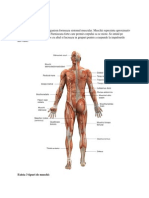
S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Neurofiziologie MotorieDocument11 paginiNeurofiziologie MotorieAnonymous cKlFFm1eNÎncă nu există evaluări
- Sistemul NervosDocument22 paginiSistemul NervosVlada ChiriacÎncă nu există evaluări
- Kinetologie 4Document28 paginiKinetologie 4Aland MediaÎncă nu există evaluări
- C13-14 Arc Reflex Elementar Receptori Musculari Analizator Kinestezic Arcul Reflex Elementar Arcul Reflex ElementarDocument18 paginiC13-14 Arc Reflex Elementar Receptori Musculari Analizator Kinestezic Arcul Reflex Elementar Arcul Reflex ElementarMarius RusuÎncă nu există evaluări
- Subiecte Fizio LP - RezolvateDocument29 paginiSubiecte Fizio LP - RezolvateDaniela DulgheruÎncă nu există evaluări
- Bazele KinetoDocument23 paginiBazele KinetoMarius MolanÎncă nu există evaluări
- Curs Metode Si Tehnici de Reeducare Neuromotorie ManoleLDocument98 paginiCurs Metode Si Tehnici de Reeducare Neuromotorie ManoleLFlorin Sbs92% (12)
- Reflex eDocument8 paginiReflex eBogdanÎncă nu există evaluări
- Curs Capitolul VIII Tema 1 2 Neurofiziologie Motorie Și Integrativă - 48797Document171 paginiCurs Capitolul VIII Tema 1 2 Neurofiziologie Motorie Și Integrativă - 48797Catalina EsanuÎncă nu există evaluări
- Reflex eDocument10 paginiReflex eMadalina MarinÎncă nu există evaluări
- Tema 4Document28 paginiTema 4Bunici Marinela0% (1)
- MotilitateaDocument17 paginiMotilitateaCocosul Cocosului CocosaruluiÎncă nu există evaluări
- Studiul MotilitățiiDocument18 paginiStudiul MotilitățiiAlexandra BuÎncă nu există evaluări
- Subiecte Fizio LPDocument27 paginiSubiecte Fizio LPhubertsÎncă nu există evaluări
- Analizatorul KinestezicDocument3 paginiAnalizatorul KinestezicBianca BrlcÎncă nu există evaluări
- SNC Tema 1Document7 paginiSNC Tema 1Pirdun JambonÎncă nu există evaluări
- Functiile Motorii Ale Maduvei Spinarii Reflexele Medulare. Controlul Functiilor Motorii Realizat de Cortex Si Trunchiul Cerebral.Document7 paginiFunctiile Motorii Ale Maduvei Spinarii Reflexele Medulare. Controlul Functiilor Motorii Realizat de Cortex Si Trunchiul Cerebral.Mihaela Botnari-Guţu100% (4)
- C 1 Sistemul NeuromuscularDocument67 paginiC 1 Sistemul NeuromuscularAlexandru MindruÎncă nu există evaluări
- Tehnici FNP BobathDocument28 paginiTehnici FNP BobathGigiEnergieÎncă nu există evaluări
- Analizator KinestezicDocument2 paginiAnalizator KinestezicRemus Coman100% (1)
- Analizatorul KinestezicDocument25 paginiAnalizatorul KinestezicEne Violeta Alina100% (1)
- Analizatorul Kinestezic: Nu Sunt Proprioreceptori! Sunt Numai Receptori DureroșiDocument3 paginiAnalizatorul Kinestezic: Nu Sunt Proprioreceptori! Sunt Numai Receptori DureroșiProfu' de biologieÎncă nu există evaluări
- Fiziologia Sistemul NervosDocument27 paginiFiziologia Sistemul NervosAurelia HayateÎncă nu există evaluări
- Analizatorul KinestezicDocument16 paginiAnalizatorul KinestezicRamona AndreeaÎncă nu există evaluări
- Kineto 2 Miscarea VoluntaraDocument40 paginiKineto 2 Miscarea VoluntaraJojo MikÎncă nu există evaluări
- Sistem MuschiDocument19 paginiSistem MuschiDan MassonÎncă nu există evaluări
- Tonus MGDocument27 paginiTonus MGCarp TheodorÎncă nu există evaluări
- Reflexe OsteotendinoaseDocument9 paginiReflexe Osteotendinoasedfgjrysertyufghr56533Încă nu există evaluări
- Anatomie 4 Curs 1Document3 paginiAnatomie 4 Curs 1AlexÎncă nu există evaluări
- Arc Reflex ElementarDocument8 paginiArc Reflex ElementarRenata ProsutinskaiaÎncă nu există evaluări
- Tesuturi Excitabile IIIDocument5 paginiTesuturi Excitabile IIINirca VadimÎncă nu există evaluări
- Fiziologia Aparatului Neuromuscular, SenzorialDocument18 paginiFiziologia Aparatului Neuromuscular, SenzorialAna Buga Lapteacru0% (1)
- Muschi IDocument5 paginiMuschi IAlex CosminÎncă nu există evaluări
- MotoriiDocument4 paginiMotoriiValentin BostanÎncă nu există evaluări
- Curs 5 Anatomie - Sistemul MusularDocument10 paginiCurs 5 Anatomie - Sistemul MusularAlina AliÎncă nu există evaluări
- Sistemul NervosDocument20 paginiSistemul NervosAlina AsaftiÎncă nu există evaluări
- Arcul Reflex ElementarDocument4 paginiArcul Reflex ElementarCalin CalinÎncă nu există evaluări
- Sistemul MuscularDocument4 paginiSistemul MuscularAndreea GÎncă nu există evaluări
- Analizatorul Kinestezic: o Corpusculii Neurotendinoși GolgiDocument4 paginiAnalizatorul Kinestezic: o Corpusculii Neurotendinoși GolgiProfu' de biologieÎncă nu există evaluări
- Curs 12 Analizatorul KinestezicDocument7 paginiCurs 12 Analizatorul Kinestezicaminalhassan963Încă nu există evaluări
- Curs 3.2. Tonus MuscularDocument35 paginiCurs 3.2. Tonus MuscularMateescu Cristian100% (2)
- C SN Somatic FRDocument15 paginiC SN Somatic FRSorin PalamariÎncă nu există evaluări
- Tema 1Document25 paginiTema 1Bunici MarinelaÎncă nu există evaluări
- Caracteristicile Generale Ale Ţesutului Muscular PDFDocument14 paginiCaracteristicile Generale Ale Ţesutului Muscular PDFsgaftonianuÎncă nu există evaluări
- Notiuni de Anatomie Si Fiziologie A Sistemului NervosDocument15 paginiNotiuni de Anatomie Si Fiziologie A Sistemului NervosMargarit MonicaÎncă nu există evaluări
- Curs 2 KinesiologieDocument43 paginiCurs 2 KinesiologieCrissu CrissÎncă nu există evaluări
- Sunetul fizicii: Acustica fenomenologicăDe la EverandSunetul fizicii: Acustica fenomenologicăEvaluare: 4.5 din 5 stele4.5/5 (2)
- NEUROPSIHOLOGIE: elementele de bază ale problemeiDe la EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiEvaluare: 5 din 5 stele5/5 (1)
- PrbsDocument1 paginăPrbsBogdan Viorel VilceleanuÎncă nu există evaluări
- Farmaco CursDocument8 paginiFarmaco CursBogdan Viorel VilceleanuÎncă nu există evaluări
- Virusologie Curs - Caractere GeneralDocument5 paginiVirusologie Curs - Caractere GeneralBogdan Viorel VilceleanuÎncă nu există evaluări
- Metodologie Concurs Erasmus 2019 Facultatea de MedicinaDocument4 paginiMetodologie Concurs Erasmus 2019 Facultatea de MedicinaBogdan Viorel VilceleanuÎncă nu există evaluări
- Diagnosticul de LaboratorDocument2 paginiDiagnosticul de LaboratorBogdan Viorel VilceleanuÎncă nu există evaluări
- Farmacologie LPDocument32 paginiFarmacologie LPBogdan Viorel Vilceleanu100% (1)
- Anatolp 1Document3 paginiAnatolp 1Bogdan Viorel VilceleanuÎncă nu există evaluări
- Farmaco CursDocument8 paginiFarmaco CursBogdan Viorel VilceleanuÎncă nu există evaluări
- Cardiologie 2017 PDFDocument18 paginiCardiologie 2017 PDFMădălina IuraşcuÎncă nu există evaluări
- HemostazaDocument27 paginiHemostazaBogdan Viorel VilceleanuÎncă nu există evaluări
- HahhahhaDocument3 paginiHahhahhaTanaseÎncă nu există evaluări
- Curs R-A, An LL, Sem.l Edit Curs Radioanatomie An 2 Semestrul 1Document119 paginiCurs R-A, An LL, Sem.l Edit Curs Radioanatomie An 2 Semestrul 1Raiiad MihaÎncă nu există evaluări
- FPAT LPDocument6 paginiFPAT LPBogdan Viorel VilceleanuÎncă nu există evaluări
- Anato II, II-Partea 1Document13 paginiAnato II, II-Partea 1georgymÎncă nu există evaluări
- LP Embrio 2Document4 paginiLP Embrio 2Bogdan Viorel VilceleanuÎncă nu există evaluări
- Ghidul Voluntarului - EduMathDocument4 paginiGhidul Voluntarului - EduMathBogdan Viorel VilceleanuÎncă nu există evaluări
- TalamusDocument7 paginiTalamusBogdan Viorel VilceleanuÎncă nu există evaluări
- Embrio LP An 1 Sem 2Document9 paginiEmbrio LP An 1 Sem 2Bogdan Viorel VilceleanuÎncă nu există evaluări
- Efectele Metabolice Ale Hormonilor TiroidieniDocument13 paginiEfectele Metabolice Ale Hormonilor TiroidieniBogdan Viorel VilceleanuÎncă nu există evaluări
- Fisa Inscriere Admitere Licenta Istorie - OkDocument2 paginiFisa Inscriere Admitere Licenta Istorie - OkDia DiduÎncă nu există evaluări
- Graficul Orientativ Al Competitiilor Din Domeniul Fizicii 2016-2017Document1 paginăGraficul Orientativ Al Competitiilor Din Domeniul Fizicii 2016-2017Bogdan Viorel VilceleanuÎncă nu există evaluări
- Anestezia GeneralaDocument7 paginiAnestezia GeneralaLica Ciocan100% (1)
- ArticulatiiDocument1 paginăArticulatiiBogdan Viorel VilceleanuÎncă nu există evaluări
- Orar I II IstorieDocument7 paginiOrar I II IstorieBogdan Viorel VilceleanuÎncă nu există evaluări
- Anato - II, II Partea 2Document15 paginiAnato - II, II Partea 2Marius CarligÎncă nu există evaluări
- Barem CineticăDocument2 paginiBarem CineticăBogdan Viorel VilceleanuÎncă nu există evaluări
- Interferon de FaraoniDocument1 paginăInterferon de FaraoniBogdan Viorel VilceleanuÎncă nu există evaluări
- Bazinul LondreiDocument2 paginiBazinul LondreiBogdan Viorel Vilceleanu100% (1)
- Tema Clubul Matematicienilor P - 162-172Document11 paginiTema Clubul Matematicienilor P - 162-172Bogdan Viorel VilceleanuÎncă nu există evaluări