S-ar putea să vă placă și
- Reactiile Serologice09Document84 paginiReactiile Serologice09VLDs92% (13)
- Biochimie Lucrari PracticeDocument8 paginiBiochimie Lucrari PracticeOctavian BotnaruÎncă nu există evaluări
- Biochimie Teste ExamenDocument221 paginiBiochimie Teste ExamenIspas Nina100% (3)
- Fizio TesteDocument73 paginiFizio TesteAdrea ElisaÎncă nu există evaluări
- Neurofiziologie MotorieDocument11 paginiNeurofiziologie MotorieAnonymous cKlFFm1eNÎncă nu există evaluări
- Colinoblocante 2017Document79 paginiColinoblocante 2017Tatiana Manoila100% (2)
- Tot 2 FizpatDocument52 paginiTot 2 FizpatRenata ProsutinskaiaÎncă nu există evaluări
- Totalizarea 2 GUDocument29 paginiTotalizarea 2 GUIulian StorojovÎncă nu există evaluări
- PanaritiileDocument30 paginiPanaritiiledr_vc100% (1)
- Raspunsuri La Examen FiziologieDocument83 paginiRaspunsuri La Examen FiziologieDaniela IstratiÎncă nu există evaluări
- Repere Totalizarea III BM Rom 2014Document7 paginiRepere Totalizarea III BM Rom 2014Irina CornilovÎncă nu există evaluări
- Tema .2 FizologieDocument6 paginiTema .2 FizologieSergiu Croitor100% (1)
- Ed. FizDocument2 paginiEd. FizIuliaÎncă nu există evaluări
- Totalizare Fiziologie1 1Document58 paginiTotalizare Fiziologie1 1Ana DubceacÎncă nu există evaluări
- Tema 1 (Fiziologia Senzoriala)Document9 paginiTema 1 (Fiziologia Senzoriala)Mihaela Botnari-Guţu100% (1)
- Itimi 2Document389 paginiItimi 2BeatriceÎncă nu există evaluări
- Nervul in Limbajul Clasic, Tripticul Mişcării EsteDocument10 paginiNervul in Limbajul Clasic, Tripticul Mişcării EsteDavidan20Încă nu există evaluări
- Teste Examen 20-21 (1) - 45805Document229 paginiTeste Examen 20-21 (1) - 45805Remmy Brawl Stars67% (3)
- BiofizicaDocument7 paginiBiofizicaGuțaÎncă nu există evaluări
- Embriolog 1Document16 paginiEmbriolog 1Elena PruniciÎncă nu există evaluări
- Transcriptia (1) .PpsDocument22 paginiTranscriptia (1) .PpsAna DubceacÎncă nu există evaluări
- Fiziologie Tot 2Document55 paginiFiziologie Tot 2Charlie McdonnellÎncă nu există evaluări
- Fiziologie 2Document11 paginiFiziologie 2Charlie McdonnellÎncă nu există evaluări
- Teste Cu Imagini Sem II 2017prima TotalizareDocument28 paginiTeste Cu Imagini Sem II 2017prima TotalizareAndr3 w00DÎncă nu există evaluări
- FIziologia Aparatului NeuromuscularDocument14 paginiFIziologia Aparatului NeuromuscularLoredana Boghez100% (1)
- Sistem MuschiDocument19 paginiSistem MuschiDan MassonÎncă nu există evaluări
- Tema 7. Metodele Clinico-Fiziologice de Cercetare A Activităţii CardiaceDocument6 paginiTema 7. Metodele Clinico-Fiziologice de Cercetare A Activităţii CardiaceAlexandra MagaleasÎncă nu există evaluări
- Întrebari Pentru Examen La Disciplina Endocrinologie Pentru Studenţii Anului 4Document103 paginiÎntrebari Pentru Examen La Disciplina Endocrinologie Pentru Studenţii Anului 4Radu GherbÎncă nu există evaluări
- Biochimie Lucrari PracticeDocument7 paginiBiochimie Lucrari PracticeOctavian BotnaruÎncă nu există evaluări
- Hormonii BiochimieDocument8 paginiHormonii BiochimieLoredanaÎncă nu există evaluări
- Anatomie Tot 3Document16 paginiAnatomie Tot 3Petru Dobrojan100% (1)
- Teste AnatomieDocument43 paginiTeste AnatomiemariutzaÎncă nu există evaluări
- Nervii FesieriDocument3 paginiNervii FesieriDan100% (1)
- Totalizare 3 Histologie Sime 3Document14 paginiTotalizare 3 Histologie Sime 3Иван БылбаÎncă nu există evaluări
- Lucru Individual Tot. 3Document80 paginiLucru Individual Tot. 3Dodushka WinstonÎncă nu există evaluări
- Calea PentozofosfațilorDocument7 paginiCalea PentozofosfațilorAlexandra MagaleasÎncă nu există evaluări
- Tema 3Document16 paginiTema 3Bunici MarinelaÎncă nu există evaluări
- Totalizarea Biofizica 3Document7 paginiTotalizarea Biofizica 3Dumitru SirghiÎncă nu există evaluări
- Fiziologie CM 1 156Document51 paginiFiziologie CM 1 156Oleg CojocariÎncă nu există evaluări
- Tema 4Document68 paginiTema 4Elena PavlicÎncă nu există evaluări
- TestDocument7 paginiTestOlaru Catalina100% (1)
- Fiziologie SNCDocument63 paginiFiziologie SNCMihaiCristianÎncă nu există evaluări
- Curs11 Anatomia Topografica Si Clinica A Peretilor ToraciciDocument5 paginiCurs11 Anatomia Topografica Si Clinica A Peretilor ToraciciSarikaya EmreÎncă nu există evaluări
- Farm Modific2Document3 paginiFarm Modific2Ilie CebanÎncă nu există evaluări
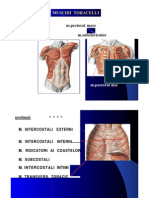
- Muschii ToraceluiDocument7 paginiMuschii ToraceluiBenny ScorpyoÎncă nu există evaluări
- Tema 12Document3 paginiTema 12Daniela LeonteÎncă nu există evaluări
- Anatomia Pe Viu Sem 3Document14 paginiAnatomia Pe Viu Sem 3Ladonna WadeÎncă nu există evaluări
- Anatomie Pe Viu Sem 1Document20 paginiAnatomie Pe Viu Sem 1Berladean Natalia100% (2)
- Afectarea Jonctiunii Neuro-MusculareDocument2 paginiAfectarea Jonctiunii Neuro-MuscularepzdtskblÎncă nu există evaluări
- 1anatomie 1 26Document21 pagini1anatomie 1 26ambulat100% (1)
- Plexul Cervical DocumentDocument2 paginiPlexul Cervical DocumentTeona MelinteÎncă nu există evaluări
- cursII PDFDocument12 paginicursII PDFPetru DariiÎncă nu există evaluări
- Tema 1. FizologieDocument7 paginiTema 1. FizologieSergiu Croitor100% (1)
- Biochimie Tot3Document29 paginiBiochimie Tot3Daniela CotoviciÎncă nu există evaluări
- Tesutul MuscularDocument10 paginiTesutul MuscularGiku MikushimirikuÎncă nu există evaluări
- Notiuni de Anatomie Si Fiziologie A Sistemului NervosDocument15 paginiNotiuni de Anatomie Si Fiziologie A Sistemului NervosMargarit MonicaÎncă nu există evaluări
- Îfiziologia Sist MuscDocument11 paginiÎfiziologia Sist MuscmalexandraÎncă nu există evaluări
- C 1 Sistemul NeuromuscularDocument67 paginiC 1 Sistemul NeuromuscularAlexandru MindruÎncă nu există evaluări
- Str. NeuronuluiDocument7 paginiStr. NeuronuluiLehaci Adelina100% (1)
- Curs 5 Anatomie - Sistemul MusularDocument10 paginiCurs 5 Anatomie - Sistemul MusularAlina AliÎncă nu există evaluări
- Sistemul MuscularDocument4 paginiSistemul MuscularAndreea GÎncă nu există evaluări
- NeuromuscularDocument13 paginiNeuromuscularBogdan Viorel VilceleanuÎncă nu există evaluări
- Tema 1Document25 paginiTema 1Bunici MarinelaÎncă nu există evaluări
- ColinomimeticeDocument1 paginăColinomimeticeLoredanaÎncă nu există evaluări
- Examinarea Clinică in Afecţiuni CardiaceDocument38 paginiExaminarea Clinică in Afecţiuni CardiaceLucica Porsega VacarciucÎncă nu există evaluări
- CalamitatiDocument4 paginiCalamitatiBunici MarinelaÎncă nu există evaluări
- Intrebari Anul III 2021-22 MG RomDocument4 paginiIntrebari Anul III 2021-22 MG RomInna995Încă nu există evaluări
- 03 - CV - Curs 2 - AscultatiaDocument49 pagini03 - CV - Curs 2 - AscultatiaOlga Cretu HaidauÎncă nu există evaluări
- AdrenergiceDocument1 paginăAdrenergiceLoredanaÎncă nu există evaluări
- 4 5. Deregl. Circ. Sang. RegionaleDocument113 pagini4 5. Deregl. Circ. Sang. RegionaleBunici MarinelaÎncă nu există evaluări
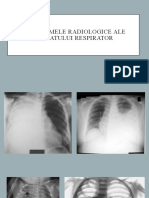
- Radioscopia. DefiniţieDocument7 paginiRadioscopia. DefiniţieBunici MarinelaÎncă nu există evaluări
- VIDocument5 paginiVIBunici MarinelaÎncă nu există evaluări
- Diagnostic SpirochetozeDocument13 paginiDiagnostic SpirochetozeАнастасия СиминенкоÎncă nu există evaluări
- Tema 6Document14 paginiTema 6Bunici MarinelaÎncă nu există evaluări
- Raportul Organelor Cu PeritoneulDocument19 paginiRaportul Organelor Cu PeritoneulBunici MarinelaÎncă nu există evaluări
- Tipurile ComunicariiDocument2 paginiTipurile ComunicariiBunici MarinelaÎncă nu există evaluări
- Anatomia Toopografica A Peretelui Anterolateral Al AbdomenuluiDocument12 paginiAnatomia Toopografica A Peretelui Anterolateral Al AbdomenuluiBunici MarinelaÎncă nu există evaluări
- Totalizarea 2Document43 paginiTotalizarea 2Bunici MarinelaÎncă nu există evaluări
- 2Document3 pagini2Bunici MarinelaÎncă nu există evaluări
- Comunicarea În Medicină Ca Baza Relației Medic - PacientDocument4 paginiComunicarea În Medicină Ca Baza Relației Medic - PacientBunici MarinelaÎncă nu există evaluări
- Tema 1 1 Fizologie MED RO-17402Document5 paginiTema 1 1 Fizologie MED RO-17402Bunici MarinelaÎncă nu există evaluări
- Reguli Pentru Elaborarea Lucrului IndividualDocument1 paginăReguli Pentru Elaborarea Lucrului IndividualBunici MarinelaÎncă nu există evaluări
- T 1Document26 paginiT 122194Încă nu există evaluări
- Mana Anatomia ClinicaDocument40 paginiMana Anatomia ClinicaAmbros SerafimaÎncă nu există evaluări
- Tema 1 1 Fizologie MED RO-17402Document5 paginiTema 1 1 Fizologie MED RO-17402Bunici MarinelaÎncă nu există evaluări
- Întrebări Stud An IV Chirurgie Tehnici Op. 2014Document3 paginiÎntrebări Stud An IV Chirurgie Tehnici Op. 2014Maria GorodencoÎncă nu există evaluări
- Tesut Exitabil 2020-16957Document115 paginiTesut Exitabil 2020-16957Bunici MarinelaÎncă nu există evaluări
- Intrebari Pe Capitole Romana M 2015 PDFDocument13 paginiIntrebari Pe Capitole Romana M 2015 PDFAnonymous nA8QoFiW4Încă nu există evaluări
- Potentialul de ActiuneDocument12 paginiPotentialul de ActiuneBunici MarinelaÎncă nu există evaluări