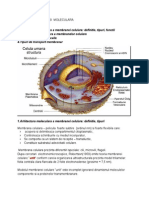
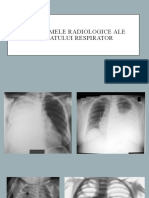
S-ar putea să vă placă și
- Structura Membranei CelulareDocument11 paginiStructura Membranei CelulareOana Palade100% (2)
- Biochimie Teste ExamenDocument221 paginiBiochimie Teste ExamenIspas Nina100% (3)
- Totalizare Fiziologie1 1Document58 paginiTotalizare Fiziologie1 1Ana DubceacÎncă nu există evaluări
- Homeostazia Celulara - Proiect BiologieDocument23 paginiHomeostazia Celulara - Proiect BiologieDaiana RotaruÎncă nu există evaluări
- BiofizicaDocument6 paginiBiofizicaMohammed ArarÎncă nu există evaluări
- Curs 1 Membrana CelularaDocument38 paginiCurs 1 Membrana CelularaBianca PopescuÎncă nu există evaluări
- Subiecte Rezolvate Complet PDFDocument71 paginiSubiecte Rezolvate Complet PDFCristian-Dorin GurauÎncă nu există evaluări
- Raspunsuri La ExamenDocument242 paginiRaspunsuri La ExamenDjMoldowa100% (3)
- Totalizarea 2Document43 paginiTotalizarea 2Bunici MarinelaÎncă nu există evaluări
- Tesut Exitabil 2022-75967Document120 paginiTesut Exitabil 2022-75967Vlad CorciuÎncă nu există evaluări
- Tesut Exitabil 2016Document103 paginiTesut Exitabil 2016Tuyu TufyuÎncă nu există evaluări
- Tesuturi Excitabile Nou 1Document103 paginiTesuturi Excitabile Nou 1Silvia Gîscă100% (1)
- Esuturi Excitabile 2022-58228Document131 paginiEsuturi Excitabile 2022-58228Viorica DandeșÎncă nu există evaluări
- Fiziologie Ro Curs 1 MD I 17-18Document46 paginiFiziologie Ro Curs 1 MD I 17-18Vladislava SavinÎncă nu există evaluări
- 2 Celula CompletDocument57 pagini2 Celula CompletBriana ChiriacÎncă nu există evaluări
- 1.2.fiziologia Membranei CelDocument48 pagini1.2.fiziologia Membranei CelgiuppiÎncă nu există evaluări
- Curs 4 Fiziologie An I Ro Seriile BD 2020Document41 paginiCurs 4 Fiziologie An I Ro Seriile BD 2020MITAÎncă nu există evaluări
- Fenomene de Transport Prin Membrana CelularaDocument38 paginiFenomene de Transport Prin Membrana CelularaNicoleta NicaÎncă nu există evaluări
- Curs 6Document34 paginiCurs 6Iulian FrunzăÎncă nu există evaluări
- CURS 2 - Mec. Excitab.Document58 paginiCURS 2 - Mec. Excitab.Miruna Alina Maria CarandaÎncă nu există evaluări
- Curs Fizio 1 - 2 RomDocument46 paginiCurs Fizio 1 - 2 RomOana MusteaţǎÎncă nu există evaluări
- Curs Fiziologie AnimalaDocument161 paginiCurs Fiziologie AnimalaStavarache LarisaÎncă nu există evaluări
- Curs - Celula Si TesuturileDocument22 paginiCurs - Celula Si TesuturileCristina ChiperÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- BiocelDocument24 paginiBiocelficioruÎncă nu există evaluări
- Fenomene de Transport MembranarDocument71 paginiFenomene de Transport MembranarPaula Mariana PrelucăÎncă nu există evaluări
- Curs - 1-2 - CELULARA - SANGE-A - 2022 CopieDocument68 paginiCurs - 1-2 - CELULARA - SANGE-A - 2022 CopieBianca RoşuÎncă nu există evaluări
- Transportul Membranar În CelulaDocument7 paginiTransportul Membranar În CelulaDragu OanaÎncă nu există evaluări
- Organizarea Moleculară A Membranei CelulareDocument13 paginiOrganizarea Moleculară A Membranei CelulareAdella MihaellaÎncă nu există evaluări
- Biocel Sub ExamDocument12 paginiBiocel Sub ExamAlexandra ConstantinescuÎncă nu există evaluări
- 1 2 Fiziologia-Membranei-CelDocument52 pagini1 2 Fiziologia-Membranei-CelantonioÎncă nu există evaluări
- Fiziologia Membranei CelulareDocument58 paginiFiziologia Membranei CelulareJim BoyÎncă nu există evaluări
- C 3 An IDocument43 paginiC 3 An ITheodora IrinaÎncă nu există evaluări
- Membrana CelularaDocument77 paginiMembrana CelularaCătălina RaicuÎncă nu există evaluări
- Curs 4 BIOFIZDocument72 paginiCurs 4 BIOFIZfloricapopovici168Încă nu există evaluări
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Celula AnimalaDocument4 paginiCelula AnimalaAlexandra Vartolomei100% (3)
- Raspunsuri La Examen FiziologieDocument83 paginiRaspunsuri La Examen FiziologieDaniela IstratiÎncă nu există evaluări
- Curs 1 Membrana CelularaDocument40 paginiCurs 1 Membrana CelularaAnonymous NK07LsS9lÎncă nu există evaluări
- Citol 1Document37 paginiCitol 1jek_HÎncă nu există evaluări
- CelulaDocument4 paginiCelulaCrAzYMuSiCÎncă nu există evaluări
- Citologie MembranaDocument33 paginiCitologie MembranaOsman Daniela100% (1)
- Cursul 5 - Semnalizarea Celulara 2018-2019 PDFDocument31 paginiCursul 5 - Semnalizarea Celulara 2018-2019 PDFIrinush PetroaiaÎncă nu există evaluări
- Anatomie Curs 1Document16 paginiAnatomie Curs 1Danut Ancuța MihaelaÎncă nu există evaluări
- 1.fiziologia Nervilor Si MuschilorDocument22 pagini1.fiziologia Nervilor Si MuschilorPS NIÎncă nu există evaluări
- Excitabilitatea Și Transportul Ionic Membranar: Curs 2Document68 paginiExcitabilitatea Și Transportul Ionic Membranar: Curs 2Gavrilescu AlexandraÎncă nu există evaluări
- 04 Structura NeuronuluiDocument5 pagini04 Structura NeuronuluirodiÎncă nu există evaluări
- Fiziologia Membranei CelulareDocument29 paginiFiziologia Membranei CelulareCondrea100% (1)
- Biomecanica CeluleiDocument24 paginiBiomecanica CeluleiCristina ElenaÎncă nu există evaluări
- C6 BiofizicaDocument31 paginiC6 BiofizicaSimona PîslaruÎncă nu există evaluări
- CELULADocument4 paginiCELULAMadalina TrandafirÎncă nu există evaluări
- HISTOLOGIE 1 - Curs 3 - 2022Document16 paginiHISTOLOGIE 1 - Curs 3 - 2022Andreea IlincaÎncă nu există evaluări
- Tema 1Document25 paginiTema 1Bunici MarinelaÎncă nu există evaluări
- ColinomimeticeDocument1 paginăColinomimeticeLoredanaÎncă nu există evaluări
- Reactiile Serologice09Document84 paginiReactiile Serologice09VLDs92% (13)
- Examinarea Clinică in Afecţiuni CardiaceDocument38 paginiExaminarea Clinică in Afecţiuni CardiaceLucica Porsega VacarciucÎncă nu există evaluări
- CalamitatiDocument4 paginiCalamitatiBunici MarinelaÎncă nu există evaluări
- Intrebari Anul III 2021-22 MG RomDocument4 paginiIntrebari Anul III 2021-22 MG RomInna995Încă nu există evaluări
- 03 - CV - Curs 2 - AscultatiaDocument49 pagini03 - CV - Curs 2 - AscultatiaOlga Cretu HaidauÎncă nu există evaluări
- AdrenergiceDocument1 paginăAdrenergiceLoredanaÎncă nu există evaluări
- 4 5. Deregl. Circ. Sang. RegionaleDocument113 pagini4 5. Deregl. Circ. Sang. RegionaleBunici MarinelaÎncă nu există evaluări
- Radioscopia. DefiniţieDocument7 paginiRadioscopia. DefiniţieBunici MarinelaÎncă nu există evaluări
- VIDocument5 paginiVIBunici MarinelaÎncă nu există evaluări
- Diagnostic SpirochetozeDocument13 paginiDiagnostic SpirochetozeАнастасия СиминенкоÎncă nu există evaluări
- Tema 6Document14 paginiTema 6Bunici MarinelaÎncă nu există evaluări
- Raportul Organelor Cu PeritoneulDocument19 paginiRaportul Organelor Cu PeritoneulBunici MarinelaÎncă nu există evaluări
- Tipurile ComunicariiDocument2 paginiTipurile ComunicariiBunici MarinelaÎncă nu există evaluări
- Anatomia Toopografica A Peretelui Anterolateral Al AbdomenuluiDocument12 paginiAnatomia Toopografica A Peretelui Anterolateral Al AbdomenuluiBunici MarinelaÎncă nu există evaluări
- 2Document3 pagini2Bunici MarinelaÎncă nu există evaluări
- Comunicarea În Medicină Ca Baza Relației Medic - PacientDocument4 paginiComunicarea În Medicină Ca Baza Relației Medic - PacientBunici MarinelaÎncă nu există evaluări
- Tema 1 1 Fizologie MED RO-17402Document5 paginiTema 1 1 Fizologie MED RO-17402Bunici MarinelaÎncă nu există evaluări
- Reguli Pentru Elaborarea Lucrului IndividualDocument1 paginăReguli Pentru Elaborarea Lucrului IndividualBunici MarinelaÎncă nu există evaluări
- T 1Document26 paginiT 122194Încă nu există evaluări
- Mana Anatomia ClinicaDocument40 paginiMana Anatomia ClinicaAmbros SerafimaÎncă nu există evaluări
- Intrebari Pe Capitole Romana M 2015 PDFDocument13 paginiIntrebari Pe Capitole Romana M 2015 PDFAnonymous nA8QoFiW4Încă nu există evaluări
- Întrebări Stud An IV Chirurgie Tehnici Op. 2014Document3 paginiÎntrebări Stud An IV Chirurgie Tehnici Op. 2014Maria GorodencoÎncă nu există evaluări
- Tema 1 1 Fizologie MED RO-17402Document5 paginiTema 1 1 Fizologie MED RO-17402Bunici MarinelaÎncă nu există evaluări
- Potentialul de ActiuneDocument12 paginiPotentialul de ActiuneBunici MarinelaÎncă nu există evaluări
- Tema 4Document28 paginiTema 4Bunici Marinela0% (1)