Examinare A TGIValerianBîcosÎncă nu există evaluări
- Tot 5 1Document6 paginiTot 5 1Elena PavlicÎncă nu există evaluări
- Remedii de Actiune Asupra Fifeitor Sisteme Si MecanismeDocument2 paginiRemedii de Actiune Asupra Fifeitor Sisteme Si Mecanismefanea_viorelÎncă nu există evaluări
- MicroDocument30 paginiMicroMariaÎncă nu există evaluări
- Morfo 4 2Document29 paginiMorfo 4 2Mihail PasaÎncă nu există evaluări
- Teste ChirDocument296 paginiTeste ChirSnejana Polișciuc100% (1)
- Tot 2 Sem 2 - 1614702610Document65 paginiTot 2 Sem 2 - 1614702610Cornel CornelÎncă nu există evaluări
- 1526279167172322Document60 pagini1526279167172322Razaria DailyneÎncă nu există evaluări
- Enunturi Examen 2021-22 Sem-5 FarmacologiemmmDocument10 paginiEnunturi Examen 2021-22 Sem-5 FarmacologiemmmElena L.Încă nu există evaluări
- Fizpat. SNC&EndoDocument88 paginiFizpat. SNC&EndoAngelina BulaiÎncă nu există evaluări
- Bazele Biochimice Ale Translaţiei 30 03-03 04 2020-6281Document9 paginiBazele Biochimice Ale Translaţiei 30 03-03 04 2020-6281LoredanaÎncă nu există evaluări
- Examen Practic 2020 2021 FIZIOLOGIE MD BUNDocument15 paginiExamen Practic 2020 2021 FIZIOLOGIE MD BUNVictorÎncă nu există evaluări
- Fizpat1 1Document65 paginiFizpat1 1Renata ProsutinskaiaÎncă nu există evaluări
- 7 - Electrocutare. ToxicoDocument14 pagini7 - Electrocutare. ToxicoCristian MihailescuÎncă nu există evaluări
- Întrebarea-Fiz-Master (1) 210119863Document54 paginiÎntrebarea-Fiz-Master (1) 210119863AlinaGutanu0% (1)
- Nervul Trigemen AnatomieDocument14 paginiNervul Trigemen AnatomieDunas DanÎncă nu există evaluări
- Afecțiunile Organelor Genitale Feminine - Patologia Sarcinii Şi LăuzieiDocument43 paginiAfecțiunile Organelor Genitale Feminine - Patologia Sarcinii Şi LăuzieiTanyaDonciuÎncă nu există evaluări
- Tema 3Document18 paginiTema 3Bunici MarinelaÎncă nu există evaluări
- Potentialul de ActiuneDocument12 paginiPotentialul de ActiuneBunici MarinelaÎncă nu există evaluări
- Mediatorii Chimici Ai SinapselorDocument28 paginiMediatorii Chimici Ai SinapselorSecuritate CRAIOVAÎncă nu există evaluări
- CalamitatiDocument4 paginiCalamitatiBunici MarinelaÎncă nu există evaluări
- Tema 1Document25 paginiTema 1Bunici MarinelaÎncă nu există evaluări
- 03 - CV - Curs 2 - AscultatiaDocument49 pagini03 - CV - Curs 2 - AscultatiaOlga Cretu HaidauÎncă nu există evaluări
- Examinarea Clinică in Afecţiuni CardiaceDocument38 paginiExaminarea Clinică in Afecţiuni CardiaceLucica Porsega VacarciucÎncă nu există evaluări
- Intrebari Anul III 2021-22 MG RomDocument4 paginiIntrebari Anul III 2021-22 MG RomInna995Încă nu există evaluări
- Reactiile Serologice09Document84 paginiReactiile Serologice09VLDs92% (13)
- Tema 6Document14 paginiTema 6Bunici MarinelaÎncă nu există evaluări
- Diagnostic SpirochetozeDocument13 paginiDiagnostic SpirochetozeАнастасия СиминенкоÎncă nu există evaluări
- ColinomimeticeDocument1 paginăColinomimeticeLoredanaÎncă nu există evaluări
- Radioscopia. DefiniţieDocument7 paginiRadioscopia. DefiniţieBunici MarinelaÎncă nu există evaluări
- Mana Anatomia ClinicaDocument40 paginiMana Anatomia ClinicaAmbros SerafimaÎncă nu există evaluări
- AdrenergiceDocument1 paginăAdrenergiceLoredanaÎncă nu există evaluări
- 4 5. Deregl. Circ. Sang. RegionaleDocument113 pagini4 5. Deregl. Circ. Sang. RegionaleBunici MarinelaÎncă nu există evaluări
- Reguli Pentru Elaborarea Lucrului IndividualDocument1 paginăReguli Pentru Elaborarea Lucrului IndividualBunici MarinelaÎncă nu există evaluări
- Totalizarea 2Document43 paginiTotalizarea 2Bunici MarinelaÎncă nu există evaluări
- Biochimie Teste ExamenDocument221 paginiBiochimie Teste ExamenIspas Nina100% (3)
- Comunicarea În Medicină Ca Baza Relației Medic - PacientDocument4 paginiComunicarea În Medicină Ca Baza Relației Medic - PacientBunici MarinelaÎncă nu există evaluări
- Raportul Organelor Cu PeritoneulDocument19 paginiRaportul Organelor Cu PeritoneulBunici MarinelaÎncă nu există evaluări
- Anatomia Toopografica A Peretelui Anterolateral Al AbdomenuluiDocument12 paginiAnatomia Toopografica A Peretelui Anterolateral Al AbdomenuluiBunici MarinelaÎncă nu există evaluări
- T 1Document26 paginiT 122194Încă nu există evaluări
- Tema 1 1 Fizologie MED RO-17402Document5 paginiTema 1 1 Fizologie MED RO-17402Bunici MarinelaÎncă nu există evaluări
- Întrebări Stud An IV Chirurgie Tehnici Op. 2014Document3 paginiÎntrebări Stud An IV Chirurgie Tehnici Op. 2014Maria GorodencoÎncă nu există evaluări
- 2Document3 pagini2Bunici MarinelaÎncă nu există evaluări
- Tipurile ComunicariiDocument2 paginiTipurile ComunicariiBunici MarinelaÎncă nu există evaluări
- Tema 1 1 Fizologie MED RO-17402Document5 paginiTema 1 1 Fizologie MED RO-17402Bunici MarinelaÎncă nu există evaluări
- Tesut Exitabil 2020-16957Document115 paginiTesut Exitabil 2020-16957Bunici MarinelaÎncă nu există evaluări
- Tema 4Document28 paginiTema 4Bunici Marinela0% (1)
- Intrebari Pe Capitole Romana M 2015 PDFDocument13 paginiIntrebari Pe Capitole Romana M 2015 PDFAnonymous nA8QoFiW4Încă nu există evaluări
- Potentialul de ActiuneDocument12 paginiPotentialul de ActiuneBunici MarinelaÎncă nu există evaluări