Documente Academic
Documente Profesional
Documente Cultură
Sistemul Oro - Facial
Sistemul Oro - Facial
Încărcat de
Anca CiutacuDrepturi de autor
Formate disponibile
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentDrepturi de autor:
Formate disponibile
Sistemul Oro - Facial
Sistemul Oro - Facial
Încărcat de
Anca CiutacuDrepturi de autor:
Formate disponibile
1
INTRODUCERE
Sistemul oro-facial numit i aparatul dento-maxilar (Bratu), sistemul stomatognat (Burlui) cuprinde totalitatea esuturilor i organelor cu o mare varietate structural i funcii difereniate, specifice ce concur n final la realizarea unor procese fiziologice de importan vital. Morfologic, putem diferenia sistemul oro-facial n: 1. ansamblul osos care este suportul pasiv de susinere i rezisten reprezentat de cele dou maxilare, articulaiile temporomandibulare, dinii. Acestea realizeaz att funciile proprii, individuale, ct i funciile globale ale sistemului; 2 ansamblul muscular fiind suportul activ al generrii forelor n regiunea oro-facial; 3. prile moi oro-faciale, limba, glandele salivare, parodoniu, vasele sanguine, limfatice, nervii, ce integreaz ntr-un tot unitar ansamblul structurilor dento-maxilare. Caracteristicile principale ale sistemului oro-facial: este un sistem biologic complex i de aceea l putem considera, de fapt, un biosistem n care se difereniaz mai multe subsisteme: mediul bucal, dini, parodoniu, muchi, etc; este un sistem biologic deschis, care face n permanen schimburi cu mediul nconjurtor; este un sistem integral: fiecare element al sistemului oro-facial este o component necesar, dar nu suficient pentru funcia sistemului; este un sistem integrat organismului uman cu care stabilete corelaii privind att funcia specific subsistemului, ct i funciile de ansamblu ale sistemului oro-facial; este un sistem intercondiionat, ntre componentele sistemului existnd circuite informaionale care fac posibil funcionarea coerent, logic a ntregului sistem;
2 este un sistem biologic automat de reglare asigurnd att homeostazia specific sistemului, ct i homeostazia ansamblului organismului. Procesele de autoreglare specifice sistemelor automate cu circuit deschis sau nchis (feed-back) sunt dinamice, adaptndu-se ncontinuu nevoilor locale i generale ale oragnismului este un sistem eficient de aprare, care prin mijloacele specifice locale iate cu cele generale, lupt n permanen mpotriva agresiunii bacteriene, n cipal. "Fronturile de lupt" i aprare sunt localizate la nivelul: gingiei, ului gingival, fosetelor, fisurilor suprafeelor ocluzale ale dinilor, suprafelor bucale ale dinilor, parodoniului, mucoasei bucale, etc. Sistemul oro-facial ndeplinete funcii eseniale vieii: funcia digestiv prin secreia salivar, masticaie, deglutiie; funcia respiratorie, asigurnd pasajul bidirecional al aerului, curirea, nclzirea, umectarea aerului; funcia de comunicare interuman i interspecii prin: fonaie, vorbire, fizionomie, etc; funcia de aprare prin integritatea structurilor i prin componenii iului bucal (IgA secretorie, lizozim, leucocite, etc); funcia de autontreinere, adic de meninere constant a unor metri structurali, umorali, chimici, nervoi indispensabili pentru realizarea funciilor proprii ale sistemului oro-facial.
FIZIOLOGIA MEDIULUI BUCAL
Mediul bucal este un ecosistem fizico-chimic care ocup i influeneaz structurile cavitii bucale. Spre deosebire de mediul intern, mediul bucal este deschis n dou direcii (buze, faringe). Spre exterior prin cavitatea bucal i spre interiorul organismului prin intermediul aparatului digestiv. Mediul bucal cuprinde:
3 elemente de tranziie: alimente, aer; elemente proprii, provizorii: saliva i lichidul crevicular; flora microbian mobil i fix (din placa dentar, tartrul dentar, saliv, etc); componentele proprii, fixe: dinii, gingia, limba, mucoasa bucal. Toi factorii mediului bucal sunt n interaciune unii cu alii, iar fluctuaiile compoziiei mediului au repercursiuni directe asupra componentelor fixe. Cavitatea bucal este un sistem natural deschis. Proprietile biologice ale sistemului cavitii bucale sunt n permanent modificare datorit factorilor de mediu, n particular, prin preluarea alimentelor, hranei, ca i prin contaminarea cu microorganisme. De asemenea, structurile proprii cavitii bucale influeneaz mediul bucal prin modificrile diurne ale fluxului salivar, componenilor salivei. Cavitatea bucal poate fi comparat cu o camer de fermentaie, care asigur mai ales noaptea un mediu de cultur pentru microbi. Constituenii salivari i resturile alimentare restante, n mediul bucal, reprezint o surs permanent de substrat proaspt, de care beneficiaz microorganismele. Mediul bucal este un ecosistem cu proprieti fizice, chimice i biologice specifice, care determin compoziia comunitii bacteriene i condiioneaz microorganismele care domin sistemul i care lupt pentru supravieuire. Cavitatea bucal este o singur entitate, dar didactic poate fi mprit n alte mici ecosisteme independente i totui interdependente. Din punct de vedere fiziologic studiul complet al mediului bucal cuprinde numeroase aspecte privind: saliva, lichidul crevicular, flora bacterian bucal, placa dentar, tartrul dentar, alimentaia, ca i interrelaia acestora cu structurile proprii ale cavitii bucale, cu sistemele de aprare locale i generale.
2.1. FIZIOLOGIA GLANDELOR SALIVARE
Saliva este produsul de secreie a trei perechi de glande mari, situate n grosimea pereilor cavitii bucale (glande parotide, submaxilare, sublinguale) i a numeroase glande mici, diseminate n mucoasa care tapeteaz cavitatea bucal. Prin canalele de excreie saliva se vars n cavitatea bucal:
4 canalul STENON al glandei parotide (gland seroas) se deschide n vestibulul bucal n dreptul celui de al doilea molar superior; canalul WARTHON al glandei submandibulare (gland mixt) se deschide n cavitatea bucal lng frul limbii; canalul BARTHOLIN din sublingual (gland mixt) se deschide aproape de canalul WARTHON, lng frul limbii; canalele accesorii RIVINIUS din sublingual se deschid n cavitatea bucal sub limb n partea anterioar. 2.1.1. Structura morfo-funcional a glandelor salivare Glandele salivare au o structur tubulo-acinoas, fiind alctuite din lobuli ce reprezint unitatea morfo-funcional. Fiecare lobul este alctuit din acini i ducte intercalate, ducte striate. Din unirea mai multor ducte lobulare se formeaz ductul excretor principal. Acinul este alctuit din celule poligonale ce delimiteaz un spaiu: lumenul acinului. La exterior, acinul este nconjurat de o membran bazal. ntre membrana bazal i celulele acinare se gsesc celule mioepiteliale n citoplasm crora exist elemente contractile actomiozinice implicate n: expulzarea produsului de secreie glandular i n creterea presiunii intraluminale. Celulele mioepiteliale sunt controlate de inervaie simpatic. Celulele acinare secretoare sunt de dou tipuri: seroase i mucoase. Celulele seroase conin numeroase mitocondrii, reticul endoplasmatic rugos bine dezvoltat, granulaii mici ce conin zimogen. Membrana luminal are microvilovilozitati, iar poriunea bazo-lateral a membranei este cu numeroase pliuri i jonciuni intercelulare strnse. Permeabilitatea acestor jonciuni intercelulare crete mult dup stimulare asigurnd trecerea din interstiiu n lumen a: Na+, K+, apei. Pe membrana luminal se gsete adenilat-ciclaz (asociat receptorilor beta-adrenergici i vipergici) iar n poriunea bazo-lateral exist receptori colinergici, alfa-adrenergici, activiti enzimatice: Na+-K+-ATP-az, anhidraz carbonic, etc. Celulele mucoase conin granulaii mari cu aspect vacuolar, reticul endoplasmatic slab dezvoltat, secret mucus. Acinul particip la formarea salivei primare. Ductul intercalat este alctuit din celule mici, cuboidale, au ule de secreie, membrana bazo-lateral este uor plicaturat, are activiti ATPazice. Ductul striat are celule nalte, mari, nucleu central i membran bazolateral plicaturat (pliurile mresc suprafaa membranei), numeroase mitocondrii (furnizeaz energia necesar proceselor de transport), granule (rol n secreia de proteine). Ducte excretorii interlobulare au epiteliul bistratificat, celulele nalte prezint de asemenea pliuri ale mucoasei bazo-laterale. Epiteliul ductal secret: kalicrein, amilaz, are activiti enzimatice: anhidraz carbonic (particip la formarea bicarbonatului), prostaglandinsintetaz (asigur sinteza de prostaglandine prin care controleaz absorbia ductal a electroliilor), 5 alfa reductaz (determin
5 reducerea androgenilor n metabolii mai activi fiziologici). Sistemul ductal particip la formarea salivei finale, definitive, prin procese de reabsorbie i secreie. 2.1.2. Vascularizaia glandelor salivare Glandele parotide sunt irigate de ramuri arteriale provenite din artera facial i carotida extern. Glandele submandibulare primesc ramuri din artera facial i lingual, iar glandele sublinguale din artera sublingual i submental. Vasele se distribuie n glande mergnd paralel cu ductele principale, terminaiile nervoase. Se realizeaz un sistem "contra curent" ntre sensul de deplasare al sngelui i salivei. O alt particularitate este faptul c arteriolele se capilarizeaz nti la nivelul ductelor, dup care realizeaz o a doua capilarizare la nivelul acinilor realiznd un sistem port. Irigaia duetelor este mai bogat dect a acinilor. Sistemul venos al glandelor salivare se colecteaz n vena jugular. Debitul salivar este dependent de debitul circulator sanguin, care crete de cinci ori n cursul salivaiei. Vasomotricitatea n glandele salivare depinde de: inervaia simpatic, parasimpatic, factori umorali. Inervaia simpatic din glandele salivare produce vasodilataie prin aciunea adrenalinei asupra receptorilor beta-adrenergici. Noradrenalina, ca i n alte teritorii, contract vasele din glandele salivare, dar indirect poate produce vasodilataie deoarece stimuleaz, prin intermediul alfareceptorilor, activitatea kalicreinogenului i formarea de bradikinin. Inervaia parasimpatic prin acetilcolin produce vasodilataie i stimuleaz secreia salivar. Factorii umorali: bradikinina are efect vasodilatator de lung durat. Kalicreina, prezent n celulele acinare i ductale, acioneaz asupra kalicreinogenului formnd kalidin apoi bradikinin. Metaboliii: CO2, H+, rezultai din activitatea glandular, au efect vasodilatator local.
Fig.l Stimularea glandei parotide (dup PATTON, 1989, modificat) Poteniale membranare n glandele salivare Cu ajutorul microelectrozilor s-a stabilit c potenialul de repaus la nivelul delor acinare, pe membrana bazo-lateral, variaz n funcie de gland n limite largi de la -30mV la -73mV, iar la nivelul celulelor ductelor striate este de -80mV pn la -90mV. Potenialul de membran este meninut la aceste valori prin participarea pompei Na + - K + canalelor de K +, CI- voltaj dependente. Permeabilitatea membranei este dominat de K+ (116mM intracelular i 3,3mM extracelular). n repaus, n glandele salivare conductanele ionice sunt: GCI > GK >GNa. Stimularea receptorilor colinergici, alfa-adrenergici produce depolarizarea membranei prin creterea conductanei pentru Na+, care intr n celul i concomitent are loc un eflux de K+, Cl-. Se genereaz un potenial de aciune numit "potenial secretor". Depolarizarea favorizeaz, i intrarea Ca++, din lichidul extracelular n celula, care va participa la mecanismul de cuplare excitaie-secreie. 2.1.4. Cuplarea excitaie-secreie n glandele salivare Substanele neuro-umorale agoniste se fixeaz pe receptorii de la
7 suprafaa celulei inducnd diferii mesageri secunzi intracelular responsabili de secreia proteic i lichidian. Receptorii de pe glandele salivare sunt: colinergici (de tip muscarinic), adrenergici (alfa i beta), dopaminergici, purinergici, peptidergici pentru VIP i substana P). Stimularea acestor receptori determin fie creterea AMPc fie a C++ citosolic, ambele controlnd att secreia proteic, enzimatic ct si lichidian din glandele salivare. Deci rspunsul secretor poate fi AMPc dependent sau/i Ca++ dependent. (Fig.l) 2.1.4.1. Rspunsul secretor dependent de AMPc AMPc, ca i n alte esuturi, apare ca mesager secund al rspunsului mediat beta-adrenergic. Fixarea adrenalinei pe receptorul beta-adrenergic, cuplat cu proteina Gs, activeaz adenilatciclaza care scindeaz ATP n AMPc. AMPc activeaz proteinkinaza A care determin fosforilarea de proteine i sinteza de noi protein-enzime. Creterea activitii proteinkinazei A ctre AMPc, precede secreia de amilaz. Tolbutamina inhib proteinkinaza - AMPc dependent i astfel inhib secreia de amilaz. Stimularea receptorilor beta-adrenergici, administrarea de toxin holeric determinain secreia de amilaz prin creterea AMPc. Teofilina este un inhibitor al diesterazei (care inactiveaz AMPc) i indirect menine crescut AMPc intracelular, astfel este un activator eficient al secreiei de amilaz. 2.1.4.2. Rspunsul secretor Ca++ dependent Stimularea receptorilor muscarinici (de ctre acetilcolin) i alfa adrenergici (de ctre noradrenalin) induce n glandele salivare rspunsuri dependente de Ca++, n particular secreia de electrolii, lichide i proteinenzime. Ca++ intr din lichidul extracelular n celulele acinare prin canale cationice neselective din membrana bazo-lateral sau prin intermediul turnoverului inozitol-fosfolipidelor membranare.
Receptorii muscarinici i alfa-1-adrenergici stimulai, activeaz fosfolipaza C prin intermediul proteinei Gq. Fosfolipaza C desface fosfoinozil difosfat (PIP2) n diacilglicerol (DAG) i inozitol trifosfat (IP3). DAG activeaz proteinkinaza C care induce fosforilarea proteinelor ca i eliberarea de acid arahionic care poate crete rata produciei de GMPc. IP3 elibereaz Ca++ din reticulul sarcoplasmatic i favorizeaz intrarea Ca++ din lichidul extracelular n celula determinnd astfel creterea Ca++ citosolic. Aceasta are ca rezultat: legarea Ca++ de calmodulin, fosforilarea proteinelor, creterea permeabilitii jonctiunilor intercelulare, interaciunea direct a Ca++ cu efectori proteici ai canalelor de Ca++, K+, Cl-, activarea proteinkinazei C, activarea fosfolipazei A2 cu cresterea eliberrii de acid arahidonic (care crete rata produciei de GMPc).Atat creterea proteinkinazei C ct i a GMPc este implicat n stimularea secreiei de amilaz, mucin. Acetilcolina i substana P acioneaz direct pe receptorii celulelor ductale si mobilizeaz Ca++ intra, extracelular producnd stimularea secreiei salivare. VIP crete indirect secreia salivar prin creterea fluxului sanguin, secundar vasodilataiei pe care o determin. Se produce astfel o secreie salivar abundenta ca volum dar cu cantiti mici de proteine (enzime, mucin). 2.1.5. Mecanismul elaborrii salivei Saliva se formeaz n dou etape la care particip: acinii care elaboreaz saliva primar i apoi duetele salivare unde au loc procese de secreie i
9 reabsorbie care modific compoziia ionic a salivei. 2.1.5.1. Funciile acinului - Saliva primar rezult dintr-un transfer plasmatic prin membrana bazo-lateral i eliminare de saliv prin polul apical al celulelor acinare. Secreia primar conine amilaz i/sau mucin ntr-o soluie ionica cu concentraie asemntoare cu a lichidului extracelular. Secreia substanelor organice proteice la nivelul acinilor are mai multe etape ce cuprind: preluarea aminoacizilor prin membrana bazo-lateral, sinteza de peptide n reticulul endoplasmatic din partea bazal a acinului, migrarea i nglobarea n cisternele Golgi, condensarea i transformarea n granule de zimogen, evacuarea prin polul apical printr-un proces de exocitoz. Secreia proteic este dependent de prezena Ca++ n lichidul extracelular, iar cantitatea de granule exocitate depinde de intensitatea stimulrii. Saliva primar este izoton (290-310 mOsm/l) are o compoziie similar cu un ultrafiltrat plasmatic, dar concentraia K+ este mai mare dect n plasma, 1-15 mM/l, CI-= 100-120 mM/l, Na+ = 125-160 mM/l). In glandele salivare exist o bogat activitate ATP-azic mai ales pe membrana bazolateral. Na+-K+-ATPaza scoate 3 Na+ din celul n interstiiu duce 2 K+ n celul. Na+ din interstiiu trece n lumenul acinului prin pasaj paracelular de-a lungul jonciunilor membranei. (Fig.2)'
29
In urma activrii receptorilor colinergici sau alfa-1-adrenergici, IP3 determinan eliberarea Ca++ din reticulul sarcoplasmatic, procesul eliberrii implicnd ambele mecanisme: deschiderea canalelor de Ca++ IP3dependente dar i Ca++ eliberator de Ca++, n urma crora Ca++ difuzeaz n citosol. Consecutiv creterii Ca++ intracelular se activeaz canalele de K+ din membrana bazo-lateral i de CI- din membrana luminal rezultnd o pierdere net de K+, CI-, ap i micorarea volumului celular. n urma stimulrii, ieirea K+ din celulele acinare se face prin maxi canalele de K+, Ca++- dependente localizate pe membrana bazo-lateral, iar CI- iese prin canalele de Cl-, Ca++- dependente de pe membrana luminal. Concomitent cu ieirea CI-, iese i HCO-3 (prin canale sau prin schimb Cl - HCO-3) astfel c raportul dintre ei rmne nemodificat, intra-extra celular. Scderea HCO3 n celula acinar determin activarea anhidrazei carbonice care catalizeaz reacia: C02 + H20 <->H2C03 <-> HCO-3 + H+ rezultand o acidifiere acinar cu 0,1 uniti de pH. Ieirea CI- n lumen creeaz o diferen de potenial, mai negativ n lumen fa de interstiiu. Pentru pstrarea electroneutralitii extracelulare are loc o migrare a cationilor (5/6 K+ i 1/6 Na+) spre lumenul acinului pe cale paracelular, prin jonciuni cation selective. Ieirea K din celula n interstiiu, prin maxi canale de K+, asigur creterea sa n interstiu i posibilitatea ca el s migreze paracelular n lumen. Astfel saliva este bogat n CI-, Na+ dar i n K+. La cteva secunde dup stimulare, Ca++ intracelular scade i crete rapid Na intracelular datorit: activrii schimbului (antiport) Na+ - H+, activrii contrasportului (simport) Na+ - K+ - 2Cl-. Creterea sodiului n celul favorizeaza reintrarea apei cu revenirea celulei la volumul dinainte de stimulare i activarea pompei Na+ - K+ avnd ca rezultat creterea K+ n celul i a Na+ extracelular. Dup ieirea iniial a K+ i Cl-, are loc un reuptake prin simportul Na+ K+ - 2Cl- -Acidifierea acinar cauzat de ieirea HCO-3 i creterea metabolismului celular este contracarat de scoaterea H+ prin antiportul Na+ - H+. Alcalinizarea cu creterea HCO-3 intracelular determin din nou o cretere a Cl- prin antiportul CI- - HCO-3. Cnd concentraia intracelular a electroliilor, volumul celular i activitatea pompei Na+ - K+ a revenit la nivelul bazal, celulele acinare sunt din nou apte sa fie stimulate i s elaboreze o nou cantitate substanial de saliv Pe membrana luminal a acinului funcioneaz urmtoarele: canalele de Cl- , Ca++ - dependente prin care clorul iese din celula n lumen, crescnd aici electronegativitatea; antiport CI- - HCO-3 care elimin bicarbonatul n lumen; + - K+ - ATPaza care reintroduce o parte din K+ n celul. Na Pe membrana bazo-lateral a acinului au loc urmtoarele schimburi: simportul Na+ - K+- 2Cl- care introduce aceti ioni n celul;
29
pompa Na+ - K+ care introduce 2K+ din interstiiu i scoate 3Na+ din celul, care vor migra paracelular n lumen; maxi canale de K+, Ca++- dependente, prin care potasiul iese masiv n interstiiu ca apoi s ajung n lumen, pe cale paracelular; antiportul Cl- - HCO-3 care introduce clorul n celul i scoate bicarbonatul n interstiiu; antiportul Na+ - H+ care scoate ionul de H+ n interstiiu i preia de aici sodiul. 2.1.5.2 Funciile ductelor salivare Componenii organici ai salivei provin din: sinteza i secreia celulelor acinare (amilaz, mucin); transportul transepitelial la nivelul celulelor ductale (steroizii); sinteza i secreia celulelor ductale. Celulele ductale sintetizeaz, stocheaz i secret: factori de cretere (factor de cretere al nervilor, factori de cretere epidermali), enzime (ribonucleaz, amilaz), hormoni (glucagon, somatostatin, parotin, sialogastron), proteaze homeostatice (renin, kalicrein ce controleaz fluxul sanguin local, transportul apei i electroliilor). IgA secretorie este sintetizat i dimerizat de imunocitele interstiiale iar celulele ductale o preiau, i ataeaz componenta secretorie dup care o elibereaz n saliv. Piesa secretorie (GM-60000) este produs de celulele epiteliului ductal, este responsabil de pstrarea structurii cuaternare a IgA secretorie i i confer rezisten crescut la aciunile proteolitice. IgA secretorie asigur aprarea antimicrobian a cavitii bucale. Transportul ductal al apei i electroliilor: ionii de Na+ sunt reabsorbii activ din saliva ductal iar K+ este secretat activ. Concentraia sodiului n saliv depinde de fluxul salivar, crescnd o dat cu creterea fluxului salivar. Membrana luminal a ductelor are o conductan mare pentru Na+. Sodiul poate intra din lumen n celula ductal prin: antiportul Na+-H+, canale de Na+. Na+ intrat n celul iese n interstiiu prin pompa Na+-K+ existent pe membrana bazo-lateral, care asigur astfel scderea concentraiei Na+ n celul i creterea concentraiei K+ intracelular. Acesta iese n lumen prin antiport K+-H+ de pe membrana luminal. (Fig.3) Cl- din duct revine n celul fie prin canale de clor, fie prin antiport Cl-HCO-3. Concentraia bicarbonatului va fi mai mare sau mai mic dect n saliva primar, n funcie de secreia sau reabsorbia sa n ducte, dependent de echilibrul acido-bazic sau concentraia Na+ salivar.
29
Fig.3 Secreia i resorbia electroliilor n celulele ductale (dup JOHNSON, 1987, modificat) Pe membrana luminal a ductelor exist: canale de Na+, Cl- prin care acetia revin n celul; antiport Cl- - HCO-3 care introduce clorul i scoate bicarbonatul din celula antiportul Na+-H+ care economisete Na+ readucndu-l n celul; antiportul H+-K+ care scoate K+ n lumen. Pe membrana bazo-lateral a celulelor ductale au loc urmtoarele schimburi ionice: scoaterea 3Na+ i introducerea de 2K+ n celul de ctre pompa Na+-K+; canalele de K+ i Cl- prin care aceti ioni revin n interstiiu; antiportul Na+-H+ care scoate ionul de hidrogen din celul n interstiiu. Ductul salivar este puin permeabil pentru ap ceea ce asigur hipoosmolaritatea salivei finale. Rezultatele acestor schimburi transmembranare sunt:
29
concentraia Na+ i Cl- este de aproximativ 15 mEq/1 pentru fiecare, adic 1/7 - 1/10 din concentraia plasmatic; concentraia K+ este de aproximativ 30 mEq/1 adic de 6 ori mai mare dect n plasm; concentraia HCO3- este de 50-70 mEq/1 deci de 2-3 ori mai mare ca n plasm. ACTH i mineralcorticoizii scad Na+ salivar i cresc concentraia de K+. Aldosteronul crete activitatea antiportului luminai Na+- H+ i a pompei Na+ -K+ bazo-lateral. VIP i GIP n concentraie de 10-11 mol/l inhib transportul de Na+. Elaborarea salivei presupune un consum energetic de 6 Kcal pentru 1 litru de saliv format. Energia este asigurat att aerob ct i anaerob din: glucoza, glicogen, fosfocreatin. Metabolismul este asigurat de un debit sanguin corespunztor care are o valoare de 0,6ml/g.min. n glanda nestimulat, iar dup stimulare este de 4-5 ml/g.min. Stimularea secreiei glandulare determin creterea debitului sanguin de 4-8 ori, dublarea consumului de O2 i creterea temperaturii tisulare cu 1C. Cantitatea de saliv secretat n 24 de ore este 1-1,5 1 (1 ml/ min). n somn secreia salivar nu depete 0,25 ml/min. Evacuarea salivei n cavitatea bucal are loc prin: fora de mpingere vis-tergo a salivei secretate, contracia celulelor mioepiteliale, presarea glandelor de pereii osoi n timpul masticaiei. 2.1.6. Proprietile salivei 2.1.6.1. Volumul salivei este de aproximativ 1000-1500 ml/zi dar variaz n funcie de anumite stri, cum ar fi: repaus alimentar: 0,3-0,5 ml/min; n somn: 0,08 ml/min; n prin stimulare alimentar ajunge pn la 2-7 ml/min. Fluxul salivar depinde de stri fiziologice (vrst, greutate, sarcin, gradul de hidratare al organismului, momentul zilei, olfacie) i ali factori cum ar fi: fumat, medicaie, factori psihici (stres, gndul la alimente, vzul lor, aspectul lor). La copii, volumul salivar este diferenial : sugari: 50-100 ml/zi; la 5 ani: 0,22 ml/min. la Modificarea volumului salivar: hipersalivaia sau sialoreea (flux salivar peste 0,5 ml/min) se ntlnete fiziologic la erupia dinilor, reflexul salivar condiionat la stimul vizual, n sarcin, consum de condiment
35
Sialoreea patologic apare n stomatite, gingivite, intoxicaii cu Pb, Hg, ulcer duodenal, cancer gastric, patazitoze intestinale, vom, epilepsie, nevralgie de trigemen, consum de medicamente, anestezice. Tutunul produce hipersalivaie. hiposalivaia ( flux salivar cuprins ntre 0,01-0,06 ml/min). Fiziologic apare la btrni, la menopauz, sau datorit sentimentului de fric. Patologic, hiposalivaia apare n hemoragii, stri febrile, deshidratri severe, stomatite infectioase, la consum de atropin, antibiotice, opiacee. aptialismul (lipsa secreiei salivare, volum sub 0,01 ml/min) apare n stomatitele atrofice, degenerescenta glandelor salivare, intoxicaii cu atropin, morfin, opiu. 2.1.6.2. Aspectul salivei este opalescent, filant. 2.1.6.3. Sedimentul salivar cuprinde celule epiteliale descuamate, leucocite( ce trec prin diapedez din capilare n anul gingivo-dentar), bacterii (streptococ lactotobacillus acidophilus). Leucocitele se gsesc n special n saliva copiilor (inainte de erupia dinilor) i n saliva adulilor edentai. La persoanele cu gingii sanatoase, numrul de leucocite este relativ mic. Numrul leucocitelor prezint o variaie diurn, crescnd de 45 ori, dimineaa i n mijlocul zilei. Majoritatea leucocitelor salivare sunt dezintegrate, ceea ce permite eliberarea enzimelor leucocitare i mbogirea echipamentului enzimatic salivar. 2.1.6.4. pH-ul salivar In repaus pH-ul mediu salivar este de aproximativ 6,7 (5,2-7,6), prezentnd variaii n limite largi (5,6-8). pH-ul este sensibil la variaiile fluxului salivar . Dup stimularea glandelor pH-ul crete la valoarea de 8 datorit creterii bicarbonatului. pH-ul este crescut la copii, n hiperpnee, alcaloz. pH-ul scade n efort, n diabet zaharat. pH-ul salivar depinde de: a) concentraia de dioxid de carbon sanguin; Dac presiunea parial a dioxidului de carbon din snge scade, pH-ul salivar este mai alcalin; b) alimentaie (spanacul asigur pH mai alcalin); c) sisteme tampon salivare: - bicarbonat - acid carbonic (4,5/1), care se opune acidifierii mediului bucal. Acidul carbonic n saliv este n concentraie apropiat de cea din plasm (1,2 - 1,3 mM), depinznd de presiunea C02. Bicarconatul (HCO3-) se formeaz n glandele salivare din C02 i H20 rezultate din catabolismul glucozei. Celulele glandulare au enzima anhidraza carbonic, ce faciliteaz formarea formarea H2CO3 respectiv HC03-. C02 + H20 <-> H2CO3 <-> HC03- + H+. HCO3- este eliminat n saliv. Stimularea glandelor salivare determin creterea fluxului salivar, creterea producerii i eliminrii HCO3- n saliv
35
(concentraia HC03- crete de la 30 mM la 60mM). pH = pK + log (HCO3-) / ( H2C03) Creterea bicarbonatului n saliv produce creterea raportului bicarbonat/ acid carbonic, rezultnd creterea pH-ului spre 7,8. - fosfat disodic - fosfat monosodic, se opune alcalinizrii; - mucin bazic - mucin acid. Proteinele datorit caracterului lor amfoter, tamponeaz att acizii ct i bazele. d) prezena anhidrazei carbonice; e) fluxul salivar mrit duce la creterea pH-ului. Capacitatea tampon a salivei prezint mari variaii diurne: este mare dimineaa, imediat dup splarea dinilor, apoi scade repede; dup mas, crete n timp de o or ca apoi s scad de asemenea ntrun interval de o or; crete pn seara; somn, pH-ul salivei scade. n Osmolaritatea salivei mixte din cavitatea bucal este de 50-100 mOsm/1 (hipoton), punctul crioscopic este de - 0,2C pn la - 0,4C. Densitatea salivei este de 1002- 1012 g/cm3. Densitatea variaz n limite largi, n funcie de debitul salivar. Vscozitatea salivei mixte este 1,08-1,32 uniti, i este dat de gncoproteinele hidratate.
2.1.7. Compoziia chimic a salivei Saliva conine 99,5% ap i 0,5% reziduu uscat format din substane anorganice 0,2% i substane organice 0,3%. 2.1.7.1. Substanele anorganice din saliva mixt bucal (tabelul 1). Tabelul 1. Concentraia principalelor substanelor anorganice n saliva mixt, comparativ cu plasma sanguin.
SALIV (mEq/1)PLASM (mEq/1)Na+8,7-32,9< + + 142K 12-16> 5 Ca+ 2,5-5,55Mg+ 0,1-13cr8,4-17,7< 103P04~7-212,4-4,4HCOj-10-6027
+
35
LAS MA
0 4,0
1,0 2,0
3,0
ml / min Fig. 4. Fluxul i componenii salivari (dup BRAY, 1989, modificat) Natriul, clorul sunt n concentraie mai mic n saliv dect n plasm. Clorul asigur activarea amilazei salivare. Potasiul este mai mare dect n plasma, bicarbonatul crete n saliv n urma stimulrii, ajungnd la 60 mEq/1 i particila la sistemul tampon salivar HCO3/H2CO3. (Fig.4) Calciul are concentraie asemntoare cu cea din plasm. n saliv calciu se gsete sub form de sruri anorganice i sub form de compui organici, fixat de macromolecule. Saliva poate fi considerat o soluie saturat de fosfat de calciu ce mpiedic disoluia calciului din smal. Cu toat aceast saturare, saliva nu permite precipitarea fosfailor respectivi datorit asocierilor cu proteinele salivare acide. Saturarea salivei cu fosfat de calciu asociat cu un pH alcalin salivar determin precipitarea srurilor, formnd sialolii (calculi salivari) sau, la nivelul dinilor produce tartru dentar. Scderea local a pH-ului, sub 5,2, favorizeaz disoluia cristalelor de hidroxiapatit i apariia cariilor. pH-ul critic este pH-ul la care au loc micri ale calciului i fosfatului n smalul dentar. Fluorul (0,01-0,05 ppm) se gsete n concentraie aproximativ cu cea din plasm, depinde de aportul alimentar de fluor, are rol n formarea Fiuorapatitei care asigur rezistena smalului. Tiocianatul se afl n concentraie mai mare dect n plasm i are un rol antibacterian, inhibnd dezvoltarea bacteriilor Gram pozitive i Gram negative, inhib creterea ciupercilor, virusurilor, micoplasmelor. n saliv se secret i iod cu rol mai puin cunoscut. Ocazional, se pot gsi n saliv sruri de plumb, de mercur. 2.1.7.2. Substanele organice se mpart n dou categorii: substane azotate (proteice i neproteice) i substane neazotate (tabelul 2).
35
Tabelul 2.
2.1.7.2.1. Substanele azotate proteice cuprind: proteine serice, proteine de origine glandular i hormoni salivari. Proteinele serice cuprind trei mari categorii: imunoglobuline, lactoferin, factori ai coagulrii. Imunoglobulinele IgG, IgM apar n saliv printr-un proces de trecere din snge in saliv. IgA secretor este sintetizat n glandele salivare. IgA secretorie se combin cu bacteriile din mediul bucal mpiedecnd aderena lor la mucoas, formeaz un strat protector antibacterian la suprafaa mucoasei bucale. Imunoglobulinele asigur aprarea antibacterian prin: aglutinarea bacteriilor, neutralizarea enzimelor, toxine bacteriene i virale, previn caria i parodontopatia. Lactoferina fixeaz fierul inhibnd multiplicarea bacteriilor ferodepente. Factori ai coagulrii sunt: factorii VII,VIII,IX plasmatici, factorul 3 plachetar. Proteinele de origine glandular pot fi mprite la rndul lor n mai multe categorii i anume: enzime, mucine, substane de grup sanguin, factor de adredare al bacteriilor, proteine bogate n prolin. Enzimele ca de exemplu: -amilaz salivar hidrolizeaz amidonul fiert sau copt pn la maltoz trecnd prin stadii intermediare de dextrine; -lipaza salivar este important la sugar datorit descompunerii lipidelor din lapte -lizozimul distruge mucopolizaharidele din peretele bacteriilor; -kalicrein- enzim proteolitic, acioneaz asupra kininogenului, formnd kinine (de exemplu bradikinin cu rol vasodilatator, crete permeabilitatea vascular, activeaz fagocitoza). Mucinele sunt glicoproteine ce conin o component proteic (apomucina) component glucidic (manoza, galactoza), au ca proprieti vscozitate, elasticitate i aderen crescut. Au rol de aprare, lubrefiere, de formare a bolului alimentar, asigur masticaia, deglutiia i vorbirea, particip la sistemele tampon salivare. Substanele de grup sanguin. In saliv pot fi secretate antigene H, A, B de grup sanguin la subiecii secretori. Factorul de agregare al bacteriilor are aciune agregant pentru microorganismele din placa dentar, agregare ce necesit prezena calciului i un Ph cuprins de la 5 pn la 7,5. Proteinele bogate n prolin pot fi cu caracter acid, cu caracter bazic i glicolizate. Dintre proteinele bogate n prolin amintim: Proteina A,C care au rol n fixarea calciului salivar. n medie, 5-52% din calciul total salivar este fixat pe aceste proteine, ceea ce determin: meninerea constant a calciului salivar, ntrzie micarea ionilor ntre suprafaa smalului i lichidele din jurul dinilor. De asemenea, din grupul Proteinelor B, staterina este un polipeptid cu 43 aminoacizi i GM-5380. Staterina inhib precipitarea spontan
2 a calciului i creterea cristalelor de fosfat de calciu (prin fixarea la suprafaa fosfatului de calciu), menine saliva n condiii de suprasaturare necesare stabilizrii i recal-cifierii smalului la suprafa, asigur integritatea smalului n timpul atacului acid inhib formarea calculilor n duetele salivare, inhib formarea osului. In categoria hormonilor salivari enumerm: hormonii steroidici, parotina, factorul de cretere al nervilor, factorul de cretere epidermal, factorii de cretere insulin-like. Corticosteroizii, se gsesc liberi n saliv, n concentraii mai mici dect n snge. Estrogenii sunt metabolizai n glandele salivare, estrona este transformat n estradiol sub aciunea unei enzime oxidoreductoare. Hormonii sexuali masculini sunt transformai, n glandele salivare, n metabolii mai activi fiziologici. Parotina este un polipeptid ce scade calciul seric, fixndu-1 n oase i structurile dentare, stimuleaz calcifierea, vascularizaia, sistemul reticulo-endotelial, crete anabolismul proteic. Factorul de cretere al nervilor (NGF) este un polipeptid ce activeaz creterea i dezvoltarea esutului nervos n perioada embrio-fetal i accelereaz erupia dinilor. Factorul de cretere epidermal (EGF) este un polipeptid cu greutate molecular de 6 kdaltoni, stimuleaz erupia dentar, asigur calcifierea precoce a incisivilor, asigur keratinizarea (creterea n grosime a epidermei), diferenierea precoce a odontosau ameloblastelor, faciliteaz vindecarea mucoasei bucale dup leziuni, ulceraii. Factorii de cretere insulin-like (IGFs) au efect mitogen pentru fibroblati (care au rol reparator), activeaz proliferarea lor. 2.1.7.2.2. Substane organice azotate neproteice sunt: uree, acid uric, creatinin, amoniac, substane ce provin din catabolismul proteic. Ureea sanguin trece prin difuzie simpl n saliv. Concentraia ureei din saliv variaz n funcie de fluxul salivar i de concentraia sanguin. Ureea salivar, n prezena ureazei bacteriene se transform n amoniac. Acidul uric se gsete n saliv n concentraie aproape constant. Amoniacul salivar se formeaz prin dezaminarea aminoacizilor sub aciunea bacteriilor. Creatinin din saliv depinde de nivelul sanguin. 2.1.7.2.3. Substane organice neazotate sunt: glucidele i lipidele. Concentraia glucidelor n saliv depinde de concentraia sanguin.
3 La
diabetici crete cantitatea de glucoza n saliv i lichidul crevicular, care va fi metabolizat de bacillus acidophilus ceea ce duce la acidifierea local a pH-ului, aceasta favoriznd apariia cariilor i parodontopatiei. Lactatul salivar crete de circa 10 ori dup ingerarea hranei, ce conine zaharuri. Concentraia lipidelor n saliv este mic, gsindu-se colesterol i acizi grai nesaturati. Importanti sunt produsii secretati rezultati din catabolismul acidului arahidonic (prostaglandine, leucotriene cu rol n procesul inflamator local, n mecanismele de aprare ale cavitii bucale i n procesele de absorbie de la nivelul tractului gastrointestinal).
2.1.8. Rolurile salivei Saliva, pe lng rolul digestiv pe care l are, este un element esenial meninerea troficitii normale a esuturilor din cavitatea bucal i prin aceasta, n meninerea structurii i stabilitii dinilor n alveola. Rolurile salivei sunt multiple: digestiv, protectiv, excretor, endocrin, n stazia hidro-electrolitic, n termoreglare, n vorbire. 2.1.8.1. Rolul digestiv al salivei cuprinde mai multe etape: a. preparativ: - de pregtire a alimentelor prin masticare, insalivare i lubrefiere (cu participarea mucinelor), formarea bolului alimentar cu ajutorul mucinelor ce ncorporeaz alimentele ntr-o pelicul, - pasajul bolului alimentar n timpul bucal al deglutiiei. Prin mucinele coninute, saliva lubrefiaz bolul alimentar i mucoasa, favorizeaza masticaia i deglutiia. B. hidrolitic: - n care amilaz salivar (ptialina) hidrolizeaz amidonul u copt pn la stadiul de maltoz, trecnd prin stadii intermediare de e (amilo-, eritro-, acrodextrin). Amilaz hidrolizeaz i alimentele de ^lucidic ce rmn depuse pe dini sau n spaiile interdentare (resturi de , jeleuri, caramele, etc). c. gastronomic: - de solubilizare a constituenilor alimentari i astfel de stimulare a receptorilor gustativi inducnd senzaia de gust i reflexele secretorii gastrice, pancreatice. 2.1.8.2. Funcia proiectiv a salivei - de meninere a troficitii normale a tesuturilor bucale, se realizeaz prin: 2.1.8.2.1. Lubrefierea structurilor moi i dure din cavitatea bucal, asigurata de glicoproteinele bogate n prolin, ce formeaz un film de mucin la suprafata alimentelor i structurilor buco-dentare. Acest film lubrefiant de mucin faciliteaz: masticaia, pasajul alimentelor pentru deglutiie, vorbirea, protejeaz mucoasa de evaporarea apei i uscare, de frecrile cu alimentele dure i corpii
4 straini; 2.1.8.2.2. Meninerea integritii mucoaselor din cavitatea bucal prin mucine si fosfoproteine pe baz de cistein (cistatine). Mucinele au solubilitate sczut, au vscozitate, elasticitate i adezivitate crescut, formeaz un film impermeabil aderent, care controleaz permeabilitatea suprafeelor mucoase, limiteaz ptrunderea agenilor iritani, toxici. Cistatinele se leag de hidroxiapatita, inhiba creterea cristalelor, asigur aprarea mucoasei bucale, sunt inhibate de cisteinproteinaze i catepsin. Reparaia esuturilor moi realizat de: factorii de cretere din saliva: NGF, EGF, IGF care activeaz fibroblatii; factorii procoagulani (VII, VIII, IX, Fp3) i anticoagulani din saliv. Plasminogenul salivar are rol n hemostaz dar i n procesele reparatorii din cavitatea bucal. Meninerea echilibrului ecologic microbian: prezena bacteriilor saprofite mpiedic multiplicarea bacteriilor patogene i aderena lor pe dini; Lavajul cavitii bucale: - asigur ndeprtarea mecanic a resturilor alimentare, bacteriilor de pe suprafaa dinilor, mucoaselor. Acest lavaj poate fi considerat echivalent cu alte reflexe de aprare, de tip: tuse, strnut, clipit. Agregarea bacterian este asigurat de sistemele directe agregante reprezentate de IgA salivar, mucine, liani, lizozim, amilaz, factori de agregare bacterian, staterine. Imunoglobulinele: - neutralizeaz enzimele, toxinele bacteriene, virale; aglutineaz bacteriile; opsonizeaz bacteriile, pregtindu-le pentru fagocitoz, n leucocite. Mucinele i lianii formai prin polimerizarea glucozei salivare (dextrani) sau a fructozei (levani), sub aciunea enzimelor microbiene, particip la formarea plcii dentare prin unirea resturilor alimentare celulare i prin facilitarea ulterior a invaziei bacteriene. Staterinele stimuleaz ataarea microbilor de hidroxiapatita i formarea plcii dentare. 2.1.8.2.7. Funcia antibacterian, antifungic i antiviral direct Funcia antibacterian direct se realizeaz printr-un mecanism complex la care particip: a) proteine salivare: - lizozim cu efect bacteriolitic; lactoferina cu rol de fixare a fierului, inhib multiplicarea bacteriilor ferodependente;
5 sistemul peroxidazic determin oxidarea tiocianatului salivar i formarea hipotiocianatului, agent puternic oxidant (rol antimicrobian). Sistemul peroxidazic cuprinde: lactoperioxidaza secretat de glandele salivare i mieloperoxidaza eliberat de polimorfonuclearele neutrofile. Ig secretate de glandele salivare (IgA secretor, IgM) i provenite din chidul gingival (IgM, IgG). leucocitele neutrofile care trec prin diapedez din vasele gingivale n interstiiu i cavitatea bucal, au capacitate fagocitar i secret substane antibacteriene. fluorul salivar: - inhib enolaza, - mpiedic dezvoltarea bacteriilor. Funcia antifungic este realizat de histatine, proteine cationice bogate n istidin. La aprarea antiviral intervin:lactoferina, lizozim, sistemul peroxidazic, gA, mucina, ct i secreia de virusuri. 2.1.8.2.8. Meninerea echilibrului acido-bazic local i neutralizarea substanelor chimice ptrunse accidental n cavitatea bucal (acizi, alcali), este asigurat de sistemele tampon salivare ale bicarbonailor (care neutralizeaz acizii), ale fosfailor, (care neutralizeaz bazele), ale mucinei. Ureea, n prezena reazei salivare de origine bacterian, se transform n ion de amoniu cu rol antiacid. 2.1.8.2.9. Meninerea integritii dinilor. n perioada posteruptiv, de maturaie, a dinilor saliva devine o soluie saturat de sruri de calciu, fosfor, magneziu, care mpiedic solubilizarea hidroxiapatitei din email. Staterinele (proteine salivare bogate n prolin) leag calciu, asigurnd saturaia salivei cu sruri de fosfat de calciu. fluorul prezent n saliva particip la formarea fluorapatitei, ceea ce crete rezistena emailului, fluxul salivar: - spal reziduurile alimentare mpiedic stagnarea, aderena i descompunerea alimentelor. 2.1.8.3. Funcia excretorie a salivei const din eliminarea pe aceast cale a:
6 metaboliilor din catabolismul proteic (uree, acid uric, amoniac) i substanelor cu molecula mic (nitrii, nitrai, tiocianat); substanelor toxice ptrunse accidental n organism - sruri ale metalelor grele Pb, Hg, Bi (formeaz lizereu gingival -semn al intoxicaiilor profesionale); medicamentelor anticonvulsivante, antidepresive, citostatice, cofeina, tetraciclina; substanelor toxice ca alcoolul, cocaina, nicotina; hormonilor circulani cum ar fi cortizol, aldosterori, estradiol, progesteron; la subiecii secretori; substanelor de grup sanguin de exemplu aglutinogenele H, A, B -
virusurilor: - poliomielitei, parotiditei epidemice, (este o cale de rspndire a infeciei n colectivitile de copii), rabiei. 2.1.8.4. Funcia endocrin a glandelor salivare - se realizeaz prin secreia de hormoni proprii: parotin, NGF, EOF, IGFs (insulin-like growth factors), glucagon-like growth factors, care au fost descrii la capitolul 2.7.2. 2.1.8.5. Rolul salivei n homeostazia hidro-electrolitic n deshidratri se reduce lichidul extracelular, scade secreia salivar, mucoasa bucal se usuc, ceea ce declanseaz, prin intermediul hipotalamusului, fie senzaie de sete cu creterea aportului hidric, fie stimularea secreiei de ADH (reine apa n organism, mpiedic pierderea de ap). ALDOSTERONUL - stimuleaz reabsorbia Na+ i secundar a Cldin saliva ductal i eliminarea de potasiu. 2.1.8.6. Rolul salivei in termoreglare La om acest rol este mai puin important, ns la cine este primordial. Cinele i realizeaz termoliza prin hiperventilaie cu gura deschis i limba scoas, favorizndu-se evaporarea. 2.1.8.7. Rolul salivei n vorbire - se realizeaz prin umectarea i lubrefierea mucoasei bucale, ceea ce faciliteaz micrile limbii, fonaia i vorbirea. Atmosfera cald i uscat ngreuneaz vorbirea prelungit. Emoiile scad secreia salivar, ngreunnd vorbirea. 2.1.9. Reglarea secreiei salivare Secreia salivar este declanat exclusiv pe cale reflex,
7 influenele umorale au doar rol corector. Reglarea secreiei prin mecanisme reflexe necondiionate i condiionate au fost precizate n urma cercetrii colii pavloviste. 2.1.9.1. Reflexul salivar necondiionat este cel mai important reflex de reglare a secreiei salivare. Arcul reflex cuprinde: zone reflexogene, ci aferente, centrii nervoi i ci eferente simpatice i parasimpatice. 2.1.9.1.1. Zone reflexogene i ci aferente - excitaiile receptorilor celor trei zone reflexogene sunt transmise prin urmtoarele ci aferente: a) Excitaiile specifice ale mugurilor gustativi din cele 2/3 anterioare ale limbii, de ctre alimente, sunt transmise prin nervul timpanico-lingual la ganglionul geniculat i de aici la central salivator superior (din punte). Pentru treimea posterioar a limbii, excitaiile ajung prin nervul glosofaringian la ganglionul pietros i de aici la central salivator inferior (din bulb). Saliva secretat n urma excitaiilor mugurilor gustativi provine cu predominant din glandele submandibulare i sublingual ("saliva de gustaie"). De la nivelul mucoasei bucale i a dinilor, sensibilitatea nespecific prin excitaii determinate de: consistena alimentelor n timpul masticaiei, durere i vorbire sunt transmise prin ramuri ale trigemenului la ganglionul Gasser i de aici la ambii centrii salivari ce vor secreta "saliva de masticaie". De la nivelul mucoaselor laringian, faringian, esofagian, gastric excitaiile determinate de iritaie, distensie (bol voluminos) sunt transmise prin fibrele nervului vag la centrul salivar inferior. Centrii nervoi sunt localizai n substana reticulat bulboprotuberanial. Funcional, exist un centru salivar superior n punte (regleaz secreia glandelor submandibular, sublingual). i un centru salivar inferior n bulb (regleaz secreia glandei parotide). Cile eferente, centrifuge sunt reprezentate de fibre ale sistemului vegetativ ce pleac de la centrii la glandele salivare. Pentru glanda parotid - fibrele parasimpatice cu origine n centrul salivar din bulb se ataeaz nervului glosofaringian, apoi la nivelul gurii jugulare ptrund n nervul lacobson (cu origine n ganglionul Andersch) apoi n nervul mic pietros superficial i ajung n ganglionul optic unde fac sinapsa. Fibrele postganglionare urmeaz traiectul nervului auriculo-temporalului i ajung la glanda parotid. (Fig. 5.).
Pentru glanda submandibular, sublingual - fibrele parasimpatice au origine n centrul salivar superior din punte, iau calea nervului intermediar al lui Wriesberg, trec n ganglionul geniculat al facialului, coboar prin nervul coarda timpanului, care n apropierea cavitii bucale se unete cu nervul lingual, se desprind din aceasta i fac sinapsa n mai muli ganglioni mici din glanda submandibular sau n ganglionul sublingual de unde fibrele postganglionare se termin n glandele respective. Fibrele simpatice, au origine comun pentru toate glandele salivare i sunt asigurate prin fibre preganglionare din mduva toracal segmentele T1-T2 ce provin din coarnele antero-laterale. Excitaiile prsesc mduva prin rdcinile anterioare, apoi prin ramurile comunicante albe ajung n lanul ganglionilor simpatici paravertebrali i fac sinapsa n ganglionul cervical superior; de aici fibrele postganglionare ajung pe calea plexurilor perivasculare la glandele salivare. Glandele mici primesc inervaia efectoare de la nervul glosofaringian. Reflexele salivare necondiionate cu punct de plecare bucal mai pot fi determinate i de contactul chemoreceptorilor cu substane insipide, acide, amare, de stimulare a proprioceptorilor si mecanoreceptorilor n cursul tratamentelor stomatologice, de contactul cu aparate dentare. Aceast variat stimulare este culeas de fibrele nervului facial, trigemen si glosofaringian. Excitaiile nociceptive de la orice nivel al organismului determina hipersalivaie. 2.1.9.2. Reflexul salivar condiional Mecanismul reflex condiionat dovedete influena cortical asupra centrilor bulbo-protuberaniali. Prezint importan, deoarece, cnd alimentele ajung n cavitatea bucal, gsesc deja o cantitate de saliv. Excitani nespecifici, vederea, mirosul, zgomotul, lumina, cuvntul (evocarea alimentelor) prin asocierea cu excitantul
9 necondiionat, devin excitani adecvai ai secreiei salivare.. Reflexul salivar condiionat are aceei cale eferent cu cel necondiionat, cea aferent este corespunztoare regiunii respective, miros, vz, auz. Se elaboreaz pe baza legturii temporare ntre zonele senzoriale ale scoarei (olfactiv, vizual, auditiv) cu reprezentarea centrului secreiei salivare. Fazele salivaiei sunt: faza cefalic (gndul, mirosul, vederea alimentelor); faza bucal (contactul alimentelor cu receptorii); faza gastric (cnd alimentele au ajuns n stomac). n toate cele trei faze se declaneaz stimularea secreiei salivare. Centrii superiori hipotalamici i corticali exercit influene asupra reflexului salivar. Impulsurile salivare hipotalamice pot apare n cadrul reaciilor: alimentare prin stimularea centrului foamei, apetitului; de agresivitate, emoii, prin stimularea hipotalamusului posterior; de termoreglare, prin stimularea hipotalamusului anterior. Influenele corticale asupra fluxului salivar se constat n: condiionarea secreiei salivare, hiposalivaii, n emoii, excitarea unor zone corticale n vecintatea gustului, mirosului. Interrelaiile funcionale, ntre respiraie, vom, salivaie se explic prin situaia anatomic, de vecintate a centrilor respectivi. 2.1.9.3. Efectele inervaiei parasimpatice i simpatice Stimularea parasimpaticului prin eliberarea mediatorului chimic acetil-colina la nivelul sinapselor neuro-glandulare are ca efect formarea unei salive bogate n volum, NaCl, amilaz, srac n K+, CO3H-, mucin, cu activitate bactericid redus. Acetilcolina acioneaz asupra receptorilor colinergici, ce pot fi blocai de atropina, substana ce mpiedic aciunea stimulatoare a parasimpaticului. Acest fapt se observ i n terapeutic, dup administrarea medicamentelor ce conin atropina, apare uscciunea mucoasei bucale. Stimularea simpaticului prin eliberarea catecolaminelor: adrenalina si noradrenalina, ce acioneaz pe receptorii alfa (secreie de K+ i ap) i beta (secreie de amilaz) are ca efect formarea unui volum redus de saliv, srac n NaCl, vscoas, bogat n mucin, n substane organice i K+, HCO-3, lizozim. Cele dou ci eferente, asupra secreiei salivare, au efect stimulator, sinergic cu anumite diferene, existnd deci o completare funcional. Efectele denervrii. Secionarea nervilor simpatici i parasimpatici este urmat de hipersecreie salivar numit paralitic (ClaudeBernard) i se datorete sensibilizrii glandei denervate la aciunea adrenalinei, noradrenalinei n cantiti crescute. Astfel stimularea continu a glandei atrofiate la nceput restabilete greutatea, iar glanda salivar simetrica celei denervate se hipertrofiaz prin mecanism compensator. 2.1.9.4. Adaptarea secreiei salivare
10 Factorii care influeneaz compoziia i volumul salivar sunt numeroi incluznd: natura stimulului: introducerea n cavitatea bucal de pulberi determin secreia unei salive apoase, ptrunderea accidental a unor acizi puternici n cavitatea bucal determin o saliv alcalin, alimentele uscate induc secreia unei salive bogate n mucin; creterea debitului salivar determin creterea salivar a Na+, HCO-3, amilazei i scderea fosfailor; regimul alimentar bogat n hidrocarbonate determin creterea amilazei salivare, vegetalele (spanac) cresc puterea tampon a salivei, regimul bogat n proteine determin creterea ureei n saliv, a puterii tampon i scderea amilazei; ritmul circadian: Na+, Cl- sunt crescui n saliva de diminea, Ca++, fosfatul cresc n secreia nocturn, K+ este mai crescut dup amiaz, enzimele salivare sunt crescute n perioadele alimentare; constelaia hormonal: testosteronul, tiroxina, graviditatea cresc secreia salivar, menopauza determin scderea secreiei salivare, aldosteronul controleaza eliminrile de Na+ si K+ n saliv, parathormonul crete semnificativ concentraia proteinelor, calciului, fosfatului n saliv, bradikinina are efect sialogog; medicamente: secreia salivar este stimulat de parasimpaticomimetice (policarpina), anticolinesterazice (prostigmina, ezerina) i inhibat de parasim- paticolitice (atropin, scopolamin) i simpaticolitice (ergotamin). Anestezicele: eter, cloroform, ciclopropan induc hipersalivaia reflex. 2.1.9.5. Perturbrile secreiei salivare Secreia salivar poate fi suprimat temporar n stri emoionale nsoite de anxietate, stri febrile i deshidratare. Suprimarea permanent a secreiei salivare se numete xerostomie sau aptialism. Hipersalivaia (sialoree) se ntalnete graviditate, carii dentare, iritaii bucale, ale limbii, esofagiene, ulcer gastro-duo-denal, pancreatita, tulburri neuro-psihice (Parkinson, schizofrenie).
2.2. LICHIDUL CREVICULAR (SULCULAR)
Lichidul crevicular (al anului gingival) este descris n literatur din 1817. Este un lichid care se observ la coletul dintelui, n anul
11 gingival, dup uscarea i izolarea fluxului salivar. Este un element al mediului bucal. Conine factori de origine seric; este un element provizoriu fiind nghitit cu saliva; este n cantititate inconstant depinznd de starea inflamatorie la locul de producere. Locul de formare al lichidului anului gingival este delimitat de: epiteliul gingival necheratinizat, rsfrnt n jos, i smal, cu deschidere printr-un spaiu virtual spre cavitatea bucal. La nivelul epiteliului gingival exist o vascularizaie foarte bogat i mare permeabilitate. Aceast permeabilitate mare vascular i epitelial condiioneaz apariia lichidului crevicular. Prin spaiile intercelulare are loc ieirea pasiv a componentelor lichidului. 2.2.1. Mecanismul formrii lichidului cervicular PASHLEY a propus o modalitate ce explic formarea lichidului crevicular Formarea depinde de diferena de presiune hidrostatic i coloidosmotic ntre capilarele sanguine, vasele limfatice, lichidul interstiial i lichidul crevicular: Debit de fluid (ml/min) = C - (L+G) n care: C = coeficientul de filtrare capilar; L = coeficientul de filtrare limfatic; G = coeficientul de filtrare lichidului crevicular. Vascularizaia bogat permite filtrarea unui lichid interstiial, care este apoi preluat de limfatice. Dac, filtrarea vascular depaeste resorbia limfatic i venoas, lichidul se acumuleaz rezultnd edemul. Acest lichid, n exces, este n funcie de: coeficientul de filtrare epitelial, diferena de presiune oncotic (coloidosmotic) a lichidului interstiial, diferena de presiune oncotic din an, care depinde de acumularea de diferite substane i metabolii, produi n principal de placa bacterian dentar, substane ce pot fi eliminate prin exocitoz. Acumularea de substane n anul gingival duce la creterea local a presiunii coloidosmotice, iar lichidul interstiial va migra spre anul gingival, antrennd spre cavitatea bucal masa de substane acumulate. Dac exist o agresiune toxic se dezvolt o reacie inflamatorie cu formarea exudatului inflamator mai bogat n proteine. Aplicarea local de histamin crete debitul lichidului crevicular prin creterea filtrrii capilare. Punerea n evidena a lichidului crevicular se poate face: benzi de hrtie de filtru aplicate n anul gingival; cu micropipete; cu prin splarea anului gingival.
12 2.2.2. Compoziia lichidului cervicular 2.2.2.1. Componenii organici a. Proteine: 70g/l (61-92g/l) sunt de origine plasmatic: albumine, alfa1-globuline: oromucoid, alfa1-antitripsin, alfa2-globuline: ceruloplasmin, alfa2 macroglobulin, alfa2 glicoproteine, haptoglobin, beta-globuline: beta-lipoproteine, transferin, gama-globuline: IgG, IgA, IgM care provin din plasm, fiind sintetizate de plasmocite, au rol n aprarea gingival (IgA, IgG), fibrinogen, bradikinin, cu rol vasodilatator, factori ai sistemului complement din plasm i sintetizai local de gingia inflamat. b. Lipide: - fosfolipidele plasmatice i bacteriene, - PGE2 in concentraie mai mare dect n snge fiind sintetizat local; este un indice al inflamaiei gingivale. c. Glucide: - glucoza poate crete la diabetici n lichidul crevicular. d. Alte substane: hidroxiprolina, rezultat din degradarea colagenului, H2S din bacterii (indic gradul inflamaiei), acid lactic, endotoxine (lipopolizaharide, LPS), uree, n concentraie mai mare dect n plasm i saliv datorit unui proces de concentrare local. Este transformat n amoniac de ctre ureazele bacteriene, determinnd alcalinizarea anului i plcii dentare. e. Enzime: de origine tisular sau bacterian. De exemplu: hialuronidaze din leucocite, streptococ, pneumococ; beta-glicuronidaze, beta-galactozidaze, din lizozomii macrofagelor, fibroblasti, celule epiteliale gingivale, bacterii; colagenaz tisular i bacterian; elastaza ce degradeaz fibrinogenul, colagenul, hemoglobina, imunoglobulinele, factorii complementului; catepsina - enzim proteolitic; fosfataz acid, corelat cu inflamaia; este de origine leucocitar, celule epiteliale descuamate, macrofage, bacterii (50%);
13 fosfataz alcalin leucocitar; lacticodehidrogenaz; sistemul plasminogen-plasmin, cu activitate fibrinolitic asupra cheagului sanguin de fibrin, determinnd liza sa. Liza precoce ntrzie cicatrizarea, liza tardiv determin formarea de esut de granulaie abundent cu cicatrice hipertrofic. f. Enzime antibacteriene: lizozim; cu rol antibacterian, antimucolitic, accelereaz eliberarea local a enzimelor din bacterii (colagenaz, hialuronidaz); peroxidaza: antibacterian, antifungic, antiviral, antimico-plasm. 2.2.2.2. Componeni anorganici: variaz n limite foarte largi Ca++ : 2,70-11.9 mmol/1 Fosfat-: 0,42 - 2,30 mmol/1 Mg++ : 0,20 - 0,60 mmol/1 Na+ : 105-222 mmol/1 K+ : 9,5 - 69 mmol/1, din liza celular Fluor ca n plasm. 2.2.2.3. Elemente celulare: a. celule epiteliate descuamate, b. leucocite: 95-97% PMN, 1-2% limfocite, 2-3% monocite, (sunt crescute n gingivite cronice). Capacitatea fagocitar este mai mic dect n snge. c. bacterii: din placa dentar, pot elibera endotoxine, contribuind la iniierea i ntreinerea inflamaiei gingivale. Cantitatea de lichid variaz cu ciclul menstrual, fiind maxim la ovulaie. Tetraciclina trece n lichid, atingnd valori maxime dup 6 ore de la administrare. 2.2.3. Rolul lichidului cervicular mecanic, de diluie i splare a substanelor din anul gingival, de aprare: realizat prin lizozim, imunoglobuline, peroxidaz, leucocite. Acest rol crete n cazul gingivitelor, lichidul devenind un veritabil exudat inflamator.
2.3. Halena
Halena reprezint mirosul cavitaii bucale i este dat de: stagnarea resturilor alimentare sau a celulelor epiteliale, ca urmare a reducerii fluxului salivar sau micarilor masticatorii. Materialul acumulat este distrus de bacteriile orale ce metabolizeaz
14 resturile proteice; distrucii tisulare n bolile parodontale sau carii, asociate cu creterea activitii microbiene; eliminarea de substane odorifere ce contin mercaptan, hidrogen sulfurat; vorbirea prelungit, foamea, situaii n care saliva nsi d mirosul prin acumularea constituenilor salivari n gur i degradarea lor de ctre bacterii. Creterea vscozitii salivare este un factor favorizant al acumulrii de constitueni salivari n cavitatea bucal. Aportul alimentar reduce parial halena prin creterea fluxului salivar, prin micrile bucale, iar prin aportul de carbohidrate se asigur substratul pentru bacteriile productoare de acizi, bacterii care vor prolifera. Dezvoltarea acestora vor suprima bacteriile ce metabolizeaz proteinele i derivaii proteici. Posibiliti de diminuare a halenei: splatul periodic al dinilor, ndeprtarea resturilor alimentare, cltitul gurii cu soluii antiseptice, consum frecvent de lichide, stimularea secreiei salivare, folosirea de ageni oxidani care s previn transferul de hidrogen.
2.4. FIZIOLOGIA PLCII DENTARE
Smalul mpreun cu mediul specific n care se afl (saliva mixt, hran i bacterii) permite formarea unei structuri distincte numite placa dentar. n compoziia acesteia gsim bacterii specifice, matrice, fluid. Se realizeaz astfel un ecosistem specific, independent de restul cavitii bucale. Definiia plcii dentare: este depozitul care se formeaz pe dinii din mediul lor natural, depozit ce nu poate fi ndeprtat prin simpla splare ci numai prin periajul energic al dinilor. Placa dentar conine: bacterii specifice, celule degradate, matrice organice glucido-proteic, mediu lichidian, derivat din saliv. Localizarea plcii dentare: pe suprafeele neocluzive ale dinilor, la punctele de contact interdentar, la marginile gingivale, pe feele alterale, fisurate ale dinilor, zone n care se pot aduna resturi alimentare. 2.4.1. Etapele formrii plcii dentare 2.4.1.1. Formarea filmului acelular (pelicula dentar) Prin splarea dinilor cu un material abraziv se ndeprteaz materialul organic de pe suprafaa dinilor. Dar, n scurt timp de la contactul suprafeei dentare cu saliv se formeaz un film acelular fin, numit pelicula catigat (90 minute). Aceasta are o grosime de 10 micrometri, dou straturi (stratul de suprafa este gelatinos) i
15 conine: proteine provenite din saliv, carbohidrai i glicoproteine cu hexoze, hexozamina, fucoz. Proteinele precipitate n prezena ionilor de calciu i fosfor au proprieti adezive, se leag att de cristalele de hidroxiapatit ct i de cele de fosfat de calciu de pe suprafaa smalului. Prezena complexelor proteine - fosfat de calciu determin creterea rezistenei smalului la disoluie. Rolurile peliculei dentare: protector: prin complexele proteine - fosfat de calciu se rol realizeaz o barier care mpiedic difuzia ionilor spre mediul bucal. De asemenea, constitute o barier n calea acizilor care au tendina s difuzeze spre suprafaa smalului. distructiv: prin faptul c fixeaz bacteriile orale ce vor forma rol placa dentar. 2.4.1.2. Formarea matricei plcii Matricea extracelular a plcii, format n absena hranei, este subire, relativ poroas i conine complexe insolubile de proteine fosfat de calciu mpreun cu glicoproteine salivare modificate. Dezvoltarea matricei dureaz circa 24 ore. Structura poroas permite ptrunderea oxigenului, salivei, lichidului gingival, produilor bacterieni. Matricea format n prezena hranei (carbohidrai) este gelatinoas, poate fi comparat cu matricea extracelular a esutului conjunctiv. Carbohidraii extracelulari sunt polimeri de glucoz (dextrani, glucani, levani, etc.) ce formeaz o structur gel-like. Aceast plas gel-like favorizeaz acumularea de material insolubil, limiteaz micarea liber a moleculelor ntre fluidul plcii i saliva, creeaz condiii anaerobe, crete producerea de acid de ctre bacterii i acumularea local a acizilor. Polizaharidele extracelulare reprezint rezerva de carbohidrai pentru bacterii. 2.4.1.3. Colonizarea bacterian Pelicula format este rapid invadat de bacterii originare din: saliv, esuturi moi adiacente, suprafaa lezat a smalului. Colonizarea plcii se datorete proprietilor adezive ale microorganismelor, adic capacitatea celulelor bacteriene de a se fixa pe suprafee solide i de a adera una de alta. Se realizeaz astfel interaciuni dintre suprafeele solide si bacteriile din lichidul supernatant. Aceste interaciuni se realizeaz n urmatoarele etape:
16 depunerea iniial reversibil, a unei singure celule la suprafaa solid, proces dependent de forje fizico-chimice noncovalente, n particular interaciuni ionice i fore van der Waals. Cristalele de hidroxiapatit la pH 6,5 au ncrctura net pozitiv, care atrage uor bacteriile. adeziunea ireversibil a celulei fixate se realizeaz prin interaciuni hidrofobice, legturi covalente (la distane sub 0,4 nm), puni de polimeri (la distane peste 10 nm). Colonizarea suprafeei implic creterea i multiplicarea bacteriilor care produc iniial colonii discrete ce fuzioneaz apoi formnd o mas bacterian. Multiplicarea bacterian este favorizat de factorii de cretere produi de nsi bacteriile colonizatoare care elibereaz, de asemenea, extracelular proteine i polizaharide.
2.4.2. Componentele principale ale plcii dentare mature Placa dentar este format din: ap 80%, din care intracelular 50% si extracelular 30%; componente organice (18-20%): 1. de origine bacterian (10%) 2. proteine extracelulare (6%) 3. polizaharide extracelulare (2%); componente anorganice, n proporii variabile. Proteinele matricei acioneaz ca polielectrolii, ionizarea lor depinde de pH-ul local. Matricea funcioneaz ca un sistem de gel filtrant, ca un schimbtor ionic (micarea ionilor depinde de gradientul de densitate local). Glucidele i ureea difuzeaz uor n matrice. Acizii organici i ionii de amoniu produi de metabolismul bacterian pot fi reinui n matricea plcii. Matricea plcii mature este o barier eficace pentru migrarea apei, ceea ce face ca apa s fie parial imobilizat n plac, iar bacteriile s fie parial izolate de restul cavitii bucale. Calciul, fosfatul, potasiul sunt n concentraie mai mare n plac dect n saliv, probabil printr-un mecanism local de concentrare. Fluorul n plac variaz n limite largi (6-180 ppm). Fluorul din plac poate proveni din trei surse: suprafaa smalului, lichidele sau alimentele ingerate, saliva i lichidul crevicular (reprezint sursa major de fluor). Rolurile fluorului n placa dentar sunt:
17 reduce solubilitatea smalului la acizi, inhib enzima glicolitic enolaza, reduce producerea de acizi de ctre bacterii (pe aceast proprietate se bazeaz capacitatea anticariogen a fluorului), inhib sinteza de polizaharide intracelulare efectuat de bacterii, inhib enzimele Mg+2 - dependente formnd un complex insolubil de Mg+2 - fosfat - fluor. 2.4.3. Procesele metabolice din placa dentar La nivelul plcii dentare au loc multiple i variate procese metabolice, perfect controlate, care au ca rezultat fie constituirea plcii, fie degradarea ei. Astfel, din anabolismul glucidic rezult polizaharide extracelulare, ce intr n componena matricei, ct i polizaharide intracelulare, ce asigur materialul nutritiv pentru microorganisme. Microorganismele ce populeaz placa produc polizaharide din zaharurile simple aflate n mediul bucal. Dezvoltarea microorganismelor n placa dentar este dependent de aportul glucidic din mediul bucal. Procesele catabolice ale glucidelor i proteinelor din structura plcii au ca rezultat degradarea plcii dentare. De asemenea, microorganismele catabolizeaz nsi polizaharidele intraextracelulare, produse de bacterii n cursul activitaii anabolice. Prin catabolismul glucidic, pe calea glicolitica pn la acid lactic, se produce acidifierea local mediului, ceea ce are semnificaie etiologic n apariia cariilor. Acidul lactic rezultat poate fi preluat de alte microorganisme, care l degradeaz. Creterea nivelului de acid lactic declaneaz mecanismele de feed-back negativ care limiteaz producerea local de noi cantiti de acid lactic. Din catabolismul proteic n placa dentar, rezult uree care este convertit n amoniac i astfel mediul plcii se alcalinizeaz, pH-ul crete. n aceste condiii se favorizeaz remineralizarea smalului i formarea tartrului dentar. pH-ul local din placa dentar este dependent de proporia dintre microorganismele productoare de acizi, respectiv, cele ce catabolizeaz proteinele. 2.4.4. Rolul plcii dentare n declanarea unor stri patologice orale Placa dentar este factorul comun n etiologia cariei i parodontopatiei. Capacitatea plcii dentare de a contribui la producerea cariilor depinde de coninutul ei n calciu i fosfat, de metabolismul bacterian propriu, de limitele valorilor ntre care poate varia pH-ul su, de accesibilitatea i componenii salivei. Predominant i decisiv pentru etiologia cariilor este metabolismul desfurat n plac sub aciunea enzimelor bacteriene proprii. Atacul cariogen rezult din interaciunea dintre bacteriile orale, hran i smal. Bacteriile care pot adera efectiv la suprafaa dinilor
18 i pot produce cantiti apreciabile de acid, vor cauza demineralizarea smalului. Bacteriile care populeaz cavitatea bucal constituie o comunitate ce cuprinde diferite specii, iar studiul interaciunii lor cu esuturile cavitii orale este necesar pentru inelegerea biologiei orale la omul snatos, ct i la omul bolnav. Cavitatea bucal are o temperatur constant, iar alimentaia i fluxul salivar asigur materia prim, condiiile ionice i de pH necesare supravieuirii i multiplicrii bacteriene, realizndu-se astfel, o cultur mixt bacterian. Bacteriile, n schimb, modific compoziia fizico-chimic a mediului bucal, stabilesc legturi att cu gazda, ct i cu celelalte specii bacteriene. Cavitatea bucal a nou-nscutului este lipsit de microorganisme, dar n scurt timp se produce colonizarea bacterian, care coincide, de obicei, cu prima mas. Bacteriile colonizatoare ale cavitaii bucale au urmatoarele proprieti: obin hrana din saliv i diet; i tolereaz variaiile fizice ale mediului: modificrile de pH, modificrile presiunii pariale a oxigenului i a concentraiei ionilor; rezist la mecanismele de aprare: lizozim, transferin, sistemul peroxidazic, anticorpi; rezist la alte bacterii; ader la suprafee. Principalele microorganisme din plac i carie sunt steptococii mutans (70%) care fermenteaz zahrul i alte carbohidrate pn la acizi organici, scznd pH-ul la 4-5, sunt deci acidogenici, produc polizaharide intra i extracelulare, Lactobacillus acidophilus este acidogenic, produce acid lactic, se gsete n cantitate redus n placa dentar, dar predomin n leziunile de carie active. Exist o relaie strns dintre placa dentar i bolile parodoniului. In placa de 48 ore se produce material toxic n concentraii suficiente s afecteze integritatea gingiilor. Substanele din plac (enzimele proteolitice de origine bacterian, amoniacul, hidrogenul sulfurat, etc.) pot difuza n esuturile gingivale fiind iritani poteniali pentru gingie. De asemenea, endotoxinele bacteriene ptrund n gingie inducnd formarea de anticorpi. Reaciile locale antigen-anticorp, activeaz complementul iniiind procesul inflamator al gingiei i parodoniului.
La scurt timp dup constituirea plcii dentare ncepe procesul de mineralizare (calcifiere) al plcii, rezultnd tartrul dentar. Deci,
2.5. TARTRUL DENTAR
19 tartrul dentar este placa dentar mineralizat. 2.5.1. Principalele componente ale tartrului dentar Compoziia tartrului dentar variaz cu poziia, vechimea depozitului, mediul geografic i cu individul. Tartrul este format din substane anorganice (80%) i organice (20%). Componentele anorganice cuprind n principal, fosfat de calciu cristalizat sub form de hidroxiapatit, brushit, etc. (tabelul 3). Tabelul 3. Varietile de fosfat de calciu cristalizat din structura tartrului dentar (dup AURORA POPESCU, 1992, modificat) Variaii cu a e vrsta % % 43,6 8,9 scade 94,8 20 nu se modific 80,7 24,2 crete 99,5 55,3 nu se modific Cristalele de apatit apar n special n matricea extracelular a plcii, dar se gsesc i ntre celulele bacteriane, mpreun cu substanele organice. Bastonaele de brushit sunt rspndite extracelular, fiind separate de substanele organice. Componentele organice sunt reprezentate de proteine, polizaharide complexe, amino-acizi, acizi grai, fosfolipide, colesterol. Cristalizarea fosfatului de calciu din tartrul dentar se face pe o matrice ce are o compoziie asemntoare cu cea a plcii dentare. Astfel, n matricea tartrului se gsesc proteine mari ce conin 12- 20% zaharuri incluznd hexoze, fucoz, hexozamine i glicozaminoglicam (acetia din urm se presupune c i au originea n gingie, neexistnd n placa dentar). Att matricea plcii dentare ct i a tartrului dentar, sunt constituite din dou faze: una solubil i alta insolubil. Se remarc, c prin transformarea plcii n tartru dentar scad componentele organice de natur glucidic din matrice i crete concentraia proteinelor (tabelul 4). Tabelul 4. Repartiia procentual a componentelor proteice i glucidice din matricea plcii dentare i a tartrului dentar (dup AURORA POPESCU, 1992, modificat) Compon enta Proteine Hexoze Faza solubil Placa Tartrul dentar dentar 30 30 10 13,4 Faza insolubil Placa Tartrul dentar dentar 35 65 20 6 Varietatea de fosfat de calciu cristalizat Brushit Fosfat octocalcic Witlochita Apatit Inciden Cantitat
20 Fucoz Hexoza mine 1,6 3,5 3,3 0,2 4 9 2 2
De asemenea, tartrul dentar poate conine celule descuamate, leucocite, microorganisme Gram-pozitive i Gram-negative. 2.5.2. Formarea tartrului dentar Tartrul se deosebete de placa dentar prin faptul c are, n structura sa, fosfat de calciu cristalizat. Depunerea cristalelor de hidroxiapatit se face n jurul i ntre bacteriile filamentoase. Depunerea tartrului ncepe n primele zile de la constituirea plcii i este bine reprezentat n placa de 12 zile. Tartrul dentar se realizeaz prin dou mecanisme: mecanismul enzimatic, la care particip enzimele salivare, n special, fosfatazele, care perturb echilibrul soluiei coloidale a fluidului bucal, cu precipitarea srurilor minerale; mecanismul fizico-chimic, explic formarea tartrului ca un proces datorat stagnrii i alcalinizrii salivei. Ritmul de depunere al tartrului dentar este de 0,10-0,15 % din masa sa pe zi. Dup poziia i sediul depozitelor, tartrul dentar poate fi localizat supragingival i subgingival. Tartrul supragingival este mai abundent pe dinii din vecintatea carunculelor salivare, mai ales pe faa lingual a frontalilor inferiori i pe faa vestibular a molarilor superiori. Ocazional, tatrul dentar se poate depune i pe faa ocluzal a dinilor fr antagoniti. Tartrul supragingival este denumit salivar, deoarece saliva reprezint sursa srurilor minerale. Tartrul supragingival se prezint ca o mas friabil, care se poate desprinde usor, este de culoare alb-glbuie. Tartrul subgingival se depune pe feele care delimiteaz anul gingival, sub marginea gingival. Tartrul subgingival este mai dur i mai dens dect cel supragingival. Tartrul subgingival este denumit seric, deoarece, srurile minerale provin din saliv i lichidul crevicular. Tartrul subgingival are culoare verzuie, brun sau chiar neagr. Diferenele de culoare dintre tartrul supragingival i subgingival sunt datorate diferenelor de compoziie, direct dependente de variaiile compoziionale ale salivei, lichidului crevicular i de populaiile bacteriene. Depunerea tartrului dentar debuteaz n copilrie, n jurul vrstei de 10 ani, crescnd proporional cu vrsta. Depozitele de tartru dentar sunt foarte frecvente la aduli. Factorii favorizani ai transformrii plcii dentare n tartru sunt:
21 ionii de calciu i fosfat n concentraie mare din plac; pH-ul alcalin local; scderea componentelor organice de natura glucidic din matrice i creterea concentraiei proteinelor; creterea amoniacului i ureei n saliv ct i n placa dentar; reducerea n saliv a fosfatazei alcaline, enzim ce inhib formarea fosfatului de calciu cristalizat. Deci, n lipsa inhibitorului calcifierii, tartrul se formeaz mai uor. formarea brushitei, prima substana mineral, cristalin a tartrului ce constituie nucleul de cristalizare ulterioar a fosfatului de calciu. Placa dentar asociat cu tartrul dentar constituie cauze inflamatorii majore pentru parodoniu i gingie
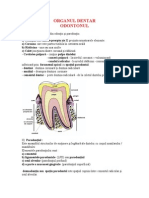
3.1. DEZVOLTAREA DINILOR
Dinii sunt formai din esuturi calcifiate i au dubl origine: ectodermal (ce va forma adamantoblastele i smalul) i mezodermal din care se vor diferenia: odontoblastele (formatoare de dentin), pulpa dentar, cementul. Organul dentar trece n cursul dezvoltrii sale prin mai multe etape ce cuprind: creterea i diferenierea celular adic proliferarea epitelial, diferenierea tisular, organogeneza, mineralizarea esuturilor dentare (calcifierea), erupia dentar nsoit de creterea radicular, uzura dentar i atrofia orizontal a parodoniului. 3.1.1. Creterea dinilor La embrionul de 6-7 sptmni, epiteliul bucal gingival se ngroas prin diviziuni rapide, se infund n esutul mezenchinal subiacent formnd creasta sau lama dentar primitiv, care va forma dinii temporari i din care prin proliferri succesive se va dezvolta i lama dentar secundar (d natere dinilor permaneni). Pe lamele dentare se formeaz cte 10 muguri (noduri) dentari. Din aceti muguri, prin procese de histodifereniere n epiteliu i esutul conjunctiv, ia natere o structur n form de clopot cu deschiderea spre maxilar. Partea epitelial va forma organul smalului (organul adamantin) n care se difereniaz dou straturi distincte morfo-fimctional:
22 zona epitelial exterioar sau stratul epitelial adamantin extern format din celule rotunjite sau cuboide; zona epitelial interioar a clopotului sau epiteliul adamantin intern alctuit din celule alungite, poliedrice numite adamantoblaste (ameloblaste), dispuse perpendicular pe papila conjunctiv. Fiecare celul are cte o prelungire fin, protoplasmatic ctre papila dentar. ntre celule se gsete substana fundamental bogat n elemente fibrilare, ce trec de la celul la celul. Dincolo de polul celular ce cuprinde nucleul, se ntinde un alt strat de celule cuboide ce formeaz stratul intermediar, bogat n fosfataze, cu rol de mineralizare a smalului. ntre stratul intermediar i epiteliul adamantin extern se gsete o substan fundamental cu caracter mucoid. Zona de la marginea clopotului, n care se face trecerea de la epiteliul extern la cel intern, are rolul inductor pentru formarea rdcinii. Mezodermul format din esut conjunctiv vascular se invagineaz n interiorul clopotului formnd papila dentar primitiv (papila conjunctiv) din care se vor diferenia: organul formativ al dentinei i schia pulpei dentare definitive. Papila conjunctiva este bogat vascularizat. Celulele situate la periferia papilei conjunctive se difereniaz n celule speciale, alungite, dispuse n palisad, celule numite odontoblaste. Acestea apar dup diferenierea ameloblastelor. Odontoblastele emit prelungiri distale, spre ameloblaste, prelungiri ce formeaz o reea bogat din care deriv fibrele Tomes. Odontoblastele au rol n formarea dentinei, conin fosfataz alcalin i transport material mineral. Epiteliul clopotului i papila dentar sunt nvelite de sacul dentar primar formnd mugurele (foliculul) dentar n care ncepnd din luna a V-a intrauterin, ncep s se formeze esuturile dure ale dintelui i aparatului de fixare dinte-alveol. 3.1.1.1. Formarea coroanei necesit diferenierea i proliferrea celulelor capabile s sintetizeze matricea proteic dur a smalului, dentinei i s o impregneze cu sruri de calciu. Formarea dentinei se realizeaz pe baza reelei fibrilare (predentina) ce provine din pulpa dentar primitiv i conine precolagen care trece apoi n colagen. n acest mas rmn canalicule fine prin care trec prelungirile odontoblastelor (fibre Tomes). Depunerile srurilor minerale au loc n jurul fibrelor Tomes (au rol n schimburile metabolice) iar predentina devine dentin. Depunerile de dentin se fac n straturi concentrice pornind de la
23 exterior ctre pulp i pe msur ce procesul nainteaz odontoblastele se retrag rmnnd numai fibrele Tomes inserate n canaliculii dentari. Dentina induce formarea matricei smalului. Formarea smalului se realizeaz de ctre ameloblaste dup ce sa depus primul strat de dentin. Funciile ameloblastelor sunt: formarea tiparului de coroan, organizarea odontoblastelor, formarea matricei smalului. Stadiile formrii smalului sunt: stadiul secretor, de producers a matricei cnd ameloblastele devin active, i dezvolt reticulul endoplasmatic, sintetizeaz proteine, membrana distal se plicatureaz, ntre pliuri existnd colagen ce va fi incorporat n stratul intern al smalului la jonciunea amelo-dentinal; stadiul de reorganizare n direcie longitudinal al organitelor celulare, care se produce dup formarea matricei, cnd celulele sunt mai mici iar structurile fibrilare din colagen se detaeaz uor de celul; stadiul de preabsorbie, caracterizat prin poziia central a nucleului i numeroase vezicule cu material mineral la jonciunea ameloblastelor cu celulele din stratul intermediar; maturarea timpurie, cnd veziculele de la captul bazal al amelo blastelor au migrat spre captul distal i depun cristalele de minerale paralel cu fibrele formate n primul stadiu. In stratul extern al smalului se gsete fosfataz alcalin care are rol n mineralizarea smalului; maturarea tardiv a matricei smalului se realizeaz cu participarea att a ameloblastelor ct i a odontoblastelor, care au proprietatea de a absorbi apa i materialul anorganic i de a secreta mineralele care intr n matrice. 3.1.1.2. Formarea rdcinii are loc dup apariia coroanei i se dezvolt la locul de unire a stratului extern-intern al organului smalului, prin creterea epiteliului n profunzime, ca o teac. Creterea n lungime a rdcinii se realizeaz prin prelungirea tecii epiteliale, apariia odontoblastelor i formarea dentinei. Vrful rdcinii se ngusteaz prin depunere de dentin, rmnnd
24 orificiul apical ce face legtura cu pulpa dintelui. Din sacul dentar se diferentiaz: - elemente celulare (cementoblaste) ce formeaza o varietate de tesut osos numit cement, asigurnd ancorarea fibrelor de membran periodontal i elemente fibrilare care vor intra n alctuirea parodoniului fixnduse pe osul alveolar i pe cement (fibre Sharpey). Formarea osului alveolar: din osul maxilar format n profunzime, o poriune nconjoar regiunea mugurilor dentari, apoi apar septuri interdentare, iar fibrele Sharpey fac legtura dintre cement i os realiznd ansamblul anatomofuncional osteo-dentar. La nceput germenii dinilor de lapte i ai celor permaneni sunt aezai ntr-o singur cavitate (alveola comun) dup care se separ alveola dinilor temporari i ai celor definitivi. Dezvoltarea dinilor permaneni - se produce din lama dentar secundar difereniat nc n viaa intrauterin. Pe lama dentar secundar, din mugurii dentari, se formeaz ncet dinii permaneni prin aceleai etape ca i cei temporari, n primii 6-20 ani de viaa. Cnd fiecare dinte permanent s-a format mpinge prin osul alveolar i determin erodarea rdcinii dintelui de lapte (rizaliz) cu cderea lui. La scurt timp dup acesta erupe dintele permanent. Dezvoltarea dentiiei ideale depinde de: formarea complet a dinilor, dezvoltarea structural normal a esuturilor dentare, erupia fiecrui grup de dini la timpul corespunztor, n spaiul adecvat i n relaie corect cu ceilali dini.
3.2. MINERALIZAREA DINILOR
Calcifierea esuturilor dure ale dinilor se face n general prin aceleai mecanisme ca la os, dar cu unele particulariti. Procesul de calcifiere cuprinde mai multe etape: formarea matricei proteice din monomer de colagen care polimerizeaz rapid formnd fibrele de colagen, ce au afinitate pentru srurile de Ca++; secretarea de substane care neutralizeaz pirofosfatul ce mpiedic cristalizarea hidroxiapatitei pe colagen; fosfataza alcalin din odontoblaste crete concentraia local a fosfatului anorganic i activeaz fibrele de colagen favoriznd
25 depunerea srurilor de calciu; precipitarea srurilor de calciu formnd o mixtur ce cuprinde: CaHPO4.2H2O, Ca3(PO4)2.3H2O i care prin substituie, adiie de atomi, prin reabsorbie i reprecipitare este convertit n cristale de hidroxiapatit. Particularitile calcifierii n esuturile dentare: n dentin cristalele de hidroxiapatit sunt mai dense dect n os i se depun pe fibre de colagen. Spre deosebire de os, dentina nu conine osteoblaste, osteocite, osteoclaste i nici spaii pentru vasele sanguine sau nervi. Dentina este depus i hrnit de odontoblaste. Srurile de calciu din dentin o fac foarte rezistent la forele de compresiune iar fibrele de colagen la forele de tensiune. Odontroblastele prin prelungirile lor n canaliculii dentari pot absorbi sruri i depune noi substane minerale n dentin: smalul este format din cristale foarte mari i dense de hidroxiapatit ce au adsorbit ioni de carbonat, magneziu, sodiu, potasiu, fluor. Depunerea srurilor minerale se face ntr-o reea proteic puternic i aproape complet insolubil. Din punct de vedere fizic aceste fibre proteice sunt similare keratinei din pr; schimburile minerale n dini: - se pot depune noi sruri n dentin, cement dac cele vechi au fost resorbite. Rata absorbiei i depunerii de minerale n cement este egal cu a osului alveolar iar n dentin rata este de numai 1/3 din a osului. Cementul are structura aproape identic cu a osului, inclusiv prezena osteoblastelor i osteoclastelor (particip la resorbia osului). Schimburile de minerale la nivelul smalului se fac foarte ncet, practic smalul i pstreaz aceeai compoziie toat viaa. Smalul poate prelua unele minerale din saliv (fluor, calciu).
3.3. ERUPIA DINILOR
Erupia dinilor reprezint micarea dinilor prin esuturile gingivale spre cavitatea oral. Erupia este diferit de noiunea de
26 cretere a dinilor, care poate fi un factor cauzal al erupiei. Suprafaa masticatorie presnd, coroana perforeaz gingia, acesta constituind primul moment al nceputului erupiei. Epiteliul cavitii bucale i sacului dentar se unesc. Din resturile sacului dentar se formeaz marginea gingival i papila interdentar, iar prinderea sacului dentar n dreptul gtului dentar va forma ligamentul circular. Deplasarea dintelui spre cavitatea bucal se consider terminate, cnd dintele a luat contact ocluzal cu cel antagonist. Dup atingerea acestui plan ascensiunea dintelui continu, dar este frnat de ocluzie i compensate cu uzura suprafeelor de contact ocluzal. Erupia dinilor se nsoete de hipersalivaie, agitaie, tumefierea mucoasei gingivale. 3.3.1. Mecanismele erupiei dinilor Teoriile privind erupia dinilor se bazeaz pe: observaii clinice pe subieci umani prezentnd anomalii de erupie, studii radiologice i histologice la subieci normali i pe observaii experimetale la animale. Pe baza informaiilor culese s-au elaborat urmatoarele mecanisme posibile, implicate n erupia dinilor:
27 creterea rdcinii dintelui care mpingnd osul alveolar foreaz coroana spre cavitatea bucal; construcia pulpei: s-a sugerat c prin creterea odontoblastelor i a grosimii dentinei se micoreaz cavitatea pulpar crescnd presiunea n interiorul cavitii ceea ce ar determina mpingerea dintelui spre cavitatea bucal. n deficit de vitamina A apare o cretere neobinuit a dentinei, vasele dentare sunt puine la numr, cavitatea pulpar este micorat, iar rata erupiei este redus; creterea pulpei: Sicher sugereaz c creterea pulpei asociat cu creterea rdcinii particip la realizarea "forei eruptive" a dintelui. La captul apical al pulpei celulele sunt n diviziune rapid, se formeaz noi fibre de colagen ce realizeaz un adevarat ligament "like-hamac" care este ancorat pe osul alveolar i nconjoar vrful rdcinii, tracionnd astfel dintele spre cavitatea bucal; creterea osului alveolar mpinge dintele printre fibrele periodontale. La dinii multiradiculari, osul dintre rdcini prolifereaz foarte activ n timpul erupiei realiznd o presiune ce foreaz dintele spre cavitatea bucal; vascularizaia bogat din regiunea apical determin formarea de lichid interstiial ntr-un spaiu restrns ceea ce exercit o presiune contribuind la erupia dinlilor. Creterea sau scderea fluxului sanguin apical produce Modificri corespunzatoare n cantitatea de hormoni, sruri minerale i factori nutritivi ce ajung la dinte iar acetia afectnd creterea tisular pot altera rata erupiei. Bryer a gsit experimental la roztoare c scderea vascularizaiei reduce rata erupiei, iar edemul pulpar crete rata erupiei. Hormonii tiroidieni i somatotropul cresc vascularizaia pulpei i deci grbesc erupia dinilor; ligamenlul periodontal poate participa i el la fora eruptiv n viaa embrionar la nivelul lamei dentare apare, de asemenea, un organ formator al dinilor definitivi (muguri dentari secundari) ce trec prin aceleai faze de dezvoltare ca i dinii temporari. Cnd rdcina definifiv ncepe s creasc, are loc resorbia rdcinii dinilor temporari, creind spaiul necesar. Resorbia sau rizaliza fiziologic este un proces autonom, se desfoar cu precizie la diferite vrste determinnd n final cderea dintelui temporar. Resorbia ncepe la un an de la creterea complet a dintelui de lapte. Osteoclastele devin odontoclaste ce particip la resorbia rdcinii dinilor temporari. Traumatismul cauzat de stressul ocluzal i procesele inflamatorii grbesc resorbia rdcinii dinilor de lapte. 3.3.2. Calendarul erupiei dinilor Erupia dinilor temporari se desfoar n urmtoarea ordine: la 6-10 luni incisivii, la 12-16 luni I-ul molar, la 16-20 luni caninii, la 2030 luni molarii secunzi (II-lea molar). Erupia definitiv se desfoar aproximativ n ordinea urmatoare:
28 la 6-7 ani apar primii molari, ntre 7-9 ani incisivii, 9-11 ani caninii, 10-12 ani premo-larii, la 6-12 ani molarii secunzi, la 16-20 ani erup molarii de minte (al III-lea molar) (tabelul 1). 3.3.3. Principalii factori ce infueneaz erupia dentar Dintre factorii ce controleaz erupia amintim civa: mineralizarea i erupia dinilor este mai timpurie la fete dect la biei. Aceast diferen s-a observat nainte de pubertate, deci ar fi controlat de cromozomii sexuali i mai puin de hormonii sexuali. Mineralizarea i erupia este de 3-5% mai repede la fete dect la biei; erupia tinde s fie mai precoce la grupurile socio-economice mai elevate; factorii genetici, de ras influeneaz erupia: la negrii erupia dinilor permaneni este cu 7 sptmni mai devreme dect la albi, cu aceei stare economic; hormonii tiroidieni, hormonul somatotrop cresc rata erupiei, etc. 3.3.4. Tulburri de erupie Tulburrile de erupie ale dinilor pot avea cauze variate, i anume: erupia ntrziat a dinilor asociat cu boli endocrino-metabolice: cretinism, rahitisin; erupia dinilor permaneni n poziia lor normal poate fi influenat de: lipsa spaiului, poziia anormal a criptei, dini suplimentari sau supranumerari, retenia dinilor de lapte care pot fi anchilozai n os; perturbri severe ale nutriiei i metabolismului ntlnite n gastroenterita infantil, hipotiroidism idiopatic, hipovitaminoza D; boli genetice: sindromul Down se nsoete de fisuri ale buzelor sau palatului n 1 din 200 pacieni, iar n zona fisurii dinii sunt abseni sau boli genetice: sindromul Down se nsoete de fisuri ale buzelor sau palatului n 1 din 200 pacieni, iar n zona fisurii dinii sunt abseni sau malformai; chimioterapia cu citostatice poate determin hipoplazia coroanei i smalului, iar rdcinile dinilor sunt scurte
29
belul 5. radiculare Rdcina complet format nceputul resorbiei 4-5 a4-5 a6-7 a4-5 a4-5 a
30
ia
Dinii Temporari
-20 l8-10 l6-8 l
3 a2-2,5 a2,5-3 a1,5-2 a1,5-2 a
10-11 a11-12 a8-9 a7-8 a 18-25 a14-16 a9-10 a12-14 a12-13 a13-15 a11 a10 a
superioriPermaneni
10-11 a11-12 a8-9 a7-8 a 18-25 a14-15 a9-10 a13-14 a12-13 a12-14 a10 a9 a
inferioriPermaneni
rine; s I-u = sptmna de via intrauterine
31
boli genetice: sindromul Down se nsoete de fisuri ale buzelor sau palatului n 1 din 200 pacieni, iar n zona fisurii dinii sunt abseni sau malformai; chimioterapia cu citostatice poate determin hipoplazia coroanei i smalului, iar rdcinile dinilor sunt scurte. 3.4. STRUCTURA MORFO-FUNCIONAL A DINTELUI Totalitatea dinilor fixai pe cele dou arcade formeaz dentiia. n cursul vieii la om sunt dou dentiii: temporar, "de lapte", alctuit din 20 de dini, apar ntre 6 luni i 2 ani i jumtate de via persistnd pn la 6-13 ani; permanent, definitiv, alcatuit din 28-32 dini ce apar ntre 6-18 ani, al treilea molar (de minte) poate apare ntre 17-35 ani. n practica morfogenetic dentaia se exprim prin "formula dentar" ce reprezint repartiia i numrul dinilor pe fiecare jumtate de maxilar i mandibul folosindu-se simbolurile: I = incisivi, C = canini, P = premolar, M = molar, numrtorul fraciilor indicnd dinii superiori, iar numitorul pe cei inferiori. Formula dentar este pentru dentiia de lapte: I = 2/2, C = 1/1, M = 2/2, iar pentru dentiia permanent: I = 2/2, C = 1/1, P = 2/2, M = 3/3. Dinii ca parte integrant a sistemului dento-maxilar intervin n prehensiunea, tierea, sfrmarea i masticarea alimentelor (n vederea realizrii bolului alimentar), ct i n actul fonaiei-vorbirii.
32 Descriptiv dintele prezint o poriune vizibil n cavitatea bucal (coroana) i o poriune fixat n alveola, ce se subiaza spre profunzime (rdcina). ntre coroan i rdcin exist un an limitat, coletul sau gtul dintelui (fig. 6). Alimentaia variat a determinat formarea de patru tipuri de dini cu forme i funcii particulare. Incisivii sunt monoradiculari, au coroana. n form de dalt, realizeaz prin ntlnirea suprafeelor ocluzale fragmentarea prin tierea alimentelor. Caninii sunt cei mai putenici dini monoradiculari, forma ascuit a coroanei realizeaz perforarea i sfierea hranei la micrile de lateralitate. Premolarii i molarii care au suprafee mari de contact ale coroanei asigur strivirea i triturarea hranei ntre cuspizi (proeminene ce ptrund n adnciturile dintelui din partea opus), prin micrile de lateralitate i circumducie realiznd transformarea mecanic a alimentelor. Funcional dintele este format din esuturi proprii dentare (odoniul) ce cuprinde emailul (smalul dentar), dentina, cementul, pulpa i esuturile de fixare n alveol (parodoniul).
3.4.1. Smalul dentar (emailul) Smalul dentar este produs de celulele epiteliale specializate numite amelo-blaste nainte de erupia dintelui. Este cel mai dur esut din organismul uman, conine substane anorganice 96%, substane organice 1,7%, ap 2,3%,. Din substanele anorganice 90% reprezint fosfatul de calciu, 5% carbonatul de calciu, 2% fosfatul de magneziu, 3% microelemente (fluorul n principal). Fosfatul de calciu se afl cristalizat sub form de hidroxiapatit fixat ntr-o reea foarte fin de fibre proteice, complet insolubil, fibre care sunt similare cu keratina din pr. Fibrele proteice din email sunt rezistente la acizi, enzime, ali ageni corozivi, fibrele de reticulin din smal se continu cu cele din dentin, iar prelungirile dentinei (fibrele Tomes) se termin n smal. Smalul este lipsit de vase de snge i nervi. Are un metabolism propriu lent, este comparat cu o membran semipermeabil ce oprete ptrunderea florei microbiene bucale spre pulp, dar are o capacitate de preluare din saliv a fluorului, calciului, fosforului mbogindu-i astfel coninutul cu 1% n 250 zile. 3.4.2. Dentina Dentina delimiteaz cavitatea pulpar, are duritate mai mic dect smalul, conine 70% substane anorganice, 20% substane organice, 10% ap. Este format din cristale de hidroxiapatit similare celor din os dar mult mai dense ce i confer rezistent la forele de compresiune i din fibre de colagen ce i confer rezistena la forele tensionale care pot rezulta cnd dinii zdrobesc obiecte solide. Dentina este depus i hrnit de stratul de celule numite
33 odontoblaste care au suprafaa lor intern de-a lungul pereilor cavitaii pulpare. Dentina se depune continuu, are un metabolism lent, este strbtut de numeroase canalicule radiare n care se gsesc prelungirile odontoblastelor din pulpa dentar. ntre prelungiri i canalicule circul limfa dentar care asigur nutriia dentinei. 3.4.3. Pulpa dentar Pulpa dentar este plasat n camera central a dintelui, este format din esut conjuctiv lax aderent la pereii cavitii, are o bogat vascularizaie i inervaie. esutul conjunctiv lax cuprinde: fibre de colagen i reticulin ce asigur: susinerea componentelor pulpare, amortizarea forelor exercitate din direcia dentino-pulpar, suportul direcional al mineralizrii dentinei n odontogenez i al dentinei secundare; elemente celulare reprezentate de: fibroblati (au rol activ n procesele reparatorii pulpare, sintetizeaz precursorii mucopolizaharidelor i colagenului), celule mezenchinale nedifereniate (reprezint elementul celular de rezerv i asigur aprarea nespecific prin fagocitoz), odontoblaste (principala funcie a lor reprezint producerea dentinei, i trimit prelungiri n canaliculele radiare ale dentinei asigurnd astfel nutriia i schimburile de sruri minerale la nivelul dentinei). Cu vrsta celulele ce delimiteaz cavitatea pulpar se extind micornd astfel cavitatea. Vascularizaia extrem de complex este asigurat de o arteriol ce strbate orificiul apical i formeaz o dubl reea de capilare, unele cu rol nutritiv, iar altele cu rol funcional (acioneaz ca o supap prin lrgirea lumenului, n caz de cretere a presiunii sangelui la marginea cavitatii). Presiunea hidrostatica din capilare, ce reprezint un factor activ al schimburilor nutritive, este modulat prin raportul diametrelor vaselor aferent-eferent, de ctre factorii metabolici locali i fibre nervoase vegetative. Inervaia pulpei este asigurat de fibre senzitive cu origine n ganglionul Gasser i de nervii mandibulari, maxilari ce ptrund odat cu vasele i se ramific formnd un plex spre periferia pulpei. Din acest plex fibre amielinice ptrund n canalele dentinare. Fibrele vegetative simpatice provin din plexul carotidian i cele parasimpatice din ganglionul sfenopalatin i otic. 3.4.4. Cementul Cementul reprezint stratul de nveli al rdcinii dintelui, este o varietate de esut osos, este secretat de celulele membranei
34 periodontale. Este puternic calcificat (55% substane minerale), conine geode n care se afl cementociii este hrnit de vasele periodontale i pulpare prin intermediul dentinei. Matricea organic cuprinde colagen tip I i proteoglicani. Colagenul din cement are dou surse: fibrele intrinseci sunt secretate de cementoblaste, iar fibrele extrinseci sunt capetele fibrelor ligamentului periodontal inserate n cement. Cementul celular conine cementocite (nglobate n structura sa) i se gsete n prima treime inferioar pn la jumtatea rdcinii dintelui, iar cementul acelular este localizat de la jonciunea amelocemental pn la orificiul apical (fig. 7).
Cementogeneza: presupune diferenierea cementoblastelor care secret precementul, fibre intrinseci de colagen, proteoglicani pe suprafaa predentinei din rdcina dintelui, iar pe msur ce predentina este mineralizat, creterea cristalelor depete jonciunea cementodentinal continund n precement. Rata depunerii cementului variaz considerabil n funcie de vrsta dintelui j locul de depunere. Cnd depunerea cementului
35 este lent, se formeaz un strat fin de cement cu puine fibre intrinseci. n timpul depunerii rapide a cementului, n regiunea apical, crete proporia de fibre intrinseci, cementul este mai neregulat i are o grosime mai mare, iar cementoblastele sunt complet nconjurate de matricea extracelulara secretat de celelalte cementoblaste, rezultnd cementul celular. Depunerea excesiv de cement se numete hipercementoz i poate produce un apex al rdcinii bulbos, globular. Aceast neregularitate a apexului poate produce dificulti n extracia dintelui, cnd este necesar aceasta. n unele situaii depunerea excesiv a cementului celular, poate duce la fuzionarea rdcinii cu osul alveolar adiacent, determinnd anchiloza. Anchiloza are urmtoarele consecine: previne cderea dinilor de lapte, mpiedic erupia dinilor permaneni, cauzeaz dificulti la extracie. Hipercementoza se ntlnete n boala Paget sau n cementom (tumor benign a cementoblastelor). Funciile cementului: asigur inseria fibrelor ligamentului desmodontal, protejeaz rdcina dintelui la stressurile anormale, menine un spaiu periodontal constant prin depunerea permanent de cement care compenseaz liza osoas. Spaiul periodontal este spaiul dintre dinte i alveol. 3.4.5. Permeabilitatea esuturilor dentare i modificrile cu vrsta a esuturilor dentare Smalul i dentina dei au un metabolism sczut sunt nc esuturi n activitate. Pentru studiul permeabilitii dentinei s-au efectuat experiene pe dini n situ, la care s-au plasat n pulp, dentin, smal, colorani solubili i insolubili, (argirol, albastru tripan), substane marcate radioactiv. Rezultatele arat c dentina este permeabil dinspre pulp i dinspre jonciunea amelo-dentinal, permeabilitatea tinde s scad cu vrsta, dentina este mai permeabil chiar dect emailul tnr. Calea major de difuzie din pulp spre dentin este prin intermediul odontoblastelor ce i trimit prelungirile n dentin. Permeabilitatea smalului este mai redus dect a dentinei. Pentru studiul permeabilitii smalului s-au folosit substane administrate pe mai multe ci: albastru de metilen plasat n dentin la dinii umani, n situ, penetreaz rapid n pulp dar ptrunde puin n email acumulnduse la jonciunea amelodentinal. P32 administrat intravenos se regsete n cantiti mici n smal: cile de ptrundere ar fi: pulpa prin intermediul dentinei, saliva, aportul sanguin n membrana periodontal i cement via jonciunea cemento-dentinal i amelo-dentinal. Calciu, Zinc, Argint marcate radioactiv ptrund n dinte prin smalul defect dar nu i prin smalul intact. Smalul poate prelua din saliv substane (fluorul), crescndu-i astfel rezistena la carii. Cu vrsta apare o pigmentare gradat a dinilor, se reduce
36 volumul pulpei dentare, scad odontoblastele, fibroblastii fiind nlocuii cu esut fibros, scade numrul de vase pulpare iar cele rmase prezint fenomene de ateroscleroz cu diminuarea capacitii de hrnire a dintelui, pot apare mici calcifieri sau mineralizri difuze a pulpei. 3.5. COMPOZIIA CHIMIC A DINILOR Studiile privind compoziia chimic a dinilor s-au fcut mai ales din esuturi "uscate" nclzite la 105C i prin difracie cu raze X care permite studiul mrimii, aranjrii constituenilor din cristale. Tabelul 6. Compoziia aproximativ a smalului i dentinei (exprimat ca procente din greutatea uscat) Substane Substane H2O anorganice organice SMAL 96% 1,3-1,7% 2,3% DENTIN 70% 20% 10% 3.5.1. Substane anorganice: Cristalele de apatit sunt sarea dubl de fosfat de calciu i alte sruri de calciu aranjate astfel 3Ca3(PO4)2Ca X, n care X poate fi: (OH)2, F2, Ch, SO4-2, C03-2. Exemplu hidroxiapatita: 3Ca3(PO4)2 Ca(OH)2 sau Ca10(OH)2(PO4)6, care exprim mai bine radicalii prezeni i proporia lor relativ n hidroxiapatit. Fluorapatita: 3Ca3(PO4)2 CaF2. Raportul Ca/P n apatit este 2/15 probabil datorit unui exces de fosfat adsorbit pe cristale. O proprietate important a cristalului de apatit este schimbul ionic: schimb heteroionic, un ion Ca+2 poate fi nlocuit cu siliciu, natriu, mangan, stroniu, iar OH- (hidroxil) poate fi inlocuit cu Cl-, Fl-; schimb izaionic, un Ca+2 poate prsi cristalul i s fie nlocuit de alt Ca+2. Viteza de schimb a ionilor din stratul hidratat al cristalului de apatit este mare, pe cnd cea a ionilor din mijlocul cristalului este foarte mic. Amestec de CaCO3 i Ca3(PO4)2 Ali constitueni anorganici din email i dentin, se pot clasifica astfel: n interior (fluor, plumb, zinc, fier, stibiu); constitueni cu concentraii mai mari pe suprafaa emailului dect
constitueni cu concentraii mai mici pe suprafaa dect nuntru (Na+, Mg+2,C03-2) constitueni cu distribuie aproape uniform (stroniu, cupru, aluminiu potasiu). Na+, Cl", Se+, se gsesc n concentraie mai mare n email dect
37 n dentin. Carbonatul n concentraii mari crete solubilitatea emailului i poate fi un factor de scdere a rezistenei la carie. Se pare c fluorul nlocuiete carbonatul n cristalele emailului i crete rezistena la carie. Carbonatul din email este mai sczut pe suprafaa extern i mai crescut la jonciunea amelodentinal. Fluorul din dini depinde de fluorul ingerat prin hran, ap, ceai, mai ales n timpul calcifierii dinilor. Fluorul se gseste mai mult n dentin i pe suprafaa smalului. Fluorul este depus parial nainte de erupie i parial dup erupie fiind preluat din saliv i ceai. Fluorul n dentin crete odat cu vrsta. Fierul intr n structura pigmentului galben din incisivii roztoarelor. Zincul are distribuie similar cu fluorul. Solubilitatea hidroxiapatitei este mai redus prin administrarea de zinc dup erupia dinilor, dar nu s-a observat c ar reduce cariile, la animale. Compoziia smalului i dentinei este influenat de: tipul de dini (sunt mici diferene ntre dinii temporari i definitivi), vrsta (crete fluorul cu vrsta, scade permeabilitatea), prezena sau absena cariei (fluorul este mai crescut la dinii fr carie, iar cnd se instaleaz caria, fluorul se acumuleaz n leziune datorit smalului defect i mai permeabil). 3.5.2. Substane organice (Tabelul 7). Tabel Nr. 7 Repartiia substanelor organice n dentin i smal n dentin = 20,83 % din n smal = 1.3 1.7 % din greutatea uscat greutatea uscat colagen - 18 % colagen - urme lipide - 0.33 % lipide - 0.6 % citrat - 0.9 % citrat - 0.1 % matrice proteine - 1.6 % - 0.3 % necolagene insolubile proteine - 0.05 % solubile Colagenul este o protein fibrilar, se gsete n matricea organic a osului, dentinei, cementului, cartilajului, etc. n compoziia colagenului intr aminoacizi: glicina (1/3), alanina, prolina, hidroxiprolina (1/3) i ali 14 aminoacizi (I/ 3). Aminoacizii sunt aezai n triplu helix, n os, dentin, piele, 2 lanuri polipeptidice sunt identice, iar al treilea difer puin n compoziia aminoacizilor. Colagenul nou format se dizolv n acizi slabi. Colagenul este sintetizat n fibroblati, n osteoblaste, odontoblaste, pe poliribozoini. Asupra precursorilor solubili ai colagenului, ce conin la captul aminoterminal un grup de aminoacizi atipici, acioneaz enzima proteolitic procolagen-peptidaz care nltur aceti aminoacizi, rezultnd fibrele de colagen insolubil. In fibroblati se gsete fosfataza alcalin cu rol n sinteza unor proteine fibrilare cum ar fi
38 keratina. ACTH, glucocorticoizii inhib formarea colagenului, mineralcorticoizii stimuleaz fibroblatii i sinteza colagenului. Rata turnoverului colagenului este scazut n dentin, n schimb n ligamentul periodontal colagenul este renoit n cteva sptmni, datorit presiunilor mecanice ce apar n timpul masticaiei i colagenazei prezent n gingie i esutul periodontal. Fibroblatii din esutul periodontal au colagenaz preformat i particip la turnoverul colagenului prin capacitatea fagocitar. Deficitul de vitamina C scade sinteza colagenului. 3.6. FIZIOLOGIA PARODONIULUI 1 A GINGIEI Parodoniul este unitatea funcional a esuturilor de susinere a dintelui n alveola i care permite transmiterea solicitrilor mecanice la care este supus dintele spre osul alveolar i maxilar subiacent.. Structura i arhitectura parodoniului este dirijat de forele care pornesc de la dini ncepnd din timpul erupiei i se continu apoi n perioada funcional a aparatului masticator. Prin restructurarea i adaptarea continu a parodoniului n timpul vieii se asigur meninerea unui echilibru funcional ntre esuturile parodontale. Parodoniul cuprinde: esuturile de fixare i susinere reprezentate de cement, os alveolar, desmodoniu; esuturile de nveli reprezentate de mucoasa gingivala. 3.6.1. Cementul Cementul este considerat o variant de esut osos secretat de celulele membranei periodontale (cementoblaste). Acoper rdcina dintelui de la colet la apex, este puternic calcificat. Funciile cementului: ofer inserie pentru fibrele ligamentului desmo-dontal, protejeaz rdcina dintelui fat de stressurile anormale, menine un spaiu periodontal constant prin depunerea permanent de cement ce compenseaz liza osoas (vezi 3.4.4.). 3.6.2. Osul alveolar Osul alveolar se mai numete i os dentar datorit rapoartelor anatomice filogenetice i ontogenetice pe care le are cu dintele. Este format din esut spongios osos delimitat vestibular i oral de ctre o cortical. Conine lojele alveolare ce adpostesc i sprijin rdcinile dinilor, lojele sunt separate prin septuri interdentare. Pe suprafaa alveolei (lamina dur) se inser ligamentul periodontal. Lamina dur este perforat de orificii ce asigur trecerea vaselor nutritive, limfatice i a nervilor. Osul alveolar este ntr-o continu remaniere: osteoblastele produc matricea organic a osului alveolar care apoi se calcific, iar osteoclastele particip la rezorbia osului. ntre dinte i osul alveolar se gsete fisura periodontal de aproximativ 0,22-0,25 mm, ce asigur mobilitatea fiziologic a dintelui. 3.6.3. Desmodoniu Cuprinde esutul conjunctiv din fisura periodontal reprezentat de:
39 fascicole ligamentare din fibre de colagen ce leag dintele de os realiznd adevratele ligamente alveolo-dentare (fibrele Sharpey). Unele fascicule se pierd n grosimea mucoasei gingivale sau papilare (fibre marginale sau cemento-papilare); altele trec de la cementul unui dinte la altul, pe deasupra septurilor alveolare (fibre transseptale sau cementoceinentare); altele trec de la cement la peretele alveolar (cementoalveolare sau ligamentul alveolodentar). formaiuni vasculare ce formeaz numeroase plexuri. Vasele au rol nutritiv dar i rol funcional-mecanic. fibroblati ce particip la neoformarea fibrelor ligamentare i a substanei fundamentale; nervi i lichid intertisular. Toate acestea asigur un "hamac" pentru dinte. Rolurile desmodoniului: prin fibrele Sharpey asigur fixarea dintelui n alveol; transmite forele ocluzale la os prin grupul fibrelor oblice i astfel forele de presiune se transform n fore de traciune asupra cementului i osului alveolar (fig. 8); acioneaz ca o frn hidraulic: o parte din forele ocluzale sunt anihilate prin sistemul hidraulic asigurat de lichidul intertisular i vasele sanguine din regiune. n apropierea apexului capilarele prezint dilatri (glomeruli) cu rol de amortizare a presiunilor dentare. Prin reeaua nervoas vegetativ bogat din jurul dilatrilor vasculare se asigur reglarea automat i pe cale nervoas a fluxului sanguin local.
40
3.6.4. Mucoasa gingival Mucoasa gingival acoper procesul alveolar, ader intim de periost, protejeaz rdcina dintelui i esuturile de susinere, este moale, elastic, este bine vascularizat i inervat. La copil i la edentatul total, aceast mucoas este continu i ader intim de periostul subjacent, care i d aspectul de fixitate. n timpul erupiei dinilor temporari i permaneni, acest bandelet fibromucoas este perforat circular, lund forma unui stativ n care sunt aezate coroanele dentare. Se descriu trei poriuni ale mucoasei gingivale: gingia aderent acoper cementul i osul alveolar de care se fixeaz prin fascicole fibroase gingivo-periostale i gingivo-cementare; gingia marginal (gingia liber) reprezint poriunea din gingie ce nconjoar dintele la colet,jealiznd un adevarat "guler" elastic datorit sistemului de fibre de colagen. n fundul de sac gingival se secret lichidul anului gingival bogat n enzime i substane antimicrobiene (lizozim, IgA secretorie). Coninutul i compoziia lichidului anului gingival variaz n funcie de starea de sntate a parodoniului; -gingia interdentar cuprinde mucoasa gingival ntre doi dini vecini. Cu vrsta apar modificri histologice i funcionale ale gingiei: atrofie gingival, procese de keratinizare, scade consumul de oxigen
41 i activitatea metabolic. Mucoasa gingival se continu cu restul mucoasei bucale printr-o zona de tranziie, denumit mucoasa alveolar. Epiteliul acestei zone este subire, corionul este srac n papile, iar stratul subinucos este bogat n esut conjunctiv lax i n vase, ceea ce confer caracterul mobil al acestei formaii. Numai n regiunea palatinal, mucoasa alveolar este ferm, fix, deoarece la aceast zon ader intim de periost. Vascularizaia parodoniului este asigurat de arterele gingivale pentru mandibul i pentru maxilar. Arterele gingivale asigur o bogat irigaie a desmodoniului nainte de a ptrunde n canalul radicular. Inervaia este asigurat de nervii dentali, ramuri din trigemen. Funciile parodoniului i gingiei:
42 menin i fixeaz dintele n alveol (fibrele Sharpey); gingia marginal asigur protecia spaiului periodontal unde se gsete ligamentul desmodontal; vasele ce provin din osul alveolar i gingie hrnesc ligamentul desmodontal. ligamentul asigur stimularea fiziologic a metabolismului osului alveo lar prin transformarea forelor de presiune ocluzal n fore de traciune. Dinii suport variate fore mecanice n timpul masticaiei, vorbirii, deglutiiei i aceste stressuri se transmit osului alveolar pe calea ligamentului periodontal. Fibrele principale ale ligamentului au un anumit grad de laxitate, prelund forele masticatorii printr-un mecanism vascoelastic. Componentele macromoleculare din matricea extracelular a esutului conjunctiv sunt hidrofilice i leag apa, realiznd o "pern de ap" prin care presiunile din ligament se transmit la osul alveolar. Ligamentul este mentinut ntr-o stare continu de intindere (tensiune). prin inervaia bogat parodoniul este punctul de plecare a numeroase reflexe de reglare a poziiei mandibulei. Sensibilitatea parodoniului i mucoasei gingivale prin releul (circuitul) reflexelor parodonto-musculare, gingivo-musculare i respectiv gingivo-parodonto-musculare descrise de Rubinov, comand ntreruperea contraciei musculare n funcie de densitatea, duritatea alimentului i experiena acumulat de subject, presiunile masticatorii neatingnd intensitti dureroase, n condiii normale; esutul periodontal particip la generarea forei necesar erupiei dinilor; celulele macrofage, fibroblatii, cementoblastele i osteoblastele care sunt rspndite n ntreg parodoniul asigur acestuia funcia de aprare i refacere. esutul conjuntiv al parodoniului este constant renoit printr-un turnover Pasiv i printr-un proces numit remodelare (care apare cnd se produce alterarea funcional a ligamentului). Cele dou procese sunt continue i practic inseparabile. Rata turnoverului colagenului din ligamentul periodontal este mai mare dect n alte esuturi
43 conjunctive iar fibroblatii i matricea extracelular sunt n permanen reamplasai. Folosind aminoacizi marcai radioactivi s-a artat c sinteza colagenului i reorganizarea sa are loc n tot cuprinsul ligamentului, n timpul micrii dinilor. Cnd presiunile exercitate asupra dintelui determin micarea lui, osul este resorbit de pe suprafaa unde se exercit presiunea i este depus pe suprafaa unde se exercit tensiunea. Deci, se activeaz osteoclastele la locul presiunii i osteoblastele la locul tensiunii (fig. 8). Boala parodontal este consecina tulburrilor locale a metabolismului esuturilor de susinere a dintelui n alveola, cauzat de modificri genetice, endocrine, nutriionale, metabolice, acidobazice, etc. Gingia sufer leziuni mecanice prin contactul cu alimentele iar osul alveolar este continuu stressat n procesul de masticaie. Aceste efecte traumatizante necesit o permanent anihilare pentru a preveni leziunile parodoniului. Dar i masajul insuficient al esuturilor parodoniului, datorat dietei relativ moale a omului modern, poate produce parodontopatie.
esuturile sistemului stomatognat sunt foarte sensibile la carenele vitaminice, manifestrile bucale ale strilor de caren vitaminic apar precoce. 3.7.1. Vitamina A Rolul major al vitaminei A este n procesul vederii i n cretere. Vitamina A stimuleaz creterea prin aciunea sa asupra epiteliilor legat de formarea celulelor noi. n lipsa acestei vitamine diferenierea celular nu mai are loc iar posibilitatea de regenerare a celulelor dispare. esuturile aparatului dentomaxilar sunt deosebit de sensibile la carenele de vitamina A. Vitamina A are rol de protecie a epiteliului i intervine n meninerea normal a sistemului ectodermal. Deficitul de vitamina A determin: keratinizarea i descuamarea pielii i a mucoasei bucale, stomatit comisural, scderea secreiei de mucin, procese degenerative n glandele salivare cu diminuarea secreiei salivare, mucoasa bucal se usuc, se ngroas, se descuameaz devenind astfel permeabil pentru microorganisme. Apare infecia i inflamaia prilor moi, iar formarea tartrului dentar prezint un punct de plecare spre afeciuni parodontale. Dezvoltarea i implantarea dinilor, formarea smalului sunt profund afectate n carena de vitamina A ca urmare a modificarilor atrofice suferite de ameloblaste (celule de origine epitelial, responsabile cu formarea smalului). Cnd carenta de vitamina A survine la mama in perioada de alaptare, sau la copil, pana la vrsta de 6 ani, se produce o calcifiere defectuoas a smalului, a dentinei i a cementului. Apar hipoplazii dentare, atrofii ale organului smalului, predentin n cantitate mare tulburri n
3.7. VITAMINELE 1 SISTEMUL STOMATOGNAT
44 erupia dentar, anomalii de poziie i ntrzieri n dezvoltarea arcadelor alveolare. Prezena vitaminei A este indispensabil, ea avnd un rol maxim n perioada formrii dinilor. 3.7.2. Vitamina D Vitamina D este implicat n reglarea metabolismului fosfocalcic. Deficitul de vitamina D scade calcemia, prin diminuarea absorbiei intestinale i reducerea osteolizei, se declanseaz un hiperparatiroidism, secundar hipocalcemiei. La nivelul aparatului dentomaxilar modificrile cauzate de carena de vitamina D apar mai trziu i afecteaz mai ales maxilarul superior care este mai bogat vascularizat. Forele care acioneaz asupra maxilarului slab calcifiat i modific forma, acesta avnd o bolt palatin alungit, ngustat n dreptul premolarilor (bolta ogival). Prin apropierea rebordurilor alveolare pe linia median se produc modificri n articulare i n ocluzie, att n plan orizontal ct i vertical. n rahitism, dinii sunt afectai n proprorie de 70-80%, se constat ntrzierea erupiei nsoit de malpoziii, nanismul dinilor temporari, hipoplazia smalului (cauzat de afectarea ameloblastelor), suprafaa smalului este rugoas, pigmentat i mai bogat n substane organice. Dentina prezint numeroase zone slab calcifiate, iar predentina este marit. Datorit slabei calcifieri a dinilor hipoplazici, crete frecvena cariilor. La adult carena de vitamina D determin osteomalacia, rezorbia arcadelor alveolare, iar n caz de fracturi se ntarzie consolidarea calusului osos. 3.7.3. Vitamina C Principalele funcii ale vitaminei C sunt: formarea fibrelor de colagen din structura tesutului conjunctiv, menine integritatea morfofuncional a endoteliului vascular, menine funcia specializat a odontoblastelor i osteoblastelor. Deficitul de vitamina C la animale determin atrofia odontoblastelor, scderea ratei de formare a dentinei, degenerescena celulelor din pulp, hemoragii (datorit creterii permeabilitii capilare), degenerescena ligamentului periodontal ( se nsoete de creterea mobilitii dinilor n alveol), degenerescena osteoblastelor care se transform n fibroblati, atrofia osului alveolar. Ameloblastele i smalul dentar sunt normale. Deficitul de vitamina C la om determin: degenerarea odontoblastelor care produc substana de tip osos ce umple canalele pulpare, hipermia pulpei, edemul pulpei care apoi se atrofiaz, calcifierea difuz a esutului pulpar, dentina este poroasa, apare osteoliza osului alveolar. De asemenea, se observ fenomene de gingivit caracterizate prin congestia gingiei, edemul gingiei, hemoragii gingivale. Mucoasa bucal devine susceptibil la infecii i la iritanii mecanici. "periajul dinilor produce hemoragii ale mucoasei gingivale. Carena cronica de vitamin C determin boala numit scorbut.
45 Vitamina B1 sau vitamina antinevritic asigur troficitatea nervilor. Deficitul de vitamin B1 determin neurite ale nervilor pulpari, o sensibilitate marcat a mucoasei bucale, edem gingival, dureri nevralgice n regiunea maxilar i occipital. Carena de B1 este un factor cauzator al parodontopatiilor. Vitamina B2 sau riboflavina particip la meninerea troficitii normale a sistemului dento-maxilar. Deficitul de vitamin B2 determin leziuni ale buzelor (cheilit), stomatit comisural, ulceraii n cavitatea bucal, glosit (inflamaia limbii), hipersalivaia reflex. Vitamina PP sau acidul nicotinic n caz de carena determin inflamaii gingivo-linguale, salivaie excesiv, dureri bucale. Acidul pantotenic - carena sever determin gingivo-stomatita necrotic, osteoporoza osului alveolar, atrofia crestei alveolare. Deficitul de vitamin B6 i B12 se nsoete cu cheilit, leziuni dureroase ale limbii, mucoasei bucale. 3.7.8. Vitamina E - carena ei n cursul dezvoltrii dinilor duce la: degenerescena organului adamantin cu hipoplazia smalului, leziuni ale vaselor mici cu repercursiuni negative la nivelul parodoniului.
3.8. EFECTELE HORMONILOR ASUPRA STRUCTURILOR BUCO-DENTARE
Hormonul somatotrop (STH): stimuleaz sinteza colagenului din dentin i cement, crete numrul osteoblastelor i activeaz funcia acestora, mbuntete vascularizaia pulpar, crete depunerea de dentin, cement i rata erupiei. Secreia acestui hormon n exces conduce la macroglosie i la hipertrofia mandibular n care se formeaz aa-numita "mandibul n galo". Hormonii tiroidieni: influeneaz pozitiv creterea odontoblastelor i formarea dentinei, activeaz vascularizaia pulpei i rata erupiei, stimuleaz creterea dinilor. Excesul producerii acestor hormoni determin creterea eliminrii pe cale urinar a calciului i fosforului, precum i a eliminrii fecale de calciu prin accentuarea peristaltismului. De asemenea, excesul de hormoni tiroidieni, prin stimularea activitii odontoblastelor, condiioneaz
46 formarea dentinei neregulat mineralizat sau poate avea efecte toxice pe odontoblaste. n cazul deficitului de hormoni tiroidieni se constat diminuarea ratei de cretere a dentinei, mineralizarea deficitar a dentinei i smalului, slbirea vascularizaiei pulpei, scderea dimensiunii coroanei, a ratei erupiei i a dimensiunii rdcinii. Se mai observ slaba dezvoltare a mandibulei care duce la ocluzie deficitar i creterea incidenei cariilor. Insulina. n cazul prezenei diabetului zaharat se pot remarca o serie de modificri, ca: gingii roii, papile interdentare hemoragice, degenerescena gras a mucoasei gingivale, scderea rezistenei la infecii. Apare gingivostomatita diabetic, parodontopatia diabetic, crete formarea tartrului dentar, se produc microabcese n gingii. Existena pH-ului acid care favorizeaz i intensific activitatea osteoclastic determin rezorbia arcadei alveolare, parodontopatie i creterea incidenei cariilor. Parathormonul (PTH): n exces, realizeaz mobilizarea Ca+2 din oase care duce la decalcifierea maxilarului, acesta devenind maleabil. Prezena crescut de citrat i de fagocite n pulp provoac decalcifierea dentar variabil. Ca urmare se constat modificarea poziiei dinilor, malocluzia i rezorbia osoas. Apariia deficitului de PTH naintea vrstei de 3-4 ani determin tulburri n calcifierea smalului, a dentinei; smalul apare opac, erodat, cu striuri transversale. Se produc hipoplazii dentare, cu rdcini scurte, neuniforme, incomplet dezvoltate i cu anomalii de poziie. Crete, de asemenea, rata formrii cariilor. 3.8.5 Hormonii sexuali feminini produc intensificarea vascularizaiei i creterea permeabilitii pulpare, dezvoltarea ligamentului periodontal, stimularea formrii osului, a exudatului gingival la ovulaie i sarcin, condiioneaz apariia gingivitei de sarcin sensibiliznd gingia la efectul iritanilor. Crete mobilitatea dinilor datorit aciunii hormonului relaxina i datorit scderii incorporrii de prolina n ligamentul periodontal. De asemenea, sarcina determin creterea frecvenei cariilor.
47
4. FIZIOLOGIA METABOLISMULUI FOSFO-CALCIC
4.1. REPARTIIA CALCIULUI N ORGANISM
Calciul este elementul mineral cel mai abundent din organismul uman. Un adult are aproximativ 1000-1500 g de calciu (27,5 moli) ceea ce reprezint 1,5% din greutatea corporal. Repartitia calciului este inegal, astfel: 4.1.1. Calciul depozitat. 99% din calciu este depozitat n oase, dinti. Calciul din depozitele osoase este de dou tipuri: - componenta fixat 1200 g (25000 moli) reprezentat de calciu din cristalele de hidroxiapatit, care poate fi mobilizat prin procese de remodelare osoas, controlat hormonal. Zilnic, prin resorbie si depunere osoas se schimb 7,5 moli; - componenta de schimb (100 moli) reprezentat de calciu de la suprafaa cristalelor de hidroxiapatit care particip la schimburile dintre os si lichidul extracelular. 4.1.2. Calciu intracelular (10 g) se gasete in reticolul endoplasmatic si mitocondrii, care l stocheaza si l stocheaz si l elibereaz cand scade calciul liber citosolic. Sub influena hormonilor, neuro-transmitorilor are loc creterea calciului citosolic fie prin eliberarea lui din depozitele intracelulare, fie prin intrarea calciului din lichidul extracelular. Creterea calciului citosolic determin rspunsuri specifice: contracia muscular, secreia glandular, rspunsuri corelate cu existena n celule a unor proteine fixatoare de calciu: troponina, calmodulina, calbindina. Calciul citosolic este mesager secund intracelular de importan vital. 4.1.3. Calciul plasmatic reprezint 4,5 - 5,5 mEq/1 sau 9 11 mg/dl sau 2,25 2,75 moli/l repartizat astfel: - calciul nedifuzibil (1,16 moli/l) reprezint 40-45% din calciul plasmatic, este legat de albumine si globuline reprezentand rezerva circulanta de calciu. Fixarea calciului pe albumine depinde de pH: alcaloza favoriznd aceasta fixare; - calciul difuzibil (0,16 moli/l) reprezint 5-10% din calciul plasmatic i cuprinde calciul fixat pe anionii organici: fosfat, citrat, bicarbonat; - calciul ionizat (1,18 moli/l) reprezint 50% din calciul plasmatic. Rolurile calciului: intervine n excitabilitatea neuro-moscular (scderea calciului determin creterea excitabilitii neuromusculare), asigur transmiterea influxului nervos, particip la contracia, relaxarea muschilor striai i netezi, stabilizeaz membranele celulare, activeaz sistemul coagularii i al kininelor, este simpaticomimetic asupra inimii, stimuleaz eliberarea de gastrina, moduleaz efectele unor hormoni, este mesagerul secund intracelular. Calcemia este riguros controlat printr-un mecanism neuro-
48 umoral complex. Scaderea calcemiei determin tetanie caracterizat prin: parestezii, hiperexcitabilitate neuro-muscular, spasm carpopedal, dublete, triplete pe EMG. Cresterea calcemiei determin tulburari cardiovasculare, calculoza, constipatie.
4.2. REPARTIIA FOSFORULUI N ORGANISM
La un adult cantitatea de fosfor este de aproximativ 600-900 g repartizat astfel: - fosforul depozitat n oase i dini sub form de fosfat tricalcic (85%); - fosforul intracelular ce intr n structura compuilor macroergici (ATP, CP) i a acizilor nucleici (14%) - fosforul plasmatic (1 - 1,45 moli/l) reprezint 0,03% din totalul de fosfor din organism. Fosforul sub form ionizat are numeroase roluri: particip la mineralizarea osului, reprezint acumulator energetic celular (ATP, CP), intrand n structura membranelor celulare, a acizilor nucleici, activeaz vitaminele B1, B6, particip la contracia muscular, intr in structura sistemelor tampon plasmatice si celulare, etc.
4.3. APORTUL DE CALCIU I FOSFOR
Necesarul zilnic de calciu i fosfor la adult este de aproximativ 800 mg. Acest necesar zilnic este crescut in efortul fizic, la batrni (datorit deficitului de digestie si absorbtie secundar modificarilor atrofice ale tractului digestiv), la copii in perioadele de crestere, la gravide si in tirnpul lactatiei (necesarul de calciu si fosfor ajunge la 1200 mg/zi). Aportul de calciu i fosfor este asigurat de urmtoarele alimente: lapte (125 mg Ca+2 la 100 g aliment), cacaval (500 mg Ca+2 la 100 g aliment), nuci (140 mg Ca+2 la 100 g aliment), pete, roii, ceap verde, varz, etc.
4.4. ECHILIBRUL FOSFO-CALCIC
n organism exist un control neuro-umoral complex care se refer la aportul, stocarea i eliminarile de calciu i fosfor (fig. 9). ntre aceste trei aspecte exist un echilibru ce asigur homeostazia fosfo-calcic. Prin diet se aduc zilnic 1000 mg Ca+2 din care n tubul digestiv se absorb aproximativ 350 mg. La nivel intestinal se
49 produce o secreie de 150 mg calciu i astfel prin fecale se elimin zilnic 650-800 mg calciu. Calciul absorbit la nivel intestinal este trecut n plasm i particip la procesele de remodelare osoas, zilnic fixndu-se n os aproximativ 500 mg calciu, aceeai cantitate fiind i mobilizat zilnic din os. Prin urin se elimin zilnic aproximativ 200 mg calciu.
Fig. 9. Echilibrul fosto-calcic (dup PATTON, 1989, modificat)
4.5. ABSORBIA INTESTINAL A CALCIULU1 SI FOSFORULUI
4.5.1. Calciul este absorbit n segmentul proximal al intestinului subire, n duoden, prin mecanism activ cu participarea proteinelor calcipexice (proteine Fixatoare de calciu) i a ATP-azei calciu dependent, prezente n marginea i perie a eritrocitelor. Acestea se sintetizeaz n enterocite n prezena vitaminei D. n restul intestinului i colonului absorbia calciului se realizeaz prin difuziune lirnitat. Transportul poate fi bidirectional: calciul trecnd din lumenul intestinal n mucoas i invers (se poate astfel spolia organismul de calciu). Factorii ce infuenteaz absorbia intestinal a calciului - n perioada de crestere rapid, la gravide, n timpul lactaiei este favorizat absorbia de calciu; - la btrni scade absorbia calciului datorit modificarilor morfofuncionale ale tractului digestiv; prezena glucidelor ca: lactoza, manitol, sorbitol, arabinoza, n intestin favorizeaz absorbia calciului; arginina, lizina, triptofanul, prezent n lumenul intestinal cresc absorbia calciului; sarurile biliare favorizeaz absorbia calciului prin formarea de complexe solubile de calciu i prin creterea permeabilitii
50 straturilor lipidice ale membranei eritrocitului pentru calciu; pH-ul mai acid favorizeaz dizolvarea srurilor de calciu ceea ce face ca absorbia calciului s fie facilitat. Administrarea de lapte acidulat la sugar asigur o buna absorbie a calciului. n caz de rezecie gastric, aclorhidrie, scade absorbia intestinal a calciului; cresterea cantitaii de lipide n intestin determin scaderea absorbiei de calciu deoarece lipidele formeaz sapunuri cu calciu. n caz de steatoree crete eliminarea prin fecale a lipidelor, calciului i fosforului; prezena sodiului n cantitate mare n intestin diminueaz absorbia calciului; acidul fitic din cereale leag calciul formnd sruri insolubile ceea ce duce la scderea absorbiei de calciu; celuloza, oxalaii scad absorbia de calciu; -- forma de ingerare a calciului: srurile solubile (clorul, gluconat, lactat, carbonat) se absorb uor iar srurile insolubile (fosfai, sulfai, oxalai de calciu) se absorb numai dupa transformarea lor n sruri solubile n prezena acidului clorhidric; raportul Ca/P n coninutul intestinal influenteaz absorbia calciului deoarece excesul de fosfat determin precipitarea intraluminal a calciului i scderea absorbiei; factorii horrnonali i vitarninici: PTH, STH vitamina D cresc absorbia calciului. Glucocorticoizii, ACTH, stresul scad absorbia calciului; factorii sezonieri: n lunile iulie, august este favorizat absorbia calciului, iar n lunile februarie, martie este defavorizat absorbia calciului. 4.5.2. Fosforul este absorbit sub forma de fosfat n jejun i colon. Absorbia fosfailor este facilitat de pH-ul uor acid, excesul de lipide din intestin, prezena sodiului n intestin, a lactozei, de PTH i vitamin D. Absorbia fosfatului este diminuat prin prezena srurilor alcaline, prezena cantiatilor crescute de calciu n intestin.
4.6. ELIMINAREA DE CALCIU I FOSFOR
4.6.1. Calciul se elimin prin: urin, fecale, transpiraie. Prin fecale se elimin sub form de fosfat tricalcic aproximativ 2/3 din calciu de aport. Eliminarea calciului n fecale este scazut n perioadele de crestere, la gravide, n prezena vitaminei D. Crete eliminarea calciului n fecale la btrni, n malabsorbie, diaree, steatoree, hipovitaminoza D, hipertiroidism insuficien de PTH. Prin urin se elimin aproximativ 200 mg calciu pe zi. Calciul
51 filtreaz la nivelul glomerulului renal i este resorbit prin mecanism activ n tubul proximal i distal. Excreia urinar a calciului depide de: nivelul calcemiei: creterea calcemiei determin creterea eliminarilor renale de calciu (hipervitaminoza D, exces de PTH, metastaze osoase, boli osteolitice); aportul alimentar crescut de proteine, glucoz, lactoz crete excreia urinar a calciului; creterea lichidului extracelular determin diureza crescut cu calciurie agenii natriuretici, acidoza metabolic determin creterea calciuriei. 4.6.2. FOSFORUL se elimin prin fecale (300 mg/zi) i prin urin sub forma de fosfai (600 mg/zi). Crete eliminarea de fosfat n urin n urmatoarele situaii: perfuzii rapide, vasodilataie renal, administrarea de diuretice (Furosemid, Acetazilamina), exces de PTH, deficit de vitamina D.
4.7. REGLAREA METABOLISMULUI FOSFO-CALCIC
Metabolismul fosfo-calcic este controlat de factorii vitaminici i hormonali care regleaz absorbia, stocarea i eliminarea calciului i fosforului meninnd astfel calcemia i fosfatemia normal. 4.7.1. Receptorul calcic este un receptor proteic cu structur i funcie asemanatoare altor receptori cuplai cu proteina G, cu deosebirea c ligandul nu este o molecul organic ci un ion anorganic. La om receptorul calcic conine 1078 aminoacizi, avnd 93% identitate de secven cu receptorul calcic bovin. Structura receptorului calcic include un domeniu extracelular mare, 7 domenii transmembranare i un domeniu intracitoplasmatic. Se remarc n poriunea extracelular expresia a patru regiuni cu reziduuri aminoacidice caracteristice proteinelor de mic afinitate fixatoare de calciu ca i calsequestrina (fig. 10). Aceste regiuni pot fi implicate n legarea calciului extracelular, detectnd modificrile fiziologice ale calciului i astfel regleaz funciile osteoclastelor, celulelor paratiroidiene, parafoliculare C.
52
Fig. 10. Schema receptorului calcic (dup NEMETH, 1995, modificat)
Gi = Proteina G inhibitoare; AC = Adenilat ciclaza; ATP = Acid adenozintrifosforic; AMc = Acid adenozinmonofosforic ciclic; PLC = Fosfolipaza C; PIP2 = Fosfatidilinozitol 4,5 bifosfat: DG = Diacilglicerol; IP3 =inozito1,4,5 trifosfat: PKC = Proteinkinaza C.
Celulele paratiroidiene, parafoliculare, nefrocitele, celulele G secretoare de gastrin conin de asemenea receptor calcic pe membrana celular . Creterea local a calciului extracelular la 5-15 mM, determnd fixarea lui pe receptorul calcic al osteoclastelor, receptor cuplat cu proteina Gq ce activeaz fosfolipaza C (PLC). PLC desface fosfatidilinozitol difosfatul (PIP2) n inozitol trifosfat (IP3) i diacilglicerol. Rezultatul creterii nivelului IP3 este mobilizarea calciului din stocul nonmitocondrial, probabil din reticulul endoplasmic. De asemenea se semnaleaz un influx de calciu prin canale operate de receptor. Deci, creterea calciului extracelular, prin intermediul receptorului calcic de pe osteoclaste, induce creterea calciului intracelular ceea ce determin, printr-un mecanism de feed-back negativ, inhibiia resorbiei osului dependent de osteoclaste. Creterea calciului intracelular dezorganizeaz podosomii inducnd detaarea osteoclastului de matricea osoas i oprirea resorbiei osului pan cnd concentraia calciului extracelular este identic cu cea prezent n microcompartimentul acid situat ntre marginea n perie a osteoclastului i os. Aceast reglare negativ a activitatii osteoclastice prin calciul extracelular este un mecanism autolimitant al resorbiei osoase. Creterea calciului extracelular urmat de fixarea lui pe receptorul calcic al celulelor paratiroide determin inhibiia adenilat-ciclazei prin proteina Gi i activarea PLC prin proteina Gq. Aceste fenomene induc creterea calciului intracelular prin eliberarea calciului din depozitele celulare i prin influx de calciu operat pe receptor. Aceast cretere a calciului intracelular produce inhibiia secreiei de parathormon (PTH). Secreia de PTH este reglat i de PKC prin:
53 feed-back negativ arnortizeaz semnalizarea pe calea receptorcalcic protein Gq PLC; fosforilarea direct care PKC a receptorului calcic ceea ce induce scderea sensibilitaii receptorului la activare prin calciu extracelular ca scderea capacitaii receptorului calcic de a se cupla cu proteina Gq. Cresterea calciului extracelular determin activarea receptorului calcic al celulelor parafoliculare C urmat de cresterea secretiei de calcitriol Stimularea receptorului calcic de pe nefrocite este implicat n cresterea sintezei de calcitriol. Semnificaia terapeutic a existenei receptorului calcic este multipl. Compui numii "calcimimetici" care mimeaz sau poteneaz aciunile calciului extracelular ca agoniti ai receptorului, acioneaz selectiv deprimnd funciile celulelor paratiroide sau a osteoclastelor. Preparatele "calcimimetice" ce se adreseaz receptorului de pe osteoclaste ar putea reprezenta o noua terapie antire-sorbtiva pentru tratamentul osteoporozei. Compusul, NPS R568 este un calcimimetic paratiroidian folosit curent n experimentele clinice ca tratarnent pentru hiperparatiroidism.
4.7.2. Vitamina D Vitaminele D cuprind mai muli compui sterolici, dar din punct de vedere fiziologic numai doi prezint importana pentru om : vitamina D2 (ergocalciferol) din regnul vegetal i vitamina D3 (colecalciferol) de origine animal exogen dar i de origine endogen. Sursele exogene de vitamina D sunt: untura de pete, sardelele, untul. ficatul, frica, lapte, ou, etc. Sursa endogen de vitamina D provine din 7-dehidrocolesterol, care n piele sub aciunea razelor ultraviolete se transform n vitamina D3 (colecalciferol). Aceasta este transportat la ficat legat de o globulin specific fixatoare de vitamina D (DBP). La ficat este activat prin hidroxilare rezultnd 25hidroxicolecalciferol, iar la rinichi sub aciunea 1-alfa hidroxilazei se formeaz 1,25 dihidroxicolecalciferol (calcitriol) (fig. 11). Formarea 25-hidroxicolecalciferolului n ficat, prin mecanism feedback, inhib hidroxilaza hepatic ceea ce are drept consecine: prevenirea efectelor excesului de vitamin D care s-ar forma n cantitate prea mare i limitarea conversiei vitaminei D3 n metabolitul activ, conservnd aceast vitamin pentru consumul ulterior. Vitamina D3 poate fi stocat n ficat luni de zile pe cnd 25hidroxicolecalciferolul se pstreaz numai cteva sptrnni. Calcitriolul, cel mai activ metabolit al vitaminei D3 se mai poate forma n placenta, keratinocite i macrofage.
54
ACTIVAREA I ROLUL VITAMINEI D Fig. 11. Activarea i rolul vitaminei D (dup PATTON, 1989, modificat) 4.7.2.1. Efectele calcitriolului Calcitriolul determin creterea calcemiei prin aciune la trei nivele: la nivelul epiteliului intestinal activeaz proteinele calcipexice, fosfataza ATP-aza calciu dependent, determinnd astfel creterea absorbiei intestinale a calciului. Proteinele calcipexice se gsesec pe marginea n perie a epiteliului intestinal determinnd transportul calciului din lumen n enterocit iar apoi prin membrana bazo-lateral calciu trece n snge printr-un mecanism de difuziune facilitat. Rata absorbiei calciului este proportional cu cantitatea de proteine calcipexice. Dintre proteinele calcipexice intestinale amintim: calbindina D9k care leag 2 ioni de calciu i calbindina D28k cu greutate molecular de 28000 i care leag 4 ioni de calciu. Efectele proteinelor calcipexice dureaz cteva saptmni dup ce s-a sintetizat calcitriolul; la nivel renal determin reabsorbia calciului i fosforului n tubul proximal; la nivel osos stimuleaz creterea numrului i funciei osteoclastelor, mobilizeaz calciu i fosforul din os, favorizeaz transportul activ al calciului din osteocite n lichidul extracelular.
55 4.7.2.2. Mecanismul intracelular de aciune al calcitriolului Calcitriolul se leag de receptori specifici intracitoplasmatici formnd complexul calcitriol-receptor. Acest complex ptrunde n nucleu, activeaz ADN-ul celular inducnd formarea unui ARNm specific, care migreaz n citoplasm, declannd sinteza de proteine specifice calcipexice i pentru pompa de calciu. 4.7.2.3. Reglarea sintezei de calcitriol Scderea calcemiei induce stimularea secreiei de parathormon ce activeaz 1-alfa hidroxilaz renal i deci sinteza de calcitriol. Scderea fosfatemiei stimuleaz activitatea renal a 1-alfa hidroxilazei. De asemenea estrogenii, prolactina activeaz enzima 1alfa hidroxilaz i sinteza renal de calcitriol. Sarcina, lactaia, hormonul somatotrop, hormonii corticosuprarenali, calcitonina, hipertiroidismul activeaz sinteza de calcitriol. Creterea fosfatului sanguin, acidoza, hipoinsulinemia inhib sinteza de calcitriol. Carenta de vitamin D determin la copil, rahitismul. 4.7.3. Parathormonul (PTH) PTH este un polipeptid ce conine 84 aminoacizi, are greutate molecular 9500, este produs de glandele paratiroide. Sinteza de PTH se realizeaz n ribozomi sub forma de prehormon (110 aminoacizi) care prin clivare proteolitic devine prohormon cu 90 de aminoacizi. Aceasta se transform n PTH n reticolul endoplasmatic i aparatul Golgi, fiind apoi nglobat n granulele secretorii ale celulelor paratiroide. Secreia PTH n celulele paratiroide este AMP ciclic dependent. 4.7.3.1. Efectele PTH Parathormonul are efect hipercalcemiant prin aciune la nivel renal, intestinal i osos. La nivel renal efectele PTH sunt rapide i constau din: stimularea 1-alfa hidroxilazei deci sinteza de calcitriol; creterea reabsorbiei calciului n ramura ascendent a ansei Henle, tub distal, tub colector; crete absorbia tubular a magneziului; inhibarea reabsorbtiei fosfatului n tubul proximal determinnd fosfaturie; inhibarea activitaii anhidrazei carbonice din nefrocit rezultnd eliminarea crescut de bicarbonat de sodiu n urin, cu alcalinizarea urinii; scderea eliminrii de H+ i NH4+, crend astfel mediu optim
56 pentru meninerea calciului n form ionic. La nivel intestinal efectele PTH se realizeaz prin intermediul calcitriolului, crescnd absorbia intestinal de calciu i fosfat. La nivel osos efectele PTH se realizeaz n dou faze. n prima faz PTH mobilizeaz calciu i fosfatul din os prin efecte rapide (activeaz pompa de calciu din osteocite i astfel calciul trece din osteocit n lichidul extracelular) i tardive (activeaz sinteza de not enzime lizozomale, colagenaze i formare de acizi n osteoclaste determinnd resorbia osului). n faza a doua inhibnd osteoblastele, sinteza de colagen, scade activitatea fosfatazei alcaline i diminund osteogeneza. PTH induce sinteza de IL-1 n osteoblaste. Aciunile PTH-ului se realizeaz prin fixarea sa pe receptorii specifici cuplai cu proteine G, i cu adenilat ciclaza, creterea AMPc n nefrocite, osteoclaste. AMPc induce: sinteza de enzime lizozomale n osteoclaste, sinteza 1-alfa hidroxilazei n nefrocite. 4.7.3.2. Reglarea secretiei de PTH Reglarea secreiei de PTH se face prin mecanismul de feed-back dependent de nivelul calcemiei. Scderea calcemiei stimuleaz secreia de PTH, iar creterea calcemiei inhib secreia de PTH. Rahitismul, sarcina, lactaia stimuleaz secreia de PTH. Catecolaminele, dopamina, histamina, vitamina A, cortizonul activeaz sinteza n celulele paratiroide de PTH. Ablatia chirurgical a paratiroidelor determin hipocalcemie, tetanie, spasm laringo-traheal, moarte. Deficitul de PTH (hipoparatiroidism) determin hipocalcemie cu parestezii, palpitaii, tulburari neuropsihice, hiperexcitabilitate neuromuscular manifestat clinic prin crize tetanice iar pe EMG prin dublete, triplete, etc. Excesul de PTH (hiperparatiroidism) determin: demineralizarea osoas, hipercalcemie, calciurie, calculoz renal, etc. Efectele excesului de PTH asupra structurilor buco-dentare sunt: decalcifierea maxilarului, care devine maleabil, se deformeaz, malpoziii dentare, malocluzie, resorbia osului alveolar. Insuficiena de PTH sub 3-4 ani determin hipoplazii dentare, tulburarea calcifierii smalului i a dentinei, radacin scurt, neuniform, incomplet dezvoltate, anomalii de poziie a dinilor, carii. 4.7.4. Calcitonina Calcitonina este un polipeptid cu greutate molecuiar de 3400, conine 32 aminoacizi, este sintetizat de celulele parafoliculare din tiroid, timus, paratiroide. n celulele parafoliculare se sintetizeaz ca preprocalcitonin din care apoi se cliveaz calcitonina i katacalcin (poteneaz aciunile calcitoninei). Timpul de njumataire al calcitoninei este de mai putin de 10 minute, de aceea
57 ea intervine n reglarea de scurta durata a calcemiei. 4.7.4.1. Efectele calcitoninei Efectul calcitoninei este hipocalcemiant. Receptori pentru calcitonin s-au evideniat n os, pe osteoclaste i n rinichi. Efectele intracelulare ale calcitoninei sunt mediate de AMPc. Calcitonina inhib activitatea osteoclastelor, inhib degradarea colagenului din os, suprim eliberarea enzimelor proteolitice, inhib mobilizarea calciului din os, deci, determin reducerea resorbiei osoase. De asemenea, stimuleaz formarea osului, scade permeabilitatea membranei osteocitare pentru calciu, mpiedicnd ieirea calciului din os. Calcitonina crete excreia urinar de sodiu, calciu, fosfat, magneziu. 4.7.4.2. Reglarea secreiei de calcitonina Reglarea secreiei de calcitonina este dependent de nivelul plasmatic al calciului: creterea calcemiei stimuleaz secreia de calcitonina care induce scderea calcemiei. Ali factori stimulatori ai calcitoninei sunt: dopamina, agoniti beta-adrenergici, estrogeni, gastrina, secretina, colecistokinina, glucagon, calcitriol, etc. 4.7.5. Hormonii glucocorticoizi Glucocorticoizii intervin n rnetabolismul fosfo-calcic prin efecte antivitamina D, stimuleaz eliberarea de PTH i inhiba secreia de calcitonina. Glucocorticoizii n exces produc osteoporoza prin scderea osteogenezei, creterea reabsorbiei osoase, scderea reabsorbiei intestinale de calciu i fosfat prin efect antivitamina D, crete eliminarea renal de calciu i fosfat. Scderea osteogenezei de ctre glucocorticoizi se realizeaz prin: mobilizarea aminoacizilor din matricea organic a osului, inhibarea proteino-sintezei, inhibarea diviziunilor celulare, reducerea funciilor osteoblastelor i a sintezei de colagen. Excesul de glucocorticoizi determin scaderea calcemiei ceea ce duce la stimularea secreiei de parathormon care faciliteaz resorbia osoas prin activarea osteoclastelor. 4.7.6. Hormonii tiroidieni Tiroxina are efect condrogenic, mrete viteza de rennoire a structurilor osoase prin stimularea direct a osteoclastelor i resorbia osoas. 4.7.7. Hormonul somatotrop (STH) STH crete eliminarea renal a calciului dar stimuleaz i
58 absorbia intestinal a calciului rezultnd din bilan calcic pozitiv. STH stimuleaz proteinosinteza osoas prin somatomedine, stirnuleaz ncorporarea sulfatului n proteinele cartilajului n cretere. 4.7.8. Hormonii sexuali Estrogenii i androgenii au efect anabolizant asupra osului, accelereaz creterea i maturarea scheletului, cresc absorbia intestinal a calciului, stimuleaz mineralizarea osului, scad eliminarea de calciu n urin. Hormonii sexuali poteneaz aciunea calcitoninei i calcitriolului. Deficitul hormonal la menopauz se insoete de osteoporoza. 4.7.9. lnsulina Are efecte anabolizante, trofice asupra osului, stimuleaza osteogeneza, diabetul zaharat netratat se menioneaza scaderea masei osoase.
4.8. FIZIOLOGIA OSULUI
Osul este o varietate de esut conjunctiv format dintr-o matrice de fibre de colagen impregnata cu sruri de calciu. Sistemul osos protejeaza organele vitale, prin rigiditatea sa asigur locomoia i suportul forei antigravitaional. Osul este bine vascularizat: fluxul total sanguin osos fiind de 200-400 ml/minut la adulti. In permanen are loc formarea i resorbia osului realizndu-se astfel participarea osului la homeostazia fosfo-calcic. Sistemul necelular al osului cuprinde componente organic (30%) i componenta mineral (70%). 4.8.I.Componenta organic major a matricei extracelulare este format la principal din fibre de colagen de tip I, sub forma de triplu helix ce cuprinde 2 lanuri polipeptidice alfa- 1 i 1 lan polipeptidic alfa-2. In structura primar a colagenului se gsesc in cantitate mare: glicin, lizin, protin. Fibrele de colagen sunt orientate perpendicular pe liniile de fort induse de solicitarile mecanice. Componentele necolagenice din matricea organic a osului sunt in proporie mic reprezentate de proteine, glicoproteine, mucopolizaharide. Dintre proteine mai bune studiate sunt: osteocalcina, este sintetizat de osteoblaste dependent de vitamina K, are o greutate molecular de 6000, conine reziduu de acid gamacarboxiglutamie ce fixeaz ionii de calciu. Sinteza osteocalcinei este reglata de calcitriol; osteonectina, are greutate molecular 32000, prezinta situsuri de legare la fibrele de colagen si cristalele de hidroxiapatita. Este implicat in fixarea calciului pe colagen;
59 osteopontina, fibronectina favorizeaza ataarea celulelor osoase la matricea extracelul. 4.8.2. Componenta mineral cuprinde fosfat tricalcic organizat in cristale de hidroxiapatita: Cato(PO4)6(OH)2, de form hexagon lamel cu dimensiuni de 20/3-4 nanometri. De asemenea in cristalele de hidroxiapatita se mai gasesc ioni de sodiu, potasiu, magneziu, carbonat, fluor. La suprafaa cristalelor se gasete atmosfer hidratat, ionizat ce reprezint componenta mineral de schimb a osului. Sistemul celular al osului este format din: osteocite, osteoblaste, osteoclaste. 4.8.3. Osteocitele sunt celule adulte, de fapt, osteoblaste prinse in matricea pe care ele insele au format-o. Osteocitele au prelungiri prin care se conecteaz cu alte osteocite sau cu suprafaa osului. Conin numeroase mitocondrii i vacuole citoplasmatice, sunt implicate n remodelarea osului. mpreuna cu osteoblastele formeaza membrana osteocitar ce separ lichidul din jurul matricei mineralizate de sectorul hidric interstiial al osului. Asigura schimburile osului cu lichidul extracelular. Calcitriolul i PTH favorizeaza transportul activ al calciului din osteocit n lichidul extracelular iar calcitonina scade acest transport (fig. 12). 4.8.4. Osteoblastele sunt celule formatoare ale osului, elaboreaza componentele matricei organice i o pregatesc pentru mineralizare. Osteoblastele provin din celulele mezenchimale medulare, maturarea lor necesitnd aproximativ 5 zile i este controlat de factori de cretere locali i hormoni. Osteoblastele sunt celule cu diametru de 20 micrometri, au un bogat reticul endoplasmatic rugos i aparat Golgi bine dezvoltat, ambele participnd la sinteze proteice. Osteoblastele sintetizeaz colagen tip I, fosfaz alcalina,fibronectin, osteocalcin, sialoproteine, factori de cretere ce regleaz local osteoformarea. Fosfataza alcalina este implicate n mineralizarea osului prin hidroliza pirofosfatului (inhibitor al mineralizrii) i prin creterea local a fosfatului. Deficitul ereditar de fosfataz alcalin duce la osteoporoza, fracturi, pierderea prernatura a dentiiei definitive, nefrolitiaz. Osteocalcina are efect chemotactic pentru precurserii osteoclastelor.
60
Fig. 12. Relaiile funcionale ale celulelor osoase (dup PATTON, 1989, modificat) PTH = Parathormon; CT = Calcitonin; DHC = 1,25 Dihidroxicolecalciferol; LEC = Lichid extracelular Factorii de cretere locali sintetizai de osteoblaste sunt: insulin like growth factors (IGF I, IGF II), factorii de cretere a fibroblatilor (fibroblasts growth factors FGF). FGF basic se leag de proteoglican, colagen, fibronectin crescndule stabilitatea i le protejeaz de proteoliz. FGF basic produce asupra celulelor osteoblastice stimularea multiplicrii celulare, inhib sinteza de colagen i activitatea fosfatazei alcaline. IGF se gsesc n cantitate mare n matricea osoas, sunt factorii anabolici importani pentru esutul osos pentru c stimuleaz proliferarea i diferenierea osteoblastelor. TGF- este mitogen i chemotactic pentru osteoblaste, stimuleaz sinteza de integrine i aderena celular la matrice, recruteaz noi celule osteoblastice la locul formrii osului, stimuleaz sinteza de colagen, inhib formarea de colagenaz i a activatorului plasminogenului. Osteoblastele au receptori numeroi pentru PTH, vitamina D3, dar pentru estrogeni (stimuleaz sinteza de colagen), progesteron (stimuleaz proliferarea osteoblastelor). Vitamina D3 fixandu-se pe receptorii specifici de pe osteoblaste determin transcripia genelor responsabile de sinteza de: colagen tip I, fosfataz alcalin
61 osteocalcin, osteopontin, fibronectin, de asemenea modific metabolismul fosfolipidelor membranare a osteoblastelor. PTH stimuleaz n osteoblaste producerea de TGF-(3 IGF, IL-1 (slimuleaz proliferarea osteoblastelor, dar inhib formarea osului prin creterea producerii de PGE-2 i prin activarea osteoclastelor), IL-6 (stimuleaz diferenierea osteoclastelor), GM-CSF (granulocite macrophage-colony stimulating factor), este un factor mitogen pentru osteoblaste. M-CSF (macrophage colony stimulating factor) este implicat n diferenierea osteoclastelor. 4.8.5. Osteoclastele sunt celule gigante, multinucleate (100 nuclei), derive din celulele stem hematopoetice sau din celulele seriei monocito-macrofagice. Au numeroase mitocondrii i vezicule cu enzime: fosfataza acid, anhidraza carbonic, hidrolaze, proteaze, catepsin. Osteoclastele sunt celule implicate n resorbia osului: se ataeaz de matricea osoas prin podosomi creind un microcompartiment ntre marginea n perie i os. n acest microcompartiment pH-ul este n jur de 5 asigurat de pompa de protoni (ATP-aza H+ dependent) a osteoclastelor. Protonii excretati sunt Produi n ajutorul anhidrazei carbonice (H2CO3 HCO3-+H+). Aciditatea local determin disoluia fazei minerale a matricei osoase. Degradarea matricei organice se realizeaz de ctre enzimele lizozomale eliberate extracelular: catepsinele i hidrolazele. Osteoclastele au receptori pentru calcitonin. Activitatea de resorbie a osteoclastelor este controlat printr-un mecanism autolimitant de ctre calciul extracelular, existnd un "receptor pentru calciu" pe membrana osteoclastelor. Creterea calciului intracelular dezorganizeaz podosomii (cu ajutorul crora osteoclastele ader la os) ceea ce antreneaz detaarea osteoclastelor de matricea osoas i oprirea resorbiei pan cnd concentraia calciului extracelular este identic cu cea din microcompartimentul acid (situat ntre marginea n perie a osteoclastelor i cristalele de hidroxiapatit). Deci aceast reglare prin feed-back negativ a activitii osteoclastelor de ctre calciul extracelular este un mecanism autolimitant al resorbiei osoase. Activitatea osteoclastelor este stimulat de PTH, vitamina D3, cortizol, hormoni tiroidieni, IL-6, IL-1, TNF i inhibat de calcitonin, estrogen, PGE-2. 4.8.6. Formarea i resorbia osului Formarea osului necesit n primul rnd sinteza de ctre osteoblaste a matricei organice de colagen urmat de mineralizare. Mineralizarea sau calcifierea osului reprezint depunerea de sruri minerale pe matricea organic i se realizeaz n mai multe etape: Nuclearea - reprezint formarea de nuclee cristaline n matricea organic a osului, nuclee cu proprieti calcifine. Nuclearea este
62 activat de colagen i inhibat de pirofosfat, mucopolizaharide. Fosfataza alcalin degradeaz mucopolizaharide favoriznd nuclearea, scindeaz pirofosfatul, crete concentraia local a fosfatului stimulnd mineralizarea. Formarea cristalelor de hidroxiapatit - osteoblastele preiau activ ionii de calciu din fosfat, i concentreaz sub form de fosfat tricalcic amorf n vezicule i l secret la nivelul zonelor calcifine. Aici are loc precipitarea i cristalizarea sub forma de hidroxiapatit. Formarea osului este stimulat de: STH, insulin, androgeni, calcitonin, vitamina D, stimuli mecanici, fracturi i este inhibat de PTH, glucocorticoizi. Remodelarea osoas este un proces fiziologic ce asigur rennoirea matricei osoase ceea ce necesit efortul comun al osteoblastelor i osteoclastelor. n prima faz are loc o activare a multiplicrii i funciilor osteoclastelor care resorb osul vechi formnd o lacuna. Aceasta este rapid populat cu celule de tip macrofagic sau/i precursori ai osteoblastelor. Este faza care precede reconstructia osului. Maturarea osteoblastelor duce la sinteza unei noi matrice osoase. Osteoblastele care sunt prinse n matricea sintetizat de ele devin osteocite care prin canaliculi au conexiuni cu celulele de la suprafaa matricei. Acest ciclu este de aproximativ 100 zile. La procesul de remodelare osoasa n afara osteoblastelor i osteoclastelor mai particip celulele stem multipotente din maduva hematogen, hormonii i factorii locali. Forele mecanice controleaz, de asemenea, remodelarea osoas. Activitatea osteoclastelor este stimulat de PTH, vitamina D, PGE2, IL-I, IL-6, GM-CSF, M-CSF i este inhibat de calcitonin. Osteoblastele sunt activate de estrogeni, progesteron, PTH (activeaz sinteza de colagenaz n osteoblaste), IGF, GM-CSF, TGFb. Principalele citokine au rol n remodelarea osoas intervin n: - diferenierea precursorilor osteoclastelor (IL-1, TNF-a, IL-6, GMCSF, M-CSF, IGF), - diferentierea celulelor osteoblastice (IL-1, TNF-a, GM-CSF, TGF, IGF), - sinteza matricei de cdtrc osteoblaste (TOE], IGF, K') (tabeltd 8). Principalii factori implicai n remodelarea osoas (dup de VERNEJOUL, 1993, modificat) Diferenierea Prolif Sinteza precursorilor erarea matricei de ctre PGE + -/+ 0 IL-1 + + TNF + + IL-6 + 0 0 GM+ + 0 M+ 0 0 TGF + + IGF + +
63 + + Legenda: + : stimuleaz; : inhib; 0 far efect; /+ : efect bifazic. Unde: PGE2 = prostaglandina E2, IL = interleukin TNF = factorul de necroz tumoral, GMCSF = factorul de stimulare al coloniilor granulo-monocitare, MCSF = factorul de stimulare al coloniilor monocitare, TGF = factorul de transformare a creterii, IGF = factorul de cretere insulin-like. FGF = factorul de cretere al fibroblatilor. 4.8.7. Tulburrile metabolismului osos Rahitismul (la copii) si osteomalacia (la adult) se caracterizeaz prin mineralizarea insuficient a osului datorit carenei de vitamina D asociat deficit de calciu. Osteoporoza se caracterizeaz prin reducerea masei osoase datorit unui exces de osteoliz sau diminurii osteogenezei. Se ntalnete n: hipertiroidism, hipogonadism, menopauz (deficit estrogenic), hiperparatiroidism, exces glucocorticoizi, involuie senil, scorbut, malabsorbie, imobilizare prelungit, alcoolism, diabet zaharat, etc. Osteoscleroza caracterizat prin exces de esut osos calcifiat, se intlnete n hipoparatiroidism, intoxicaie cu plumb, cu fluor, cu vitamina D. vitamina A. 4.8.8. Mecanismele cresterii si dezvoltarii osoase la nivelul aparatului dento-maxilar Organogeneza i morfogeneza aparatului dento-maxilar se realizeaz prin dou procese strns corelate: creterea (definit ca "mrirea dimensiunilor") i dezvoltarea ("progres catre maturitate" cu definitivarea structural i functional). Modificrile de dimensiune i form a aparatului dento-maxilar se realizeaz prin mai multe mecanisme: creterea periostal, cresterea la nivelul suturilor, cresterea la nivelul cartilajelor, activitatea osteogenetic i osteoformatoare a ligamentului alveolodentar, modificri de dimensiune, form i raporturi prin aciuni osteotransformatoare i modelante (resorbie si apoziie). 4.8.8.1. Creterea periostal Creterea periostal are un rol deosebit n dezvoltarea oaselor de FGF 0
64 membran. Faa intern a periostului este acoperit cu un rand de osteoblaste, care produc noi straturi de os periferic ce se ncorporeaz intim n osul format anterior. Dup constituirea piesei osoase, activitatea periostal de marire a osului continu s se produc pan la stadiul adult. n timpul vieii intrauterine, toate oasele craniene i faciale cresc activ pe toata suprafata lor. Pe msur ce se apropie de stadiul adult, ritmul de mrire a dimensiunilor devine mai lent, n schimb se precizeaz, din ce n ce mai mult, prin modelaj, forma adult. Modelajul formelor exterioare se realizeaz, n parte, datorit factorilor funcionali i n special aciunii musculaturii. Musculatura, la locurile de inserie are o activitate stimulatoare, realizand zone de osteogenez periostal crescut. Maxilarul superior i mandibula prezint cretere periostal, rata creterii periostale fiind mai mare la mandibul fat de maxilarul superior. 4.8.8.2. Creterea la nivelul suturilor Suturile reprezint modul obinuit de unire dintre oasele extremitii cefalice, ele fiind considerate adevrate periosturi de conjugare. n timpul evolutiei, suturile trec prin mai multe stadii stadiul de sincondroz cele dou oase sunt unite printr-un estut cartilaginos; stadiul de sinfibroz cnd ntre oase exist un esut fibros, ce permite o libertate de deplasare relativ a oaselor; -- stadiul de sinartroz cnd cele dou suprafee osoase n contact, sunt acoperite de periostul de conjugare; studiul de sinostoz cnd ntre cele dou piese s-a creat o punte osoas. Creterea la nivelul suturilor este influenat de fore mecanice. Principalele suturi care asigur dezvoltarea facial i raporturile cu baza craniului sunt orientate n planuri similare ce privesc de sus n jos i dinainte inapoi. Datorit activitii suturilor, faa se mareste n direcie anterioar i inferioar n raport cu neurocraniul. Tipuri de suturi: suturi armonice, n care cele dou oase vin n contact prin suprafete netede (ex. sutura palatin, sutura maxilo-malar); suturi dentate,n care cele dou oase au marginile dantelate ( ex. oasele boltii craniene); suturi scuamoase, cele dou oase se acoper una pe alta (ex. sutura temporo-parietal, palatino-maxilar); sutura de "mbucare", n care una dintre marginile osoase este sub form de creast, iar cealalt sub forma de san (ex. sutura vomero-sfenoidal).
65
4.8.8.3. Creterea la nivelul cartilajelor Cartilajul primar al mandibulei (segmentul anterior al cartilajului Meckel) are rol de susinere, de meninere a formei si asigur o anumit stabilitate a dimensiunilor mandibulei. Un rol important n creterea i dezvoltarea facial revine capsulei nazale i septului nazal cartilaginos. Capsula nazal cartilaginoas formeaz scheletul primordial al prii superioare a feei, iar n jurul ei se dezvolt oasele: maxilare, nazale, lacrimale, etmoidul, vomerul, oasele palatine. Deci, capsula nazal conjunctiv are rol important n determinarea formei iniiale a fetei, iar cartilajul septului nazal intervine n creterea vertical i sagital a prii superioare a feei. Partea postero-superioar a cartilajului condilian, asigur creterea, ndeosebi, vertical a ramurii ascendente a mandibulei. Cartilajul condilian este similar, funcional, cartilajelor de conjugare. 4.8.8.4. Activitatea osteogenetic i osteotransformatoare a ligamentului alveolo-dentar Prezena dinilor determin anumite particulariti ntr-o poriune a oaselor maxilare, alctuind aa numitul os maxilar bucal Watry sau os alveolar. Acest os alveolar se consider ca aparinnd dinilor deoarece se dezvolt odata cu dinii, i regleaz creterea dependent de prezena lor i dispare data cu dispariia dinilor. Fenomenele de cretere ale osului alveolar se realizeaz la nivelul feelor laterale, septurilor interdentare. Erupia succesiv a dintilor produce mriri evidente ale feei n sens vertical. Procesele alveolare i dubleaz dimensiunile ntre 5-12 ani i continu s creasc evident pan la 23 ani. Fortele declanate de aparatele ortodontice produc modificari histologice osoase, n sensul resorbiei i apoziiei. Activitatea ligamentului alveolo-dentar este reglat n primul rnd de influene funcionale. Procesele alveolare, cu toate particularitile lor fac parte din oasele maxilare, se comport n general, ca i sisternul osos. Impulsurile realizate din contactele funcionale interarcadice sunt transmise, prin intermediul unitaii dinte-proces alveolar, restului maxilarelor, iar mai departe oaselor faciale si bazei craniului. Deci totul este un ansamblu unic, arhitectonic i funcional. Schirnbrile din sistemul de transmitere a forelor pot determina modificri n structura i forma oaselor (de exemplu, mrirea sinusurilor frontale n progenii-Prognatisme mandibulare). 4.8.8.5. Modificri de dimensiune, form i raporturi prin actiuni osteotransformatoare i modelante (resorbie i apoziie)
66 Oasele cutiei craniene, pe lang creterea sutural, sufer un proces de apoziie pe faa exterioar, n timp ce resorbia are loc la nivelul tablei interne. La osul frontal aria de resorbie ar fi situat predominant n fundul concavitii, iar la celelalte oase ale bolii craniene ar predomina la periferia osului. Spre aria central, se poate observa apoziie osoas i la nivelul tablei interne. Resorbia osoas are importan i n procesul dezvoltarii la nivelul structurilor faciale: intervine n coborrea planeului cavitailor nazale, intervine n expansiunea cavitailor orbitare, asociat cu creterea sutural, asigur dezvoltarea sinusurilor, sub influena factorilor funcionali. Resorbia i apoziia osoas, n cadrul unui proces corelat, reprezint o modalitate important de cretere mandibular. Astfel, la nivelul marginilor anterioare ale ramurii ascendente, a apofizei coronoide i condilului, se produc resorbii osoase, iar concomitent si paralel are loc un proces de apozitie osoas pe marginea posterioar a ramurii ascendente, a apofizei coronoide, la incizura sigmoid, pe faa posterioar a condilului. 4.8.9. Generaliti privind creterea i dezvoltarea craniofacial intrauterin n saptmana a III-a de viat intrauterin, multiplicarea celular intens face ca discul embrionar s aib o form alungit, dezvoltndu-se tubul neural, mugurii frontali, maxilari i mandibulari care incep s delimiteze cavitatea bucal primitiv (stomodeumul). Invaginarea celulelor ectoderrnice din stomodeum realizeaz lama dentara primar. In saptamana a VI-a de viaa intrauterin, ncepe diferenierea osteoblastelor, cementoblastelor, odontoblastelor ce vor produce esut osos, respectiv cement, respectiv dentina. n aceast perioad se dezvolt mugurii palatinaii, care vor forma bolta palatin secundara. ntre saptamana a VII-a i a XI-a se remarc o multiplicare intens a celulelor din mezoblast, se delimiteaz mugurele frontal se unete cu mugurele maxilar i mandibular formnd faa. Deci, n prima luna intrauterin crete mugurele frontal, n luna a II-a predomin creterea maxilarului superior, iar n luna a III-a predomin creterea mandibulei. La sfritul lunii a III-a distingem maxilarul superior prin apariia celor cinci perechi de centre de osificare desmal. Mandibula se contureaz bine n saptmana a VII-a, cand, lateral de cartilajul Meckel, apare primul centru de osificare desmal urmat de apariia altor centri n ramura ascendent a mandibulei i n poriunea
67 bazilar a viitoarei mandibule. Arcadele alveolare se dezvolt ncepnd din saptamna a VI-a, prin centrii de osificare desmal ce apar n jurul mugurilor dentari. Bolta palatin este realizat prin unirea palatului primar, ce evolueaz din mugurele frontal, cu palatul secundar, derivat din mugurii maxilari. n etapa de morfogeneza, ce urmeaz procesului intens de histodifereniere, se dezvolt scheletul aparatului dento-maxilar i musculatura cervico-facial. Baza craniului: n perioada fetal, baza craniului formeaz masa cartilaginoas (condrocraniul) n jurul cruia se dezvolt elementele scheletice i neuromusculare dupa o anumit schema ereditar. La inceputul perioadei fetale (luna a IV-a) ncepe osificarea bazei craniului prin metaplazie. Maxilarul superior mpreun cu osul zigomatic se formeaz, prin unire, din cele cinci centre de osificare desmal. Mandibula: n luna a IV-a de viat intrauterin n mandibul exist patru centre de osificare desmal, care au tendina sa se uneasc. Centrele de crestere endocondral i desmal din mandibul au o intens activitate influentat si de dezvoltarea limbii (din materialul apartinnd a 5 arcuri branchiale). Bolta palatin se realizeaz prin fuzionarea centrelor de osificare nazofrontal cu cele laterale, care deriv din maxilarul superior i osul palatin. Fuzionarea se face dinapoi nainte, ramnand fii suturale despritoare la nivelul crora bolta palatin va continua s creasc i dup nastere. Muchii aparatului dento-maxilar se difereniaz i se dezvolt n ultimele cinci luni de via intrauterin, mrindu-i volumul de 50 de ori, cea mai important dezvoltare o au muchii masticatori. Contraciile musculare, frecvente n ultimele luni ale vieii intrauterine, au rol important n stimularea dezvoltrii aparatului dento-maxilar. Craniul neural se dezvolt mult mai mult, comparativ cu cel visceral, sub impulsul dezvoltrii creierului, baza craniului se dezvolt mai ales n laime. deteminnd largirea feei i reducerea "botului". Coordonarea insuficient a creterii diferitelor segmente formatoare a complexului oro-facial determin fuzionarea necorespunztoare sau chiar insuficient de fuzionare a segmentelor. Aceasta determin "crpturi" ce deformeaz masivul facial. Crpaturile persistente de-a lungul liniilor poteniate de fuziune ntre segmentele embrionare i anornaliile sunt denumite dup criteriul localizrii. Cele mai frecvente defecte sunt crpaturile buzelor i palatului: "buza de iepure", "gura de lup". Crpatura palatinal singur are o inciden de 1 la 2500 de nateri vii afectnd mai ales sexul feminin. Craptura combinat a buzelor si palatului are incidena de 1 la 800 pan la 1 la 1000 de nateri vii, afectnd n proporie mai mare sexul masculin. Incidena
68 acestor anomalii este mai mare dac copii provin din printi care la randul lor au avut anomalii oro-faciale. Studii1e familiale privind fisura palatin arat ca o poriune a cromozomului sexual X este responsabil de apariia acestui defect genetic. Fisura bolii palatine poate s apar ca una din manifestrile altor anomalii genetice, rare. Macrostomia reprezinta insufucienta de fuziune a proceselor maxilare i mandibulare rezultnd o cavitate bucal mult largit. Invers, microstomia se datoreaz unei fuzionri precoce, realiznd o cavitate bucal micorat. 4.8.10. Creerea i dezvoltarea cranio-facial dup nastere La nastere prile constitutive ale cutiei craniene sunt separate ntre ele prin existena a 6 fontanele. Bosele frontale i parietale sunt foarte accentuate iar angulaia bazei craniului este foarte discret. Masivul facial, n raport cu neurocraniul, este nca putin dezvoltat i din aceast cauz copilul la nastere nu ii poate menine capul, care are tendina de cdere spre inapoi. Dup Chiari i Forceps raporturile fa/craniu sunt: 1/8 la nastere, 1/6 la 2 ani, 1/4 la 5 ani, 1/3 la 10 ani, 1/1 la 20 ani. Dup natere pn la pubertate neurocraniul i mrete volumul de 4 ori, iar faa se mareste de 12 ori. Raportat la talie, craniul reprezint: 1/4 la nastere, 1/5 la 1 an, 1/6 la 8 ani, 1/7 la 15 ani. 4.8.10.1. Dezvoltarea unor pri din neurocraniu Rata creterii extrauterine a volurnului cerebral i a craniului variaz astfel: volumul se dubleaz de la nastere dnd n luna a VI-a i se tripleaz pn la trei ani. Creterea craniului se face n toate sensurile pan la 8 ani, varst dup care anumite regiuni se opresc complet din crestere. Bolta cranian crete pe seama suturilor i a proceselor de apoziie/ resorbie de la nivelul tablelor. Forma cranian este determinat de: factori genetici, nchiderea prematur a unor suturi (sinostoze), aciunea unor factori deformani, constelaia hormonal, dezvoltarea creierului. Rolul creierului n dezvoltarea craniului este dramatic demonstrat n condiii patologice ca: hidrocefalie, microcefalie, anencefalie.
69 Baza craniului ndeplinete rol de fixare, de sprijin pentru scheletul facial. Creterea sa depinde de: dezvoltarea creierului, corelaiile legate de dezvoltarea facial, curba general de cretere, activitatea sincondrozei sfenooccipitale ca factor moderator al ajustrii bazei craniului la creterea creierului, respectiv la pasajul nano-faringian al aerului. Baza craniului se dezvolt, n special, la nivelul sincondrozelor sfenoetmoidal intrasfenoidal, sfenoocipital i intraoccipital. Etmoidul, os de natur encondral, se dezvolt foarte rapid, asigurnd protecia organelor de sim. Sutura sfenoetmoidal favorizeaz o rat foarte rapid de cretere pan la trei ani, dup care se inchide aproape complet, rata de cretere scade, oprindu-se definitiv la 7-8 ani. n perioada erupiei molarilor permaneni se poate relua activitatea de cretere la nivelul suturii sfenoetmoidale. Sfenoidul, prin poziia i raporturile sale, este considerat osul de baz al ntregului masiv cefalic. Creterea sfenoidului se face parial pe seama sincondrozelor, parial prin apoziie de os la suprafa, cretere modelat de forele ce se realizeaz la acest nivel. Sutura sfenooccipital are o activitate mai ndelungat, constituind principalul focar de alungire a bazei craniului. Sincondroza sfenooccipital se include dupa erupia ultimilor molari. Angulaia bazei craniului se apreciaz dupa unghiul sfenoidal (132 pentru zona european). S-a ncercat sa se stabileasc o corelaie ntre unghiul sfenoidal, pe de o parte, dezvoltarea facial i anomaliile dento-maxilare, pe de alt parte, precum i infiuena unor tratamente ortodontice asupra angulaiei bazei craniului. Osul frontal, formeaz peretele anterior al cutiei craniene, de asemenea, ia parte la formarea etajului superior al figurii, iar prin raporturile suturale pe care le are cu unele oase faciale, este n interdependen cu dezvoltarea cutiei craniene. Sinusul frontal este o structur dinamic, formarea i dimensionarea sa este n corelaie cu transmiterea forelor la nivelul aparatului dento-maxilar. Punctul nasion (intersecia dintre planul median al capului si sutura frontonazal) este foarte utilizat la examenul masivului facial. 4.8.10.2. Dinamica creterii unor pri a viscerocraniului Regiunea nazal, din punct de vedere al dezvoltrii poate fi imprit in dou zone: o zon superioar interorbitar, o zon inferioar, interrnaxilar, delimitate de o tangent imaginar la marginea inferioar a orbitelor. La nastere, zona interorbitar are o nlime dubl fa de cea intermaxilar, ca apoi,
70 la adult, cele dou zone s aibe o nltime aproximativ egal. Dezvoltarea transversal a celor dou segmente prezint ritmuri asemantoare celei verticale. Creterea sagital este corelat cu cea a oaselor din vecintate. Dezvoltarea orbitei se face foarte rapid, legat de dezvoltarea ochiului. Datorit relaiilor funcionale dintre orbit i aparatul dentomaxilar s-au stabilit unele puncte orbitare folosite n studiul i diagnosticul anomaliilor dentomaxilare (exemplu punctul infraorbital, planul Frakfurt la marginea inferioar a orbitei). Osul malar (zigontatic) prin forma i poziia sa face legatura dintre osul maxilar superior i osul frontal, temporal, asigurnd dirijarea transmiterii forelor ce apar n cursul funcionrii aparatului dentomaxilar. Pe partea sa interioar se inser muschiul maseter. Osul malar este legat de celelalte oase prin suturi care au o perioad indelungat de activitate. Ritmul de cretere al osului malar este reglat n funcie de influenele funcionale, contribuind la dezvoltarea facial si la meninerea unui echilibru ntre dezvoltarea cranial si cea facial. Arcada zigomatic este realizat prin participarea apofizei temporale a malarului i a apofizei malare a osului temporal. Diametrul dizigomatic este folosit pentru aprecierea: ritmului de dezvoltare a feei n sens transversal, concordanei dintre marimea dinilor i dezvoltarea facial, dezvoltrii arcadei superioare n funcie de dezvoltarea facial, variaiilor de ritm privind dezvoltarea facial n cele trei planuri spaiale. Distana dizigomatic serveste ca element de comparaie fa de care se apreciaz marimea dinilor (abaterile de la normal fiind interpretate ca macro respectiv microdonie relativ) i marimea arcadei superioare (abaterile fiind definite ca arcad ingustat sau arcad lrgit Maxilarul superior este ntr-o strns legatur cu celelalte oase faciale cu oasele de la baza craniului de care este unit prin numeroase suturi fibroase de tip armonic. Prin unirea celor doua pri pre i postmaxilare rezult o formaiune unic, simetric legat de baza craniului, care influeneaz dezvoltarea. Poziia maxilarului superior fat de baza craniului depinde de creterea de la nivelul sincondrozelor sfenoetrnoidal, sfenooccipital. Creteri dimensionale schimbri de poziie, n special datorit creterii septului nazal i a oaselor cu care se nvecineaz, determin rnodificarea raporturilor spatiale ale maxilarului superior. Dezvoltarea maxilarului se face n mai multe direcii: a. creterea n inlime se face prin: creterea sutural, apoziie de os alveolar i pe suprafaa palatinal inferioar, resorbia pe podeaua foselor nazale i electiv la cea a cavitilor orbitale. b. cresterea n lrgime se realizeaz prin:
71 creterea sutural, creterea apozitional de natur periostal i creterea divergent a apofizelor alveolare laterale. Datorit prezenei dinilor i a legaturilor cu maxilarul se creeaz la nivelul maxilarului o unitate anatomo-funcional particular fa de restul oaselor organismului. Astfel, se disting dou tipuri de os: osul maxilar propriu-zis i osul alveolar. Delimitarea exact a osului alveolar i a corelaiilor cu osul maxilar este destul de dificil datorit urmtoarelor considerente: imposibilitatea de stabilire exact a unei linii de demarcaie. Astfel, s-a propus separarea bazei maxilarului superior de partea sa alveolar, printr-un plan orizontal ce uneste apexul dintilor. Dar toate sistemele traiectoriale descrise att la maxilar, ct i la mandibul, nu conin elemente care s permit o delimitare net ntre partea alveolar i cea bazal. implantarea dentar i raportul dintre arcade pot avea rsunet la distant, n profunzimea structurilor. Raporturile incorecte intermaxilare din prognatismele mandibulare determin, pe de o parte, rmanerea n urm a dezvoltrii maxilarului superior, iar pe de alt parte se produce o dezvoltare exagerat, a sinusurilor frontale. Aceste modificari ale sinusurilor apar datorit unor particulariti n transmiterea spre cutia cranian a forelor ce iau nastere din contactele interarcadice. De asemenea, modificarea raporturilor interdentare i interarcadice determin largirea maxilarelor. partea bazal i cea alveolar a maxilarelor au o origine comun, fiind de natur osoas, ntre ele existnd o unitate de structur i continuitate: creterea lor depinde de aceeai surs sanguin i nervoas; formarea i apariia dinilor la marginea maxilarelor ca i prezena ligamentelor alveolo-dentare induce o activitate osteogenetic important; legatura specifiec os-dinte; stimulii variai i repetai ce iau nastere din contactele interarcadice, determin modificri ale structurii funcionale a osului inducnd o cretere mai accentuat n timpul eruptiei dintilor; dezvoltarea oaselor maxilare este influenat de o multitudine de factori locali (sistemul dentar, activitatea reflex muscular, crterea sutural, creterea periostal) i generali (factori metabolici, endocrini, genetici osul poate, la randul su, s influeneze sistemul dentar (obstacolele osoase pot modifica ritmul i direcia de eruptie a dintilor, pot forma deformaii radiculare). Alveola dentar definete spatiul sau cavitatea existent n osul maxilar, care gzduiete rdcina unui dinte. Fiecare alveol este mrginit de catre pereii alveolari, alctuii din substant osoas cortical (prelungire a corticalei externe osoase). n masa osoasa sunt nglobate extremitile fibrelor Sharpey, prin intermediul crora
72 primete forele ce se transmit de la dinte. Procesul alveolar definete corticala alveolar i o zon subire de os spongios care inconjoar rdacina unui dinte i care il insoete n toate deplasrile sale. Procesul alveolar este direct legat de formarea i creterea radcinii, se resoarbe odat cu rizaliza dinilor temporari, dispare la cateva sptmni dup extracia dentar. Dup extracia dentar, marginile restante ale maxilarelor sunt cunoscute sub denumirea de creste alveolare i ajut la realizarea masticaiei. Osul alveolar sau apofiza alveolar ar cuprinde zona osoas, care la maxilarul superior, se prezint ca o prelungire n jos i excentric a marginii sale. Limita sa superioar este situat la un plan raportat la apexul dinilor. La mandibul pentru partea anterioar a arcadei se folosete aceeai delimitare. n schimb, n regiunile laterale ale ramurii orizontale a mandibulei, datorit dezvoltrii compactei interne i externe, se formeaz un jgheab n care se gsete osul spongios, n care sunt spate alveolele. n aceste condiii, osul alveolar desemneaz zonele osoase situate deasupra liniei oblice externe i interne, dar limitele nu sunt bine distincte nici anatomic, nici funcional. Forele realizate din contactele interarcadice sunt transmise i influenteaz procesul alveolar, apofiza alveolar, osul bazal i se propag la nivelul celorlalte oase ale masivului cranio-facial. Forele musculare se exercit: prin intermediul dintilor; direct asupra esutului osos prin presiunea grupului de fibre musculare pe suprafaa osoasa sau ca rezultat al aciunii musculare la nivelul regiunilor de inserie. Creterea osului alveolar se face prin marire transversal a perimetrului, alungire posterioar a arcului alveolar i creterea n inlime. Caracteristicile creterii perimetrului sunt: resorbii la nivelul pereilor orali i apoziii osoase la nivelul pereilor vestibulari; intermitena de cretere dependent de apariia dentiiei i de creterea facial; dependena de tonusul muschilor interni i externi; la arcada superioar se face prin cretere sutural median care este foarte activ pan la 6-7 ani. La mandibul activitatea suturii simfizare nceteaz rapid, de aceea aici predomin procesul de resorbie i apoziie. Alungirea posterioar a arcului alveolar se face prin producerea de os nou, n legtur cu erupia molarilor: dezvoltarea tuberozitailor la maxilarul superior i a zonei retromolare la mandibula, asociate cu o micare puternic mezial ce determin o remaniere marcat a rebordurilor alveolare. Creterea cea mai marcat se face n sens vertical determinnd transformarea profilului copilului, prin alungirea etajului inferior al feei.
73 Bolta palatinal Centrul de osificare al osului palatin apare imediat dup cele ale maxilarului superior determinnd creterea n dou direcii: orizontal, spre palat, vertical, pe partea intern a peretelui capsulei nazale. Cele dou jumtai ale bolii se unesc ntre ele i cu septul nazal (absena fuziunii duce la apariia crapturilor). Creterea i modificarea formei bolii palatine este legat de creterea septului nazal, de activitatea sutural i dezvoltarea osului alveolar, Coborrea palatului se face prin resorbie pe suprafaa nazal i apoziie pe faa oral. Creerea mandibulei Din punct de vedere al dezvoltrii, mandibula se compune dup. Symons din: arcul propriu-zis, cu o parte orizontal paralel la palat i o parte vertical ce se termin n condil; structurile anexe, ataate la arcul propriu-zis: apofiza coronoid, pe care se inser muschiul temporal; unghiul posterior, pe care se inser muchii maseteri i pterigoidian intern; anexa orizontal ce serveste inseriei muschilor obrazului, muchilor planeului bucal; partea care cuprinde dinii. Componentele mandibulare pot fi clasificate, din punct de vedere funcional, n: os alveolar, os bazal i os muscular. Mandibula provine din material aparinnd primului arc branchial. Cartilajul Meckel se dezvolt n luna a 11-a intrauterin, n viaa intrauterin caracteristicile creterii mandibulei sunt: se pstreaz relativ constant proporia dintre lungimea total i cea a corpului mandibulei ("discul alveolar"); se pastreaz relativ constant proporia dintre deschiderea unghiului mandibulei i laimea total; corpul mandibulei prezint o cretere n largime mai rapid dect restul. La natere cele dou hemimandibule sunt scurte, iar eminena articular este abia schitat (fig. 13). Simfiza mentonier se prezint ca o linie subire de fibrocartilaj care se osific ntre lunile IV-XII postnatal, cnd cartilajul superficial este nlocuit cu os.
74
Fig. 13. Proporiile mandibulei la diferite vrste. (dup BOBOC, 1996, modificat) Zonele de cretere mandibular sunt: regiunea retromolar, care este n strans legatur cu formarea succesiv a molarilor; cartilajul condilian, care determin creterea vertical i orizontal influentnd: proporia dintre ramura orizontal i cea vertical; cantitatea de cretere n partea posterioar a ramurii verticale; cantitatea de os alveolar necesar s umple spaiul intermaxilar; apofiza coronoid, incizura sigmoid i regiunea unghiului mandibulei unde are loc o cretere apoziional dependent de activitatea musculaturii ridictoare mandibulare; osul alveolar: creterea este continu orientat n sus si n afar, avnd importan n stabilirea i meninerea relaiilor interarcadice ca i n conformaia facial de ansamblu; la nivelul periostului au loc fenomene de resorbie pe fata intern a mandibulei i apozitie pe faa extern. Dezvoltarea mandibului n cele trei dimensiuni spaiale se face astfel: alungirea se produce prin cretere la nivelul cartilajului condilian i apoziie pe marginea posterioar; dezvoltarea vertical: se realizeaz prin cretere la nivelul condilului, apofizei coronoide i cretere alveolar important; dezvoltarea transversal se realizeaz: prin creterea la nivelul simfizei mentoniere (n primul an); cretere apoziional pe toat faa extern (n corelaie cu resorbie pe faa intern); cretere apozitional la marginea distal a marginii verticale. Dezvoltarea articulaiei temporo-mandibulare Articulaia temporo-mandibular este deja schiat la embrionul de 8 sptamani, n trimestrul 11 de viat intrauterin sunt vizibile meniscul i muschiul pterigoidian extern. La natere, fosa glenoid este foarte superficial i se prezint ca o depresiune uoar de form aproximativ circular, iar condilul mandibular este abia schiat.
75 n primele luni de viat mandibula are o mare libertate de deplasare, sub aciunea muchilor mobilizatori. Din aceast perioad articulaia ternporomandibular sufer modificri succesive care duc treptat la: formarea i adncirea cavitii articulare a temporalului (cavitatea glenoid); apariia i dezvoltarea tuberculului articular anterior; dezvoltarea condilului mandibular; dezvoltarea i sistematizarea componentelor cartilaginoase i fibroase. Anatomia articulaiei temporo-mandibulare se schimb succesiv, dependent de factorii ereditari i factorii funcionali (elementele neuromusculare si fortele de ocluzie). Forele musculare influenteaz creterea mandibulei astfel: muchii pterigoidieni interni i externi, trag segmentul temporo mandibular nainte; muschiul temporal are o aciune predominant vertical; muschiul maseter imprim o direcie oblic nainte i in sus; muschiul sternocleidomastoidian orienteaz n jos i nainte apofizele mastoide; musculatura puternic a cefei, tinde s deplaseze osul occipital n jos, influennd activitatea la nivelul sincondrozei sfenooccipitale. Fora gravitaion influenteaz i ea activitatea diferitelor grupe musculare. Erupia dinilor, realizarea contactului ocluzal determin o coordonare mai complex a micrilor musculaturii, modificri morfologice ale elementelor osoase: creterea condilului; adncirea cavitaii glenoide; schiarea tuberculului articular anterior. Tratamentele cu aparate ortodontice influenteaz articulaia temporomandibular determinnd: resorbii pe suprafetele de contact cu presiune; apoziie compensatoare pe versantele opuse; deplasri consecutive restructurrii condilului i restului osului mandibular; modificri ale cavittii glenoide. 4.8.10.3. Dinamica dezvoltrii scheletului cavitaii orale n ansamblu Creterea vertical a cavittii orale este legat de: creterea vertical a maxilartilui superior; creterea mandibular dependena de dezvoltarea cartilajului Meckel i a cartilajului secundar condilian; creterea osului alveolar are o pondere mare, este corelat cu fenomenele dentiiei i cu distana ce se creaz intre scheletul facial superior i mandibul, osul alveolar tinznd s umple acest spatiu.
76 Creterea transversal a cavittii orale se realizeaz prin dezvoltarea la nivelul suturii intermaxilare i mediopalatine ca i prin dezvoltarea la nivelul suturii dintre premaxilar i postmaxilar. Factorii principali de dezvoltare transversal a cavittii orale sunt: procesul de resorbie de pe faa intern a mandibulei; creterea divergent a ramurilor ascendente; creterea divergenii n jos i in afara osului alveolar superior, ceea ce determin forma palatului. Palatul ingust este rezultatul creteri alveolare n sens vertical, iar palatul plat este rezultatul unei creteri alveolare orizontale. Creterea sagital (antero-posterioar) a cavitii orale se realizeaz prin: dezvoltarea septului nazal i activitaii suturale, care impinge inainte maxilarul superior; dezvoltarea mandibulei prin aportul cartilajului condilian, resorbtia marginii anterioare, apozitie pe marginea posterioar, translaia mandibulei; dezvoltarea osului alveolar in directia jos i inainte, apoziia osoas la nivelul tuberozitilor i translatia inainte a dentitiei.
4.9. METABOLISMUL FLUORULUI
Observatiile clinice au artat ca fluorul particip la mecanismele de prevenire a cariogenezei i pentru aceasta fluorul trebuie luat in timpul formrii emailului i continuat dup eruptia dinilor: pentru dinii temporari fluorul se administraz inainte de eruptie, iar pentru dintii definitivi adrninistrarea se face dup eruptie. 4.9.1. Aportul de fluor este de l mg/zi pana la maximum 2,5 mg/zi asigurat de alimente, ap, agenti farmacologici. Concentraia fluorului n alimente este de sub 0,5 prti per milion (ppm) ceea ce determin un aport zilnic de aproxirnativ 1 mg. Concentratia fluorului in ap trebuie s fie aproximativ 1 ppm. Alinentele bogate in fluor sumt: petele (are pielea bogata in fluor), ceaiul, conine 1 ppm de fluor ceea ce asigura un aport de 3 mg/zi la consumul a 15 cni de ceai. Dac ceaiul se face cu ap fluorinat aportul poate s ajung la 8 10 mg/zi. Pastele de dini pe baz de monofluorfosfat de sodiu asigur o absorbie de 3,015 0,030 mg fluor la fiecare spalat de dinti cu I gram pasta ce confine 1 mg F/gr. Aplicaiilc topice, locale de solutii de fluorur de sodiu 2%, geluri, amine fluorinate determin: - creterea continutului de fluorapatita la suprafata emailului, care devine mai rezistent; - remineralizarea emailului parial demineralizat; - diminuarea fermentaiei dulciurilor n cavitatea bucal, O alts surs ocazional de fluor sunt tabletele de fluorur de sodiu sau de calciu administrate in doze de 0,25- 1 mg F/zi.
77 4.9.2. Absorbtia fluorului se realizeaza pasiv, sub forma de fluoruri la nivelul stomacului si intestinului subtire. Fluorul din ape este usor absorbit. Absorbtia fluorului este diminuata de consumul de lapte, de prezenta in cantitti mari a calciului, magneziului, fosfatilor care pot forma cu fluorul saruri insolubile, putin absorbabile. In schimb citraii, fixatorii de calciu faciliteaz absorbia fluorului. 4.9.3. Fluorul in plasm este aproximativ 0,1 ppm ( 0,50 ppm) din care 10 - 20% este liber iar restul este fixat pe serum-albumine. In laptele matern (uman) i saliv fluorul este in concentratie asemanatoare cu plasma. 4.9.4. Depozitele de fluor in organism sunt: oasele scheletului si dintii. Fluorul are afinitate pentru os care reine 20-50% din fluorul de aport (1-20ppm). Fluorul substituie OH- din cristalele de hidroxiapatita, rezultand fluorapatita, conform reactiei: Ca 10(PC:14)5(011)2 + F- H Ca-1004)40HW + OHcare este mai putin sensibil la disoluie. Formarea fluorapatitei este un fenomen reversibil care depinde de: viteza reanoirii osului, gradul de vascularizatie, aprovizionarea cu fluor. Cantitatea de fluor in os creat n varsta. Din os fluorul poate fi mobilizat prin activitatea osteoclastelor, fluor care apoi este redepozitat sau eliminat. Timpul de injumatatire al fluorului din os este de 8 ani. Fluorul se acumuleaz in structurile dentare inainte de eruptie, proportional cu aportul de exogen, n momentul mineralizrii. Depunerea de fluor este de 2-3 ori mai mare in dentin dect in email. Dup mineralizarea emailului, fluorul se depune la periferie, concentratia lui fiind aici de 10 ori mai mare dect n profunzimea emailului. Dup erupie, dentina se imbogaete lent cu fluor, iar emailul de suprafata se concentreaz n fluor provenit din saliva, lichid gingival. La suprafaa emailului pot avea loc fenomene ciclice de demineralizare (datorit fermentaiei bacteriene) i reprecipitare, care depind de variatiile locale de pH 11 concentraia ionic n plac, pe dinti. Fluorul prezent faciliteaz reprecipitarea formarea fluorapatitei. Cristalele de fluorapatita de la suprafa pot avea un diametru de 2 ori mai mare decat cristalele din profunzime. Fluorul poate inlocui OH-, CO3-2 din cristalele de apatit. Se realizeaz la suprafaa emailului un depozit de fluor, mobilizabil in faza de demineralizare ca apoi s reprecipite formnd cristale de fluorapatita, mai rezistente. La baieti fixarea fluorului n dinti este cu 10% mai mare dect la fete. 4.9.5. Eliminarea flourului din organism se face prin rinichi, fecale glande sudoripare, lapte matern. In cazul unui aport constant, excreia renal este echivalent cu aportul. Fluorul filtrezta glomerular i este reabsorbit tubular n proportie de 60-90%. Scaderea pH-ului urinar determin reabsorbia crescut de fluor i incarcarea osoas excesiva n fluor. La gravide si la copii n crestere,
78 eliminnd pe cale renala numai 30-50% din cantitatea dc fluor ingerat. In fecale se elimin 10% din fluorul ingerat, cantitate ce crete n funcie de aport. Eliminarile pe cale sudoral sunt mici, pot crete n caz de transpiratii masive, dar prin aportul adecvat de lichide se asigur autoreglarea bilantulu fluorului. Laptele matern conine 0,02 - 0,05 ppm fluor legat de grasimi, albumine globuline sau cazein. La gravide, cantitti mici de fluor pot trece transplacenta i se incorporeaz in tesuturile dentare, osoase ale fatului. Fluorul vehiculat de saliva reprezinti numai 1% din fluorul ingerat. Lichidul gingival conine fluor mai mult decat n plasma. 4.9.6. Rolurile fluorului in organism sunt: crete stabilitatea cristalelor de apatite prin inlocuirea OH- cu F rezultnd fluorapatita i astfel scade solubilitatea la acizi a acestor cristale din o: sau tesuturi dentare; scade incidena cariilor atat prin incorporarea n tesuturile dentare, n particular n stratul superficial al emailului cat i prin concentrarea fluorului in placa dentar bacterian. Formarea cristalelor de fluorapatita la suprafata emailului asigur rezistena crescuta a acestuia la atacul acid. La nivelul plcii dentare bacteriene cantitatea de fluor este de 70 280 ori mai mare decat n saliv. n plac, fluorul se gasete fie liber n faza apoas extracelular, fie legat de constituieni anorganici, de produse dializabile eliberate de scaderea locala a pH-ului, de bacterii. Placa dentar se poate mbogati n fluor din: alimente, apa, de pe suprafata emailului,saliv, lichid gingival.Fluorul din placa dentar perturb aderena bacteriilor pe email diminu sinteza polizaharidelor intra, extracelulare,inhib producerea de acizi. S-a crezut ca fluorul inhib enolaza, enzima ce particip la glicoliza i astfel diminu producerea de acizi. Fluorul inhib: peroxidaza, catalaza si activeaz adenilat ciclaza, favoriznd astfel formarea AMP, i efectele hormonilor dependeni de AMP,. 4.9.7. Toxicitatea fluorului Intoxicaia acut cu fluor se insoeste de diaree, vom, dureri abdominale, Ingestia cronic de fluor peste 20 mg/zi, n caz de coninut crescut de fluor n ap sau la cazurile de hemodializat cronic, duce la intoxicaia cronic numita fluoroza. Fluoroza osoas se caracterizeaz prin: acumularea de fluor in os cu densificri osoase; creterea volumului osteoidului; scderea mineralizarii; degenerescena osteocitar, i n final
79 osteomalacia prin efecte citotoxice osoase. Fluoroza dentar se insotete de prezenta plcilor opace pe email, zone de hipomineralizare sau cu porozitate crescut n dini.
FIZIOLOGIA LIMBII
5.1. LIMBA
5.1.1. Structura limbii Limba este un organ foarte mobil, capabil s execute micri de mare finee i varietate. Limba este un organ muscular, acoperit de mucoasa. Limba ocup aproape n intregime cavitatea bucal propriu-zis. Ea se rnuleaza pe suprafata palatinal a dinilor, pe care tinde vestibulizeze. Limba are un schelet fibros, reprezentat prin membrana hioglosian i prin septul median ce se intinde de la apofizele geniene la membrana hioglosian. Osul hioid ofer insertii muschilor extrinseci ai limbii. Limba este alctuit din dou grupuri musculare: grupul muscular intrinsec format din muschii: longitudinal superficial, longitudinal profund, transvers si vertical. Acestia incep si se termin liberi in limb si nu au insertii osoase. Prin contracia muschilor intrinseci se produce turtirea limbii. grupul muscular extrinsec format din: genioglos, hipoglos, stiloglos, glosopalatin. Aceti muschi se inser pe oasele craniului, mandibul, iar celelale capete se termin in limb. Muschii extrinseci au rolul de a fixa scheletul fibros al limbii, pentru a permite muschilor intrinseci sa se contracte i s deplaseze limba n toate direciile. Muschii extrinseci sunt reprezentai de cinci perechi musculare: muchii genioglosi, asigur, prin contractia lor, scoaterea limbii in afar; muschii hipoglosi, prin contractia lor mic limba in jos i inapoi; muschii stilogloi, asigur micrile limbii n sus i inapoi; muschii faringogloi i muchii palatogloi, regleaz deschiderea istmului gatlejului i faringelui. n muchi se gsesc numeroase fusuri neuromusculare. Mucoasa limbii are un epiteliu foarte gros i un corion dezvoltat, bogat vascularizat i inervat. Mucoasa de pe fata dorsal este mai groasa i mai ferm., iar cea de pe fata inferioara este mai subtire i mai elastic La nivelui unirii treimii posterioarc cu dou treimi anterioare exist dou anuri oblice ce formeaz "V"-ul lingual n care sunt adpostite 9 11 papile caliciforme. De asemenea, pe faa dorsal a limbii se
80 gasesc papilele gustative. filiforme, foliate, fungiforme. napoia "V"tilui lingual, mucoasa este subtire si inegal mamelonat, datorita foliculilor limfatici, care formeaz amigdala lingual i datorit glandelor foliculare seroase, mucoase. Faa inferioar a limbii este acoperit cu o mucoasa subtire, care prezint, median, frul limbii, ce face legtura cu arcada alveolar inferioar. Lateral de frul limbii se remarc: venele ranine, conductele excretoare ale glandelor sublinguale. Mucoasa lingual conine, pe lang receptorii specifici gustativi receptori tactili, termici, durerosi. Vascularizaia limbii este asigurat de arterele linguale ale cror ramificaii formeaz plexuri sub mucoasa lingual irignd att mucoasa cat i muschii. Sangele venos este drenat prin venele linguale spre vena jugular intern. Capilarele limfatice se formeaz n jurul papilelor i dreneaz limfa spre trei grupuri ganglionare: ganglionii subhioidieni; ganglionii submaxilari; ganglionii jugulari interni. Proiectia cortical a limbii se realizeaz pe o suprafa relativ mare, datorit densitaii crescute de receptori i varietaii mari. Varful limbii, buzele, pulpa degetelor, prin bogia de receptori de la nivelul lor, particip la realizarea simtului tactil i de stereognozie. 5.1.2. Inervaia Inervaia are o component senzitiv, senzorial i motorie (fig. 14). Inervatia mucoasei linguale este asigurat de ramuri din cinci nervi cranieni: nervul trigemen, prin nervul lingual, inerveaz 2/3 anterioare ale mucoasei linguale; nervul facial, prin nervul coarda timpanului, inerveaz receptorii gustativi din 2/3 anterioare ale mucoasei linguale; nervul glosofaringian, inerveaza 1/3 posterioar a mucoasei linguale, att senzitiv ct i senzorial pentru papilele caliciforme i foliate (fig. 15); nervul vag, prin laringeul superior trimite ramuri senzitive i vegetative pentru regiunlie epiglotica si glosoepiglotica; Impulsurile aferente ajung la nucleul tractului solitar din bulb si/sau complexul senzorial al trigemenului din bulb si maiduva cervical superioar. nervul hipoglos inerveaz muchii
81
Fig. 14. Inervatia senzitivo-senzorial i motorie a limbii (dup ATKINSON, 1997, modificat)
Fig. 15. Inervaia senzitiv a limbii (dup GANONG. 1993. modificat) Nervul hipoglos ii are originea in nucleul hipoglosului, situat in bulb. Inervaia limbii este ipsilateral, adic muchii din partea dreapt a limbii primesc fibre motorii cu origine din aceai parte a nucleului hipoglosului. Nucleul hipoglosului are o anumit somatotopie, care asigur coordonarea micarilor specifice limbii. Limba realizeaz miscari complexe: protractie, retractie, deviere lateral, rsucire si ridicarea varfului limbii. Leziunea unilateral a nervului hipoglos duce la atrofia limbii, devierea limbii, tulburdri de masticatie, deglutitie, fonatie.
82 5.1.3. Funciile limbii rolul gustativ, realizat prin receptorii gustativi specifici, din mucoasa lingual (vezi Analizatorul gustativ); limba este un important organ tactil, avand o mare capacitate de discriminare a marimii, formei i structurii de suprafaa a obiectelor. De aceea, detecteaz cu uurin orice neregularitate a suprafetelor din cavitatea bucala. La sugari, particip la realizarea simului tactil si stereognozic. functia de aparare: prin receptorii gustativi, termici, tactili, de pe suprafaa limbii se detecteaz cele mai mici modificari fizico-chimice ale substantelor introduse n cavitatea bucal i astfel se protejeaz tractul gastrointestinal fa de noxe, de substane cu gust neplacut, de temperaturi extreme. Deglutiia se produce numai cand senzorii linguali arat c temperatura, consistena i gustul mncrii este acceptabil. particip la masticatie prin prehensiunea alimentelor, presarea alimentelor pe bolta palatin mentinerea lor pe arcadele dentare, formarca bolului alimentar. Prin miscarile precise ale limbii alimentele din vestibul sunt returnate ntre arcadele dentare i de asemenea are loc curairea vestibulului de particolele mici. Limba ii poate modifica forma adaptand-o la procesul de masticatie. Exist o coordonare foarte complex i delicat ntre micarile limbii si ale mandibulei, care ferete limba de actiunea fortelor mari ce se pot dezvolta ntre suprafetele ocluzale. Reflexele ce stau la baza coordondrii micarilor limbii cu micrile mandibulei, pornesc de la receptorii din muschii masticatori sau din mucoasa lingual. De asemenea, n aceasta reglare intervin fusurile neuromusculare ce se gesesc in numr mare pe partea cea mai flexibil a limbii. Deschiderea pasiva a gurii determin contracia limbii cu indoirea varfului spre planeul bucal. Impulsurile nociceptive de la nivelul limbii induc reflexul de deschidere a gurii. participa la timpul bucal i faringian al deglutitiei (vezi capitolul Fiziologia deglutitiei); particip la procesele de fonaie, vorbire. Precizia micarilor este necesar pentru articularea sunetelor in timpul vorbirii. In acest sens,limba este controlat riguros prin mecanisme de feed-back pornite de la proprioceptorii muschilor limbii i prin participarea sistemului nervos central. Informatiile senzitive pornite de la limb sunt, de asemenea, importante, de aceea, vorbirea este perturbat n urma pierderii sensibilitatii ca urmare a anesteziei nervului lingual, n timpul unor proceduri dentare de rutina. Marimea limbii i forma pe care o exercita, contribuie la mentinerea unor relatii ocluzale normale a dintilor. Pozitia dintilor este determinat, in parte, si de fortele musculare opozante ale limbii i buzelor. Mucoasa lingual ca i alte zone ale cavittii bucale este susceptibil la numeroase boli. Leziuni de diferite cauze, ale nervilor
83 senzitivi sau motori, pot determina scaderea inervatiei limbii. Astfel lezarea nervului lingual determin pierderea sensibilitatii a 2/3 anterioare a limbii si a inervatiei parasimpatice a glandelor sublinguale i submandibulare ipsilateral. Perturbarea secretiei salivare este minor atat timp cat celelalte glande salivare functioneaz normal. Pierderea ins a sensibilitatii gustative poate produce un gust metalic, neplacut, in gur. Dac un nerv hipoglos este lezat se produce paralizia flasca a muchilor limbii. Diminuarea funciilor senzitivo-senzoriale sau motorii ale limbii determin perturbarea variabila a vorbirii, masticatiei, deglutitiei.
5.2. ANALIZATORUL GUS'I'ATIV
Simtul gustului asigur aprecierea calitatilor sapide ale alimentelor i substantelor solvite in saliva. Functia gustativ este asigurat de: receptorii specifici din cavitatea bucal, caile gustative de conducere si segmentul central cortical al analizatorului gustativ. 5.2.1. Receptorii gustativi sunt de tip chemoreceptori, localizati n papilele gustative, au forma ovoid .Papilele gustative sunt de patru forme: calciforme, mari, 8-12 dispuse in "V" lingual, la baza limbii fungiforme, situate la varful i in 2/3 anterioare ale limbii; foliate, situate pe marginea limbii, in numar de 150; filiforme, numeroase, pe faa dorsal a limbii (fig. 16). Receptorii gustativi sunt reprezentai de mugurii gustativi (din papilele gustative), contin 5-40 celule gustative, alungite ce prezint la extremitatea apical microvilozitati (cili gustativi), care ajung prin porul gustativ la suprafata limbii (fig. 17). Microvilozitatile vin in contact cu substantele dizolvate n saliva. Polul bazal al celulelor gustative este inconjurat de fibre nervoase ale nervilor cranieni VII, IX. X, ce care realizeaz sinapse. De esemenea, mugurele gustativ conine celule de sustinere, plate i celule bazale, bogate in mitocondrii, celule ce nlocuiesc dup 10 zile celulele senzoriale.
84
Fig. 16. Distributia mugurilor si papilelor gustative pe limba (dupa BRAY, 1989, modificat)
Fig. 17. Structura mugurelui gustativ (dupa BRAY, 1989, modificat) Fibrele aferente ale mugurilor gustativi, la origine amielinice, formeaz n jurul celulelor senzoriale plexul intragemal, sub membrana bazal formeaz plexul subgemal, iar in jurul mugurelui gustativ realizeaz plexul perigemal, dup care devin mielinice si alcatuiesc nervii gustativi. 5.2.2. Calea gustativ Impulsurile gustative de la nivelul celor 2/3 anterioare ale 1imbii iau calea nervului coarda timpanului (ramur a nervului facial, VII), iar cele de la niveluI 1/3 posterioare iau calea nervului glosofaringian (IX) (fig. 18).
85
Fig. 18. Inervaia gustativ a limbii (teritoriul nervilor VII; IX) (dupa BRATU, 1993, modificat) Primul neuron este localizat in ganglionul geniculat pentru coarda timpanului si in ganglionul Andersch pentru glosofaringian. Al doilea neuron (deutoneuronul) este situat n nucleul gustativ din tractul solitar bulbar. Impusurile de la baza limbii, faringe, palat moale, epiglota, laringe, ajung pe calea nervului vag tot n tractul solitar din bulb. De la nucleul solitar, calea gustativ trece de partea opus i urca in lemniscul medial pan la nucleul ventro-postero-median al talamusului, unde face sinaps cu al III-lea neuron (fig. 19).
86
Fig. 19. Diagrama cailor gustative (dupa GANONG, 1997, modificat) 5.2.3. Proiectia cortical (segmentul central) este localizat n partea inferioar a circumvolutiei parietale ascendente (aria 43), in apropiere de centrii senzoriali i motori ai limbii, musculaturii masticatorii si actului deglutitiei. Caile reflexe gustative isi au originea la nivelul tractului solitar, de unde pornesc colaterale, fie direct, spre centrii salivatori superiori i inferiori (care prin impulsurile eferente catre glandele salivare controleaz secreia salivar in timpul digestiei bucale), fie indirect, prin ramuri catre nucleul reticular al formatiunii reticulate, de unde, pe calea tractului reticulo-bulbar, ajung fibrele eferente atat la glandele salivare cat si la muschii pieloi ai fetei, muschii masticatori, muschii limbii, muschii deglutiiei, coordonand miscrile asociate senzatilor gustative. 5.2.4. Stimularea mugurilor gustativi Excitantii adecvati ai sensibilitatii gustative sunt constituiti de proprietatile chimice ale diferitelor alimente. Excitanii substantele sapide, trebuie sa indeplineasc anumite conditii pentru a stimula receptorii gustativi: solubilizarea intr-un mediu lichid (saliva are rol de solvent, de diluare, de curire a cavitatii bucale dupa contactul
87 substantelor cu mugurii gustativi), sa actioneze sub forma de solutie ionizat de sruri sau acizi, sa aib o anumit temperatur (optima 38C), rdspandirea i omogenizarea substantelor pe suprafata receptoare. Cand substantele actioneaza asupra limbii imobile durata de stimulare este mai mare (receptie pasiv), iar cand actioneaz in timpul miscrilor limbii (receptie activ), perioada de latena este redusa deoarece se asigur o dizolvare mai rapid a substantei, o distributie pe suprafata mai mare, un contact mai bun cu receptorii. Receptorii gustativi pot fi stimulati si de substantele sapide introduse n circulaia sanguin (exemplu decolina folosita pentru determinarea timpului de circulatie, induce gustul amar). n procesul de excitare al mugurilor gustativi se disting dou faze: faza prenervoas de activare a receptorilor prin traversarea porului gustativ de ctre substanta sapid. Porul actioneaz ca un diafragm n reglarea cantitaii substanei sapide ce patrunde n mugure, prin interactiunea cu proteinele celulare din membrana porilor. Grupurile tiolice micoreaz porul, oligoelementele Cu+2, Zn+2, dilat porul. faza nervoas, cand substana sapid vine in contact cu microvilii celulelor senzoriale, induce .formarea unui potential de receptor care, dup atingerea valorii prag, genereaz potentialul de aciune ce se propag prin nervii gustativi la centrii nervoi. Prin interaciunea unei pari din substana sapid cu o protein specific de pe membrana receptorului gustativ, crete permeabilitatea membranei pentru Na+, se induce astfel un potential de receptor. Mecanismele prin care moleculele din solutie produc potentiale generatoare, difer de la substanta la substanta. Substantele srate, depolarizeaz probabil celulele receptoare pentru gustul srat, printr-un influx pasiv de Na+ prin canale apicale. Acizii, depolarizeaz celulele receptoare prin H+ blocnd canalele apicale de K+ ce determin gustul dulce, se leaga de receptorii membranari cuplai cu proteina Gs, activand adenilat ciclaza i rezultnd creterea AMPc. AMPc actioneaz prin intermediul unei proteinkinaze A care fosforileaz canalele de K+ de pe membrana bazo-lateral, determinand reducerea conductanei pentru depolarizarea membranei. Substanele ce determin gustul amar acioneaz pe calea receptorilor cuplai cu proteina Gq i fosfolipaza C care hidrolizeaz fosfatidil inozitol difosfatul (PIP2) n inozitol trifosfat (IP3) i diacilglicerol. Inozitol trifosfatul (IP3) declaneaz eliberarea de Ca+2 din reticulul endoplasmatic. Hidroliza PIP2 se asociaz cu deschiderea de canale cationice. Recent s-a izolat o nou subunitate a proteinei G numit a gusducin care se gsete n mugurii gustativi. - gusducin se aseamn cu transducina (proteina Gti), care
88 inactiveaz GMPc n receptorii retinieni. GMPc acioneaz direct pe canalele de Na+ meninndu-le n poziie deschis. Scderea GMPc determin nchiderea canalelor i hiperpolarizarea receptorilor retinieni. In timpul alimentrii sunt stimulai mai muli receptori (olfactivi, tactili, termici, proprioceptori), ns informaia specific cea mai mare este adus de la receptorii gustativi. Se percep 4 gusturi fundamentale (dulce, acru, amar, srat) iar prin combinarea gusturilor fundamentale se realizeaz o mare varietate de senzaii gustative. Gustul dulce se percepe cu vrful limbii, este dependent de gruprile chimice OH-, COO-, de prezena ionilor metalelor grele (plumb, beriliu) i de o anumit conformaie spaial a substanei. Gustul acru este produs de acizi, iar intensitatea senzaiei este aproximativ proporional cu logaritmul concentraiei de H+ . Totui acizii organici sunt mai acri comparativ cu cei minerali. Gustul srat este dat n special de cationii srurilor ionizabile: Na+, NH4+, Ca++, Li++, K+. Gustul amar este dat de unii alcaloizi (morfina, stricnina, chinina), de glicozizi, de sruri de Mg++, Ca++, de sruri biliare (gustul amar din icter rezultat n urma difuziunii srurilor biliare sanguine la nivelul mugurilor gustativi). Prin excitaia receptorilor gustativi cu un stimul neadecvat (curent electric) se pot obine senzaii gustative nespecifice. Pragul sensibilitii gustative reprezint concentraia minim de soluie necesar pentru a genera senzaii gustative. Pragul sensibilitii gustative depinde de temperatur, lumin, suprafaa de aplicare, durata aplicrii, graviditate, prezena senzaiei de foame, modificri ale compoziiei sngelui, factori de ordin psihofiziologic. Pragul sensibilitii gustative este pentru NaCl 0,01 M, HCl 0,0009 M, zaharoza 0,01 M, chinina 0,000008 M. Senzaia de foame crete acuitatea gustativ dar cnd nu este satisfcut scade pentru dulce, srat i crete pentru amar. Adaptarea gustativ - sub aciunea prelungit a unui stimul diminua pn la dispariie senzaia gustativ, (fenomen ce apare mai rapid pentru substanele srate i dulci). Adaptarea gustativ este proporional cu intensitatea stimulului; Adaptarea este urmat de procesul invers, de restabilirea sensibilitii. Cel mai repede se restabilete gustul srat, iar cel mai ncet gustul amar. La contientizarea gustului particip i sensibilitatea olfactiv, tactil, termic, dureroas a regiunii faciale ca i sensibilitatea proprioceptiv a muchilor masticatori. Aceasta se datoreaz stimulrii receptorilor respectivi din regiunea cilor reflexe gustative de la nivelul tractului solitar bulbar, ct i din proiecia gustativ cortical alturat sensibilitii respective, artnd relaiile de reciprocitate dintre ele.
89 Explorarea funciei gustative se poate realiza pe ntreaga suprafa a limbii sau pe papile izolate. Se pot folosi soluii apoase pentru cltirea gurii, pentru tamponarea limbii, sau stimuli electrici. Explorarea funciei gustative are importan pentru verificarea unor aptitudini profesionale (gustarea de alimente, vinuri), pentru diagnosticul tulburrilor de sensibilitate gustativ. Rolul sensibilitii gustative: - selectarea preferenial a hranei n funcie de dorina i nevoile organismului; - mpiedic ingerarea substanelor nocive, a alimentelor alterate; - declanarea reflex a secreiilor digestive; - aprecierea calitii unor produse alimentare (degusttorii de vinuri); - mbogete viaa psiho-afectiv a individului, etc.
5.2.5. Modificri ale senzaiei gustative: - cantitative: - hipogeuzie (scderea gustului), - ageuzie (lipsa sensibilitii gustative); - calitative: - disgeuzia (gustul este greit sau confundat). Aceste modificri pot s apar datorit unor tulburri endocrine, unor procese locale (stomatite, traumatisme, arsuri chimice, termice ale limbii, cavitii bucale, acoperirea palatului cu proteze dentare), unor leziuni ale cii de conducere (paralizia facial infecioas, traumatic, paralizie a glosofaringianului), leziuni ale centrilor corticali integratori ai senzaiei gustative.
FIZIOLOGIA ANALIZATORULUI OLFACTIV
La multe animale, simul mirosului (olfacia) este important pentru selecia hranei, activitatea sexual, recunoaterea altor animale, recunoaterea teritoriului. La om, acest sim are roluri n: - selecia hranei, - activitatea sexual i instinctual, - adaptarea la mediu - intervine n protecie i recunoaterea mediului, - mbogirea vieii psihice a individului prin reacii afective pozitive sau negative, - activitatea secretorie i motorie reflex a tractului digestiv.
90
6.1. RECEPTORII OLFACTIVI
Receptorii olfactivi sunt receptori de natura chimic, telereceptori, localizai ntr-o regiune specializat a mucoasei nazale
(fig. 20). Receptorii olfactivi sunt constituii din: a) neuroni bipolari (celule olfactive) - situai la nivelul mucoasei cornetului superior care reprezint mucoasa olfactiv, de culoare glbuie, avnd la om suprafaa de 5 cm2. Neuronii sunt fusiformi i prezint: - o prelungire periferic, ajungnd pn la suprafaa mucoasei, care se termin cu o poriune dilatat ca o cup, numita vezicul olfactiv, din care ies cilii olfactivi (10-12 cili/neuron receptor); - o prelungire central ce intr n alctuirea nervului olfactiv; b) celule de susinere - secret mucus, au microvili (fig. 21).
n mucus exist una sau mai multe proteine fixatoare de substane odorante (OBP) care: fixeaz odorantul, l concentreaz, l transfer receptorilor.
91 Pe cilii celulelor olfactive se gsesc receptorii specifici pentru substanele odorante. Receptorii sunt cuplai cu proteina G, asemntoare cu Gs. Proteina G face legtura receptorilor cu adenilat ciclaza care transform ATP n AMPc ce se leag i deschide canalele cationice, permind intrarea Na+ n celulele receptoare, care vor fi depolarizate (fig. 22).
6.2. CILE DE CONDUCERE
Celulele olfactive reprezint protoneuronul cii olfactive, avnd rol n recepie i conducere: axonii celulelor olfactive formeaz nervul olfactiv, strbat lama ciuruit a osului etmoid i merg la bulbul olfactiv (centrii olfactivi primari) unde fac sinaps cu deutoneuronul, reprezentat de celulele mitrale i celulele n pensul din bulbul olfactiv; de aici fibrele tractului olfactiv merg la centrii olfactivi secundari din: cortexul prepiriform, care reprezint proiecia specific principal, cortexul periamigdalian, hipocamp (fig. 23).
92
n centrii olfactivi secundari se formeaz senzaia olfactiv grosier. De aici, pornesc fibre spre centrii corticali teriari localizai n lobul temporal i cortexul prefrontal. Aici n asociaie cu funciile: vizual, tactil, somatic, vegetativ se formeaz senzaia olfactiv contient. Cile olfactive reflexe (corticofuge) leag hipocampul (rinencefal) cu centrii din diencefal, mezencefal, nucleii trunchiului cerebral, formaiunea reti-culat, determinnd micrile reflexe i rspunsurile vegetative la stimulii olfactivi, care includ activitile automate alimentare sau rspunsurile legate de anumite stri emoionale (frica, excitaia, plcerea, comportamentul sexual).
6.3 CARACTERISTICILE STIMULILOR OLFACTIVI
Excitanii olfactivi sunt de dou tipuri: - specifici (adecvai) - substane odorifice ai cror vapori stimuleaz nemijlocit celulele olfactive;
93 - nespecifici (inadecvai) - substane chimice inodore n sine, dar care exercit o anumit aciune asupra celulelor epiteliale (excit terminaiile nervului V): substane iritante ce produc strnut, lcrimare, inhibarea respiraiei. Pentru a fi excitani olfactivi, substanele trebuie s ndeplineasc urmtoarele condiii: - s fie volatile - creterea volatilitii intensific excitaia; - s se dizolve n mucusul ce acoper epiteliul olfactiv; - s se dizolve n lipidele din compoziia protoplasmei celulare; - s aibe o anumit presiune pariala a vaporilor; - s difuzeze rapid; - s fie absorbabile. Substanele odorifice pot ajunge n cavitatea nazal pe calea aerului inspirat (n cazul adulmecrii), prin aerul expirat sau pe calea sanguin. Substanele odorifice se mpart n trei grupe: - substane cu structur asemntoare i miros asemntor, - substane cu structur asemnntoare dar miros diferit, - substane cu structur diferit i miros asemntor. CLASIFICAREA MIROSURILOR Se deosebesc 6 senzaii olfactive principale (prisma olfactiva Hensing): mirosuri de flori, fructe, putrefacie, condimente, prjire, rin. Senzaiile olfactive sunt apreciate prin tonul emoional - afectiv, plcut sau neplcut.
6.4. DINAMICA SENSIBILITII OLFACTIVE
Aparatul olfactiv este sensibil, reacioneaz la concentraii foarte mici de substan. Pragul de sensibilitate olfactiv se msoar cu olfactometrul. Factorii de care depinde pragul de sensibilitate olfactiv: - variaz de la o substan la alta; - variaz n funcie de gradul de concentraie, de persoan; - lumina - stimuleaz olfacia; - temperatura: - optima 37-38C; - creterea temperaturii determin scderea sensibilitii olfactive; - umiditatea - n aer uscat concentraia substanelor trebuie s fie mai mare; - vrsta - sensibilitatea olfactiva este n cretere ntre 6 i 25 de ani; dup aceast vrst scade treptat; - foamea - crete pragul de sensibilitate olfactiv;
94 - saietatea - scade pragul de sensibilitate olfactiv; - sexul - bieii au acuitate olfactiva mai sczut dect fetele - ovulaia, graviditatea - cresc acuitatea olfactiv; - interaciunea mirosurilor - n amestec acestea pot fi percepute: simultan, difereniat, anihilat de altele, slbit; - stri patologice: - hiposmie, anosmie (n boli ale nasului, lezarea mucoasei olfactive); - hiperosmie (n graviditate, halucinaii olfactive n tumori, boli psihice).
6.5. ADAPTAREA SENSIBILITII OLFACTIVE Mirosul este nalt i rapid adaptabil, prin: - scderea sensibilitii n cursul aciunii substanelor odorifice - restabilirea sensibilitii dup ncetarea aciunii substanelor odorifice. Adaptarea poate fi omogen (pentru o substan), sau heterogen, difereniat n cazul unui amestec de substane.
SENSIBILITATEA GENERAL A CORPULUI I OROFACIAL
7.1. SENSIBILITATEA GENERAL A CORPULUI
Sensibilitatea general a corpului este asigurat de urmtoarele elemente structural funcionale: receptorii senzoriali, segmentul de conducere i segmentul central cortical. Receptorii senzoriali sunt celule nervoase specializate sau structuri celulare aneurale ce recunosc, selecioneaz, acomodeaz n intensitate i transform informaiile coninute n diveri stimuli ai mediului, n impulsuri nervoase. Deci, receptorii transform formele variate de energie mecanic, termic, n impulsuri nervoase. Receptorii se pot clasifica n: - exteroceptori ce cuprind receptori ai sensibilitii superficiale cutanate (termici, tactili, dureroi) i telereceptori (auz, vz, miros); - proprioceptori responsabili de sensibilitatea profund; - interoceptori ce pot fi: baroreceptori, chemoreceptori, osmoreceptori, etc. Segmentul de conducere asigur conducerea impulsurilor nervoase de la neuronii senzitivosenzoriali la centrii nervoi superiori. Este asigurat de trei neuroni:
95 - primul neuron (protoneuronul - I) i are originea n ganglionii spinali; - al doilea neuron (deutoneuronul - II) este situat n coarnele posterioare ale mduvei spinrii, respectiv nucleii Goll i Burdach din bulb. De obicei, axonul celui de al doilea neuron se ncrucieaz (trece de partea opus); - al treilea neuron (III) este situat n talamus. Sensibilitatea tactil ( exteroceptiv ) este asigurat de calea spinotalamic ventral (are deutoneuronul n coarnele posterioare ale mduvei) i calea spinobulbar (are deutoneuronul n nucleii Goll, Burdach). Sensibilitatea termic i dureroas (exteroceptiv) este asigurat de calea spinotalamic dorsal. Sensibilitatea proprioceptiv contient este asigurat de calea spinobulbar (II neuron n bulb). Sensibilitatea proprioceptiv incontient are primul neuron n ganglionul spinal, al doilea neuron n coarnele posterioare ale mduvei de unde urc, fie de aceeai parte (fascicolul spinocerebelos direct Flechsig), fie de partea opus (fascicolul spinocerebelos ncruciat Gowers), spre scoara vermisului cerebelos. Segmentul central este reprezentat de cortexul parietal (aria somato-senzorial primar S1) unde are loc decodificarea informaiei din impulsurile nervoase, realizarea percepiei i prelucrarea ei n corelaie cu experiena acumulat anterior de subiect. 7.1.1. Sensibilitatea tactil, vibratoare i de presiune general Tactul, presiunea, vibraiile sunt senzaii separate, dar determinate de intensiti diferite ale aceluiai stimul de baz. 7.1.1.1. Receptorii sensibilitii tactile sunt de tipul mecanoreceptorilor, descriindu-se 6 tipuri (fig. 24): - terminaii nervoase libere din epiderm i derm, care recepioneaz durerea, tactul, presiunea;
96
- corpusculii Meissner din derm, sunt foarte numeroi n vrful degetelor zona palmei, vrful limbii, buze, faa plantar a picioarelor. Au rol n sesizarea caracteristicilor spaiale ale obiectelor, sunt sensibili la atingerile fine; - corpusculii Merkel, sunt situai n epiderm, zone acoperite cu pr, sunt stimulai de atingerile puternice; - terminaiile nervoase amielinice din jurul foliculilor piloi, sesizeaz micrile de ndoire a firului de pr; - corpusculii Ruffini sunt localizai n hipoderm, sunt receptori specializai pentru cldur; - corpusculii Pacini sunt localizai n esutul subcutanat, muchi, articulaii, detecteaz presiunea, vibraiile care produc modificri rapide n starea mecanic a esuturilor (ndoire, deformare, elongare). Capacitatea de discriminare tactil a tegumentelor depinde de densitatea receptorilor pe unitatea de suprafa fiind maxim pe pulpa degetelor (2,3 mm) i minim pe spate, coapse (67 mm). Topognozia reprezint capacitatea de localizare a unei excitaii tactile de ctre subiect. Dermolexia reprezint capacitatea de recunoatere a literelor, cifrelor scrise cu un ac bont pe tegumente. Stereognozia reprezint capacitatea de recunoatere cu ochii nchii a obiectelor plasate pe tegumente.
97 7.1.1.2. Cile de conducere sunt reprezentate de: - protoneuronul localizat n ganglionul spinal; - deutoneuronul: localizat n cornul posterior medular, de unde pleac calea spino-talamic ventral i dorsal (conduc sensibilitatea protopatic tactil, de presiune, termic i dureroas). Deutoneuronul poate fi localizat i n nucleii Goli i Burdach, care primesc aferene pe calea spino-bulbar, ce conduce informaii privind tactul epicritic, senzaiile vibratorii i de presiune; - al III-lea neuron se gsete n nucleii talamici (fig. 25).
7.1.2. Sensibilitatea termic general 7.1.2.1. Receptorii pentru cald (Ruffini) i pentru rece (Krause) sunt localizai n derm. Senzaiile de rece, cald sunt apreciate subiectiv n raport cu zeroul fiziologic (echivalent cu temperatura suprafeei tegumentelor, de la 20 - 36C. 7.1.2.2. Cile de transmitere sunt: - protoneuronul localizat n ganglionul spinal; - deutoneuronul localizat n cornul posterior medular, de unde
98 pleac fascicolul spino-talamic dorsal; - al III-lea neuron este localizat n talamus. Cile ascendente ale sensibilitii termice realizeaz conexiuni cortico-hi-potalamice, talamo-hipotalamice, conexiuni ce particip la homeostazia termic. 7.1.3. Sensibilitatea proprioceptiv general Analizatorul proprioceptiv sau kinestezic recepioneaz excitaiile legate de micrile i poziia diferitelor segmente ale corpului, transmise de la nivelul muchilor, tendoanelor, ligamentelor i suprafeelor articulare. 7.1.3.1. Receptorii sensibilitii proprioceptive sunt de 4 tipuri: - fusurile neuro-musculare, care recepioneaz ntinderea i modificrile de tensiune muscular; - organul tendinos Golgi localizat la jonciunea musculotendinoas recepioneaz tensiunea realizat n tendon n timpul contraciei musculare; - corpusculii Vater-Pacini sunt localizai n tendoane, articulaii, periost fascii musculare, au rol n recepionarea presiunii, elongaiei; - terminaiile nervoase libere din articulaii, tendoane, muchi particip i la transmiterea durerii profunde. Receptorii musculari asigur tonusul muscular bazai i postura incontient a membrelor, iar receptorii articulari asigur simul poziiei. 7.1.3.2. Cile de conducere ale sensibilitii proprioceptive contiente sunt asigurate de cile spino-bulbare Goll-Burdach. Din aceste ci se trimit colaterale la motoneuronii din coarnele posterioare ale mduvei (asigur arcuri reflexe senzitivo-motorii), la calea spino-cerebeloas i formaiunea reticulat (controleaz adaptarea automat). Sensibilitatea profund incontient ce recepioneaz i transmite informaii privind starea fusurilor neuro-musculare i a organelor tendinoase Golgi, prin calea spino-cerebeloas spre cerebel.
7.2. CAVITATEA BUCAL CA ZON REFLEXOGEN: REFLEXE MOTORII I SECRETORII
Cavitatea bucal reprezint regiunea organismului cea mai expus influenei mediului exterior. Receptorii existeni n cavitatea bucal transform stimulul recepionat n influx nervos, l codific i l transmit sistemului nervos. In cavitatea bucal se gsesc: interoceptori ce culeg informaii din vase, pro-prioceptori (receptori
99 musculari) ce asigur activitatea motorie a sistemului dento-maxilar (masticaia, deglutiia, vorbirea) i exteroceptori ce asigur sensibilitatea tactil, termic, dureroas, simul gustului. Receptorii tactili au densitatea cea mai mare la nivelul buzelor, vrfului limbii, gingii, mucoasa palatului dur, zone de sensibilitate maxim a organismului. Receptorii tactili au rol i de presoreceptori. Receptorii senzaiilor termice au densitate cea mai ridicat la nivelul buzelor i a limbii. Receptorii senzaiei dureroase ( algoreceptori) sunt teminaii nervoase libere amielinice sau mielinice la nivelul mucoasei, muchilor, articulaiei, periostului. Receptorii gustativi sunt localizai n principal pe suprafaa dorsal a limbii ca i n mucoasa jugal, epiglot, palatul dur, vlul palatin, culeg senzaii gustative, particip la procesele secretorii i motorii ale tubului digestiv. Dinii au o mare sensibilitate la nivelul dentinei, pulpei dentare, parodoniu. Cavitatea bucal constituie o zon reflexogen deosebit de important datorit bogiei i diversitii de receptori localizai aici. Reflexul reprezint transformarea unei stimulri senzitivo-senzoriale ntr-o reacie efectorie motoare sau secretoare. Reflexele pot fi: elementare (nnscute, au ci preformate) i condiionate (ctigate) se realizeaz prin legturi ntre centrii nervoi din scoara cerebral. Arcul reflex este suportul anatomic al actului reflex. Arcul reflex elementar, somatic sau vegetativ, este format din: - receptori, care pot fi de tip interoceptori, proprioceptori, exteroceptori; - ci senzitive, aferente, centripete; - centru nervos localizat n mduva spinrii sau trunchiul cerebral; - cale efectoare, centrifug, motoare sau secretoare, reprezentat de nervii corespunztori somatici sau vegetativi; - organ efector: muchiul striat (pentru reflexele somatice), muchiul neted, cardiac i esutul glandular (pentru reflexele vegetative). Clasificarea reflexelor: a) dup tipul de efector: - somatice - efectorul este musculatura striat; - vegetative - efectorii sunt glandele, muchii netezi ai vaselor i viscerelor; b) dup tipul de rspuns: - motor - reflexul masticator, deglutiie, voma, reflexul suptului, vorbirea; - secretor - secreia salivar, lacrimal, gastric, biliar, intestinal; c) dup funcie: - de nutriie: masticaie, supt, deglutiie, secretorii digestive; - de aprare - voma, tusea, lcrimare, strnut;
100 - de reglare a poziiei - reflexul de contracie tonic a muchilor masticatori, reflexul maseterin. Deci, cu punct de plecare din cavitatea bucal, se declaneaz numeroase reflexe cu importan att local ct i pentru organism n ntregime.
7.3. SENSIBILITATEA SISTEMULUI ORO-FACIAL
Sistemul oro-facial este dotat cu o mare sensibilitate la diferii stimuli din mediul extern, inclusiv stimuli nociceptivi, datorit inervaiei somato-senzitive deosebit de bogat i organelor senzoriale (gustativ, olfactiv, vizual, auditiv) ce sunt concentrate n regiunea cefalic. 7.3.1. Sensibilitatea tactil a sistemului oro-facial Simul tactil este considerat n primul rnd un sim cutanat dar include i receptorii tactili din cavitatea bucal, mai ales din vrful limbii, ca i mecano-receptori din ligamentul periodontal i articulaia temporo-mandibular. Receptorii din tegumente i mucoase sunt de tip mecanoreceptori stimulai de deformrile mecanice. Se descriu mai multe tipuri: - corpusculii Meissner localizai la nivelul buzelor i n vrful degetelor sunt sensibili la atingerile fine i se adapteaz rapid; - discurile Merkel se gsesc n pielea glabr, semnaliznd atingerea continu, puternic a obiectelor cu pielea; - terminaiile nervoase libere sunt rspndite pretutindeni i servesc la detectarea atingerilor uoare i a variaiilor de presiune; - corpusculii Ruffini i Pacini ce detecteaz presiunile mari adic deformrile mai pronunate ale tegumentelor sau a mucoaselor. Densitatea i felul receptorilor din diferitele arii tegumentomucoase condiioneaz sensibilitatea la stimulii mecanici a acestor zone. Pragul tactil reprezint presiunea minim necesar pentru a determina o senzaie tactil. Pragul tactil la nivelul nasului, buzelor, pulpa degetelor este de 2-3 g iar pentru regiunea spatelui este de 50 g. Pragul de discriminare reprezint distana minim ntre dou vrfuri de ac aplicate pe piele, la care sunt percepute ca stimuli separai. Pentru buze, pulpa degetelor acest prag este de 1,2-2 mm iar pentru spate este 60-70 mm. Receptorii din ligamentul periodontal sunt de tip mecanoreceptori, conin fibre nervoase care merg de la vrful rdcinii dintelui spre marginea gingiei. n ligamentul periodontal au fost descrise fibre
101 nervoase mielinice cu diametrul 10-12 micrometri implicate n senzaia tactil i fibre nervoase cu diametru mic, 6 micrometri, ce conduc sensibilitatea dureroas. Stimulul mecanic aplicat pe dinte determin deplasarea dintelui n alveol urmat de stimularea mecanoreceptorilor periodontali chiar la o deplasare de 2-3 micrometri. Mecanoreceptorii periodontali sunt implicai n: controlul prin feedback senzorial al micrilor de masticaie, n senzaia tactil i de presiune de la nivelul dinilor asigurnd aa numitul "sim ocluzal". La dinii normali, pragul tactil este pentru incisivi 1-0,44 g, pentru canini 1,32 g, pentru molari 6-7 g. La dinii depulpai pragul tactil a crescut cu 57%. Prin anestezia ligamentului periodontal pragul de discriminare este redus, dar nu abolit. De la receptorii tactili specializai impulsurile nervoase se transmit prin fibre senzitive de tip A beta cu vitez de conducere 30-70 m/sec. Terminaiile nervoase libere transmit impulsuri prin fibre amielinice de tip C, cu viteza de conducere 0,5-2 m/sec. 7.3.2. Sensibilitatea termic a sistemului oro-facial Sensibilitatea termic este rezultatul stimulrii receptorilor specifici pentru cald sau rece situai n derm sau mucoas. Clasic, receptorii pentru rece au fost considerai corpusculii Krause, iar pentru cald, receptorii Ruffini. Terminaiile receptorilor pentru rece sunt asociate cu fibrele senzitive A delta, iar pentru cald, sunt asociate fibrele C amielinice cu vitez mic de conducere.
7.3.3. Cile de conducere intranevraxiale pentru sensibilitatea somestezic a sistemului oro-facial Inervaia senzitiv a sistemului oro-facial este asigurat de nervii faciali (VII), glosofaringieni (IX), vagi (X) i trigemeni (V). Nervul facial culege sensibilitatea din regiunea feei, conductul auditiv extern, pavilionul urechii, regiunea retroauricular i din 2/3 anterioare a limbii. Primul neuron este n ganglionul geniculat. Nervul glosofaringian culege sensibilitatea din faringe, membrana timpanului, 1/3 posterioar a limbii, primul neuron se gsete n ganglionul Andersch. Nervul vag culege sensibilitatea din jurul conductului auditiv extern, regiunea retroauricular, mucoasa laringofarigian, primul neuron aflndu-se n ganglionul jugular. Protoneuronii acestor nervi fac sinaps cu deutoneuronul situat n nucleul solitar, iar de aici, dup ncruciare, fibrele ascendente (axonii) ajung n talamus (al IlI-
102 lea neuron) de unde sunt proiectate n regiunea inferioar a girusului postcentral. 7.3.4. Fiziologia trigemenului Nervul trigemen este a V-a pereche de nervi cranieni. Este un nerv mixt: senzitiv i motor. 7.3.4.1. Ramurile senzitive ale trigemenului Cele trei ramuri senzitive ale trigemenului sunt: nervul oftalmic, nervul maxilar superior i ramura senzitiv a nervului mandibular (Fig. 26). Nervul oftalmic culege stimulii din tegumentul frunii, pleoapa superioar, globul ocular, cornee, conjunctiv, plafonul cavitii nazale, piramida nazal. Nervul maxilar superior culege stimulii din regiunea infraorbitar, bolta palatin, cavitatea nazal, gingia superioar, dinii de pe arcada maxilar, buza superioar. Nervul mandibular senzitiv culege stimulii din regiunea temporal, paro-tid, maseterin superioar, buza inferioar, planeul bucal, gingie, parodoniu, dinii de pe mandibul. Prin poriunea motorie, nervul mandibular, culege sensibilitatea proprioceptiv a muchilor masticatori: maseter milohioidian, pntec anterior al digastricului, temporal, pterigoidian intern i extern. Trigemenul asigur prin fibrele sale senzitive inervaia tegumentelor feei i poriunii anterioare a capului, conjunctivei, mucoaselor nazale, bucale i sinusale, dinilor, precum i a unei suprafee intinse din duramater. Fibrele celor trei ramuri ptrund n cutia cranian, n ganglionul Gasser, unde se afl primul neuron. Nervul trigemen are trei nuclei senzitivi cu rol n inervaia senzitiv i reflexele monosinaptice ale muchilor masticatori. Cei trei nuclei senzitivi ai trigemenului sunt:
103
- nucleul mezencefalic cuprinde corpul neuronilor din desmodoniu care nu intra n ganglionul Gasser. Acesta prelucreaz i transmite excitaiile venite de la proprioceptorii din muchii temporali, maseter, pterigoidian, desmodoniu, creste alveolare, receptorii gingivali, parodontali ca i sensibilitatea profund i durerea muscular; - nucleul senzitiv principal al trigemenului este aezat n punte, primete excitaiile exteroceptive tactile din regiunea corespunztoare celor trei ramuri ale trigemenului, este nucleul integrator al masticaiei; - nucleul spinal al trigemenului prelucreaz i transmite excitaii termice i dureroase din ntregul tract trigeminal. n ganglionul Gasser se gsete protoneuronul sensibilitii tactile, de aici, axonii si fac sinaps n nucleul senzitiv principal al trigemenului cu deutoneuro-nul, de unde, axonii acestora intr n lemniscul trigeminal (tract trigeminotala-mic) i ajung n nucleul ventro-postero-median al talamusului (III neuron), de unde se proiecteaz n circumvoluia parietal post central S1 (fig. 27).
104
Din ganglionul Gasser pleac fibre descendente ce formeaz tractul bulbospinal al trigemenului, ce conduce sensibilitatea termic, dureroas i fac sinaps cu deutoneuronul n nucleul spinal al trigemenului. De aici, axonii se ncrucieaz pe linia median, se altur lemniscului trigeminal (tract trigeminotalamic) i conduc sensibilitatea termic, tactil brut spre nucleul ventro-posteromedian al talamusului (al III-lea neuron). Din nucleul spinal al trigemenului se trimit fibre i spre neuroni din substana reti-culat, formndu-se calea trigemino-reticulo-talamic, ce conduce sensibilitatea dureroas spre nucleul ventro-postero-median al talamusului (al III-lea neuron) de unde se proiecteaz n girusul postcentral, partea inferioar. Protoneuronul, din nucleul mezencefalic al trigemenului, ce primete fibre senzitive de la proprioreceptorii muchilor masticatori, mecanoreceptorii perio-dontali i gingivali, trimite axonii n tractul trigeminotalamic (tract cvinto-talamic) ce ajung n talamus. Din traiectul ascendent al cilor senzitive se desprind colaterale:
105 fibre pontopontine, fibre proprioceptive trigeminale pentru cerebel i pentru cile descendente, cu care nchid reflexe monosinaptice sau polisinaptice. 7.3.4.2. Ramurile motorii ale trigemenului Nucleul motor al trigemenului este localizat n poriunea superioar a punii. Rdcina motorie a trigemenului este alctuit din prelungirile neuronilor localizai n nucleul motor. Rdcina motorie inerveaz: muchii ridictori ai mandibulei (maseter, temporal, pterigoidian intern, extern, milohioidian), perista-filinul extern (muchiul dilatator al trompei Eustachio), muchiul ciocanului (muchi tensor al timpanului). Nucleul motor al trigemenului primete aferente de la: fusurile proprioceptive musculare, nucleul senzorial mezencefalic al trigemenului (asigurnd reflexul miotatic de ridicare a mandibulei) i de la cortexul motor prin cile cortico-pontine (asigurnd reflexul masticator coordonat, finalizat, care determin realizarea bolului alimentar). Trigemenul are i o funcie neurovegetativ reprezentat de funcia secretorie, vasomotorie, sudoripar i trofic. Funcia secretorie este realizat de fibrele secretorii de natur parasim-patic ce traverseaz trigemenul i care vin n realitate de la nuclei ce aparin facialului i glosofaringianului (nucleul lacrimonazal, nucleul salivator superior, nucleul salivator inferior). Funcia vasomotorie se datorete fibrelor de natur simpatic vasomotorie ce parcurg ramurile trigemenului. Dup seciunea trigemenului reaciile vasorno-torii sunt intense i de durat. Funcia sudoral se realizeaz cu participarea fibrelor de natur simpatica ce strbat ramurile trigemenului. Excitarea unei ramuri cutanate, nazale sau gustative duce la hiperhidroz facial. Funcia trofic cuprinde toate esuturile feei i este dependent de ganglionul Gasser. Lezarea trigemenului poate determina pierderea precoce a dinilor, descuamarea corneei sau chiar ulceraie. Trigemenul particip la numeroase reflexe alimentare i de aprare: deglu-titie, reflexul masticator, reflexul maseterin-miotatic, reflexul suptului, reflexul de strnut, reflexul oculo-cardiac, reflexul lacrimal, reflexul corneean de clipire. Deglutiia este posibil numai dac are loc nchiderea gurii prin contracia maseterului. Reflexul maseterin: se produce o contracie a maseterului la percuia arcadei dentare inferioare, bolnavul avnd gura ntredeschis. Reflexul miotatic: ntinderea maseterului prin coborrea mandibulei, determin stimularea proprioceptorilor musculari, ceea
106 ce induce reflex, contracia maseterului urmat de nchiderea gurii. Reflexul suptului: la sugar, atingerea buzelor sau a altor zone din apropiere declaneaz suptul. Reflexul dispare n jurul vrstei de un an. Reflexul de strnut: este un reflex de aprare, declanat de factori iritani ai mucoasei nazale, inervat de trigemen. Strnutul ncepe printr-o inspiraie profund urmat de contracia puternic a muchilor expiratori cu glota nchis, ceea ce determin creterea presiunii intratoracice urmat de deschiderea brusc a glotei i epiglotei. Are loc expulzarea exploziv a aerului ce antreneaz corpul stin sau substana iritant din nas. Reflexul oculo-cardiac: compresiunea ferm a globilor oculari, cu pleoapele nchise, induce prin aferena trigeminal activarea centrului bulbar depresor cardiac. Acesta, pe cale eferent vagal, determin: scderea frecvenei cardiace, scderea forei de contracie miocardic, vasodilataie, hipotensiune arterial. Reflexul lacrimal: este declanat de iritaiile corneei, conjunctivei i mucoasei nazale, zone inervate de trigemen, dar poate fi iniiat i cortical (plnsul). Reflexul corneean de clipire: atingerea corneei cu un tampon mic de vat determin clipirea. Reflexul corneean de clipire este diminuat sau abolit bilateral n com (n funcie de profunzimea comei) i n anestezie general. Leziunea unilateral a rdcinii motorii a trigemenului determin devierea spre partea bolnav a ovalului gurii. Leziunea bilateral a rdcinilor motorii a trigemenului determin coborrea permanent a mandibulei cu imposibilitatea de masticaie.
7.4. SENSIBILITATEA DUREROAS GENERAL I OROFACIAL
Durerea, constituie un sistem de alarm, care semnalizeaz aciunea unor ageni nocivi asupra organismului, ndeplinind astfel o funcie de salvare a vieii. Durerea se manifest sub anumite forme: neptur, arsur, durere surd, nsoite ntotdeauna de o reacie afectiv. n patologia oro-facial durerea ocup un rol principal i este nsoit adeseori de reacii afective importante din partea individului. 7.4.1. Receptorii nociceptivi Acetia sunt terminaii nervoase libere ale fibrelor mielinice subiri A delta, cu vitez medie de conducere 15 m/s i ale fibrelor amielinice de tip C cu vitez de conducere 0,5 m/s. Receptorii
107 nociceptivi, n funcie de stimul, pot fi: termoalgoceptori, mecanoalgoceptori, chemoalgoceptori. Nociceptorii au posibilitatea de a rspunde la mai multe categorii de stimuli, nu se adapteaz, sunt rspndii n toate esuturile, dar au densitate mai mare n stratul superficial al pielii, periost, pereii arteriali, suprafeele articulare. 7.4.2. Durerea n regiunea-orofacial Durerea dentar Inervaia dinilor este asigurat de nervii dentali (ramuri din trigemen) ce ptrund prin orificiul apexian, urc n pulpa dentar, se rspndesc ctre bolta i pereii camerei pulpare, formnd un plex de fibre mielinice i amielinice sub-odontoblastic (plex Raschkow). La acest nivel s-au pus n eviden i fibre vegetative ce sunt asociate vaselor de snge pulpare. Se accept, n general, c fibrele nervoase ptrund n stratul intern al dentinei avnd funcie de receptori, asemntor terminaiilor nervoase libere din tegument. Terminaii nervoase libere trigeminale s-au gsit n dentina circumpulpar coronar, stratul odontoblastic, plexul Raschkow. n schimb dentina radicular, jonciunea amelo-dentinar nu au inervaie. Inervaia dentinar senzitiv apare odat cu erupia dinilor, densitatea ei crescnd pn la maturizare, paralel cu sensibilitatea dureroas la stimuli termici i electrici. Receptorii nociceptivi intradentinali se pare c sunt constituii de ctre odontoblaste a cror prelungiri intr n dentin, ntre odontoblaste i terminaiile nervoase senzitive existnd jonciuni de tipul sinapselor. S-a emis i ipoteza hidrodinamic, dup care receptorii din pulp ar fi stimulai de perturbrile mecanice consecutive deplasrii coninutului tubilor dentinali. Stimulii nociceptivi ar induce deplasarea nucleelor odontoblastelor n tubii dentinali, deplasarea de lichide prin dentin, care ar determina modificri de presiune la nivelul pulpei, bogat inervate. Din punct de vedere practic ns se asociaz durerea dentinar cu durerea pulpar, deoarece esutul pulpar este aproape ntotdeuna implicat n suferina dentinei. Durerea de origine vascular poate rezulta din leziuni ale vaselor sau esutului perivascular. Vasele sanguine au chemoreceptori, sensibili la substane productoare de durere: bradikinina, histamina, serotonina. Inflamaia pulpar se nsoete de eliberarea de substane algogene, care stimuleaz chemoreceptorii vasculari pulpari genernd durere. Durerea muscular este dat de spasmul, ischemia, inflamaia, rupturi ale muchilor masticatori, ale periostului, articulaiei temporomandibulare, ce determin durerea facial. Mecanismul de producere are la baz: scderea O2, pH acid, acumularea de acid lactic, histamina, care induc durerea muscular.
108 Durerea de origine salivar - se produce datorit prezenei sialoliilor n glandele salivare. Durerea apare n timpul alimentaiei cnd saliva secretat produce distensia sistemului canalicular, evacuarea ei fiind diminuat prin obstrucia realizat de sialolii. Durerea raportat (iradiat) este durerea resimit simultan n alt loc dect n cel afectat. Convergena la nivelul trunchiului cerebral a fibrelor periferice pentru sensibilitatea dureroas oro-facial ca i spasmul vascular reflex, pot explica iradierea durerii. Durerea este cel mai frecvent simptom pentru care pacientul se adreseaz medicului, durere asociat cu reacii emoionale puternice. Cauzele cele mai frecvente ale durerii n regiunea maxilarelor i a feei sunt pulpitele i periodontita periapical secundare cariei dentare.
PRINCIPALELE CAUZE DE DURERE N REGIUNEA OROFACIAL (dup CAWSON, 1991) 1. AFECIUNI ALE GURII I MAXILARELOR a) Boli ale dinilor i esuturilor de susinere: - Pulpite secundare: cariilor, fracturilor de coroan, leziunilor termice din timpul manevrelor stomatologice - Periodontita periapical - Abcese laterale (periodontale) - Gingivita ulcerativ acut b) Boli ale mucoasei orale: - Afte recurente - Lichen plan eroziv - Herpes simplex - Herpes Zoster - Tuberculoz - Carcinom i alte neoplasme c) Boli ale maxilarelor: - Fracturi - Osteomielit - Tumori maligne d) Durerea la pacientul edentat: - Traumatisme ale protezei - Boli ale mucoasei bucale - Boli ale maxilarelor e) Durerea postoperatorie: - Osteit alveolar - Fractura maxilarului - Osteomielit
109 - Leziuni ale trunchiului nervos sau afectarea nervilor n esutul cicatricial f) Durerea masticatorie: - Boli ale articulaiei temporomandibulare - Arterit temporal - Nevralgia trigeminal - Boli ale dinilor i esuturilor de susinere - Calculi salivari (durerea se declaneaz la nceputul alimentaiei) 2. BOLI EXTRAORALE a) Boli ale sinusurilor: - Sinusite acute - Carcinoame b) Boli ale glandelor salivare: - Parotidita acut - oreion - Calculi salivari - Sindrom Sjogren - Neoplasme c) Boli ale urechilor: - Otit medie - Neoplasme d) Boli ale ochilor: - Glaucom, foarte rar poate cauza durere n maxilarul superior e) Durerea de origine vascular: - Migrena - Arterita temporal - Infarct miocardic f) Dureri ale sistemului nervos: - Nevralgia trigeminal - Tumori intracraniene - Nevralgia postherpetic - Zona Zoster - Scleroza multipl g) Durerea facial psihogenic. Durerea i anxietatea sunt totdeauna asociate. Durerea cauzeaz anxietate, iar anxietatea crete sensibilitatea la durere i ocazional poate crea sindromul dureros. Controlul durerii, la pacieni cu grad mare de nervozitate este dificil. Cnd anxietatea devine insuportabil, este necesar sedarea pacientului prin medicamente anxiolitice. 7.4.3. Cile de conducere al sensibilitii dureroase generale Nociceptorii sunt n legtur cu primul neuron, localizat n ganglionul spinal, care face sinaps cu neuronii intercalri din
110 substana gelatinoas' Rolando, care sunt interpui ntre protoneuron i deutoneuron. Al doilea neuron (deutoneuronul) este localizat n cornul posterior medular unde i are originea tractul spinotalamic. De la deutoneuron se trimit conexiuni la un neuron vegetativ din cornul lateral medular, astfel c un impuls dureros declaneaz n acelai timp i un reflex vegetativ. Din tractul spinotalamic se descriu dou ci: - tractul neo-spinotalamic (lemniscal), specific, care se termin n nucleul ventro-postero-median. Este oligosinaptic, conduce rapid, are o somatotopie foarte precis, este corelat cu percepia senzaiilor dureroase precis localizate; - tractul paleo-spinotalamic (extralemniscal, subcortical al durerii) este multisinaptic, nespecific, conduce ncet, nu are o somatotopie precis, se termin n nucleii talamici. Din fascicolul spinotalamic se desprind colaterale, ce realizeaz conexiuni cu substana reticulat a trunchiului cerebral, legturi responsabile de manifestrile respiratorii i circulatorii din cursul durerii. Cile de conducere al sensibilitii dureroase oro-faciale Ramurile senzitive ale nervului trigemen culeg informaiile dureroase de la nociceptori i le trimit n ganglionul Gasser (protoneuronul). De aici, un grup de fibre descendente merg la nucleul spinal al trigemenului unde fac sinaps cu deutoneuronul, de unde o parte din fibre se ncrucieaz pe linia median, se altur lemniscului trigeminal (tract trigemino-talamic, sau cvinto-talamic) i conduc sensibilitatea tactil brut, termic, iar alte fibre se ndreapt spre substana reticulat formnd cale trigemino-reticulo-talamic, ce transmite sensibilitatea dureroas. Deci, din nucleul spinal al trigemenului, informaiile de la nociceptori se transmit, fie prin lemniscul trigeminal, fie pe calea trigemino-reticulo-talamic, difuz n talamus i de aici se proiecteaz difuz pe scoar, la nivelul ariilor asociative, integrative ale rinencefalului, hipotalamusului i nucleii cenuii. Prin excitarea i interaciunea complex a unor arii corticale variate se realizeaz comportamentul afectiv, emoional i cognitiv al durerii. Modularea durerii Controlul percepiei dureroase se realizeaz prin stimuli afereni i prin mediatori. Controlul durerii prin stimuli afereni: - la nivelul medular: activitatea fibrelor A alfa groase blocheaz transmiterea impulsurilor nociceptive vehiculate de fibrele A delta i C. Aceasta se explic prin teoria "controlului de poart" care se bazeaz pe fenomenul de inhibiie presinaptic. Fibrele A alfa excit
111 interneuronul inhibitor din substana gelatinoas Rolando, iar aceasta i exercit efectul inhibitor asupra deutoneuro-nului n "T" din cornul posterior medular, blocnd astfel transmiterea impulsurilor nociceptive ("nchiderea porii"). Fibrele C subiri i A delta inhib interneuronul din substana gelatinoas, determinnd fenomene excitatorii asupra deutoneuronului, favoriznd astfel transmiterea impulsurilor nociceptive (tendina de "deschidere a porii"); - substana reticulat a trunchiului cerebral are rol supresor asupra mesajelor ascendente spinale nociceptive; - nucleul talamic ventro-postero-median filtreaz i controleaz aferentele nociceptive; - la nivelul cortical aferentele dureroase se proiecteaz pe lobul temporal (circumvoluia retrorolandic), de aici calea de conducere se continu pe lobii frontali unde se realizeaz percepia dureroas complex integrat n personalitatea subiectului. Aceast integrare manifestat prin capacitatea de evaluare a senzaiilor dureroase i reacia motivaional afectiv se realizeaz prin conexiunile cortexului cu rinencefalul, hipotalamusul i substana reticulat a trunchiului cerebral. Controlul durerii prin mediatori (neurochimia durerii): Mediatorii chimici ai durerii sunt substane apte de a stimula receptorii algogeni periferici, de a transmite excitaiile la cile durerii, de a modula transmiterea excitaiilor dureroase, mai ales n sens inhibitor. Substane inhibitoare ale transmiterii durerii sunt n principal opiaii endogeni: enkefalinele (acioneaz la nivel medular) i endorfmele (acioneaz la nivel diencefalo-mezencefalic). Exist trei familii de opiai endogeni: enkefaline, endorfine i dinorfine. Enkefalinele se gsesc n medulosuprarenal, n sistemul nervos central (avnd rol de transmitor sinaptic), la terminaiile nervoase din tractul gastro-intestinal (scad motilitatea intestinal) i n substana gelatinoas (avnd rol analgezic). Enkefalinele sunt polipeptide, una conine metionin (metenkefalina) i alta izoleucin (leu-enkefalina). Sinteza enkefalinelor pornete de la un precursor i anume molecula de proenkefalin din care deriv: - 4 molecule de met-enkefalin i cte o molecul de leu-enkefalin, octapeptid, heptapeptid. Enkefalinele acioneaz pe receptori opioizi de tip: 5 (delta), cuplai cu proteina G0 inducnd nchiderea canalelor de calciu. Metabolizarea enkefalinelor se realizeaz de ctre peptidazele: enkefa-linaza A, B, aminopeptidaza. Principalul efect al enkefalinelor este analgezia (vezi tabelul 9), prin aciune la dou nivele: spinal i supraspinal. La nivelul coarnelor dorsale ale mduvei spinrii interneuronii enkefalinergici, prin
112 inhibiie presinaptic, scad eliberarea substanei P (mediatorul chimic al durerii) de la nivelul fibrelor nociceptive A8 i C. Supraspinal, neuronii enkefalinergici activeaz sistemul descendent inhibitor al recepiei nociceptive, prin aciune asupra substanei cenuii periapeductale i a nucleului rafeului bulbar. Sistemul descendent inhibitor, prin calea final comun serotoninergic, inhib deutoneuronii cilor nociceptive din nucleii senzitivi ai trigemenului i din coarnele dorsale medulare. Endorfinele tip (beta) sunt polipeptide cu 31 de aminoacizi care au ca precursor pro-opiomelanocortina ce se gsete n lobul anterior i intermediar al hipofizei i n creier. Endorfinele se gsesc, n special, n nucleul arcuat i n nucleii periven-triculari (sistemul neuronilor secretori de P-endorfine) i n adenohipofiz (de unde se elibereaz n snge printr-un mecanism calciu-dependent). Endorfinele acioneaz pe receptori specifici membranari (rniu), cuplai cu proteina G. Activarea acestor receptori determin: creterea conductanei membranei pentru potasiu, hiperpolarizarea neuronilor centrali i aferentelor primare, scderea excitabilitii. Endorfinele (gama) au efect neuroleptic (diminua activitatea spontan, hiperreactivitatea provocat i agresivitatea), prin inhibiie presinaptic blocant a eliberrii de dopamin. Endorfmele a au efect psihostimulant. Dinorfinele au ca precursor molecula de pro-dinorfin. Se gsesc n duoden (dinorfina 1-17), n neurohipofiz i hipotalamus (dinorfina 1-8). Dinorfinele acioneaz pe receptori membranari K (kappa), inducnd nchiderea canalelor de calciu. Ca i alte peptide opioide, dinorfinele au efect analgezic. Tabelul 9. Receptor Ligand endorfine Efecte analgezie depresie respiratorie constipaie euforie dependen mioz
113 K dinorfine analgezie diurez sedare mioz disforie
enkefaline
analgezie
Acupunctura are efect analgezic prin stimularea producerii de opiai endogeni i prin eliberarea de endorfine hipofizare. "Analgezia de stress" se produce datorit eliberrii de endorfine hipofizare i enkefaline medulosuprarenale. La nivelul tractului digestiv, opiaii endogeni au efecte miorelaxante i antisecretoare, prin scderea eliberrii de acetilcolin din plexurile intramurale. Serotonina, prin intermediul formaiei reticulate (unde se realizeaz sinapse serotoninergice), blocheaz transmiterea ascendent a mesajelor nociceptive. Substane productoare de durere i care faciliteaz transmisia ei sunt: bradikinina, histamina, serotonina, ionii de potasiu i calciu, hidrogen ionii (cnd pH scade sub 6,2, durerea maxim este la pH 3,2), ATP, substana P, prosta-glandinele. Hipoxia local determin acumularea de hidrogeni ioni, acumularea de substane vasoneuroreactive. Acidoza local activeaz prekalicreina care se transform n kalicrein, ce acioneaz asupra kininogenului, rezultnd diverse kinine cu rol vasodilatator i care stimuleaz nociceptorii (Fig. 28).
114
Substana P este un peptid endogen, mediator, ce stimuleaz neuronii n "T" din corpul posterior medular activnd deci transmisia ascendent a mesajelor dureroase. Fibrele amielinice senzitive din pulpa dentar conin substane P. Prostaglandinele (PG) reprezint un grup de metabolii oxidai ai unor acizi grai eseniali, cu 20 atomi de carbon, numii eicosanoizi (Fig. 29). Prostaglandinele poteneaz efectul substanelor productoare de durere i anume efectul bradikininei. De exemplu, PGE2, prostaciclina stimuleaz formarea bradikininei care are efect excitator la nivelul nociceptorilor. Aspirina, Indometacinul, inhib ciclooxigenazele deci sinteza de prostaglandine i astfel au efect analgezic.
115
Nevralgia nervului trigemen Nevralgia trigeminal este dominat de simptomul durere, durere care este una din cele mai cumplite din patologia uman. Nevralgia trigeminal se caracterizeaz prin dureri violente, paroxistice, care survin n crize i cuprind una sau dou ramuri ale trigemenului. Clinic, se descriu trei tipuri de nevralgie: - nevralgie trigeminal primitiv; - nevralgie trigeminal secundar; - sindromul trigeminal.
116 Nevralgia trigeminal primitiv (NTP) Nevralgia trigeminal primitiv este descris nc din secolul I .e.n. i este cunoscut n literatur sub denumiri ca: nevralgie trigeminal paroxistic, tic dureros al feei, nevralgie esenial, nevralgie idiopatic, etc. NTP apare mai frecvent ntre 40-60 ani, afectnd ambele sexe. Caracteristicile clinice ale NTP: Caracteristicile clinice ale nevralgiei primitive sau eseniale de trigemen se pot sistematiza astfel: - durerea este limitat ntotdeauna la teritoriul ramurilor trigemenului. n mod cu totul excepional durerile pot iradia spre alte regiuni ale corpului cum sunt regiunile cervical, occipital sau auricular. - ramurile trigeminale sunt afectate ca frecven n urmtoarea ordine: ramura a III-a, ramura a II-a i ramura I. Afectarea concomitent a celor trei ramuri ale trigemenului este foarte rar. Durerea poate fi localizat strict la una dintre ramurile trigemenului, dar n acelai timp poate iradia pe una din ramurile vecine sau se poate generaliza. Durerea trigeminal bilateral este cu totul excepional i atunci cnd exist ea poate fi simultan sau alternativ. - durerea din nevralgia trigeminal esenial apare n crize.Crizele ncep brusc i dispar brusc, fiind urmate de perioade de linite total. - durerea este descris n mod diferit de bolnavi fiind comparat cu o neptur de cuit, cu o fulgertur, constricie, apsare, arsur, etc. - durerea este ntotdeauna declanat de factori mecanici (vorbire, masticaie, atingere, splat, brbierit, suflatul nasului ).Examenul stomatologic poate declana de asemenea criza dureroas. - durerea are ntotdeauna acelai punct de plecare periferic denumit zon trgaci sau "trigger zone" a lui PATRICK. n practic este adesea greu de identificat zona trgaci, bolnavul relatnd confuz i inconstant declanarea durerii. Extrem de rar "trigger zone" poate fi localizat extratrigeminal: pe rdcinile C2-C3 respectiv nervii auricular mare i marele occipital n haluce, n axil. - criza dureroas din nevralgia de trigemen este constant nsoit de fenomene motorii i vegetative. n timpul crizei se produce reflex o contracie generalizat a muchilor mimicii de partea hemifeei bolnave, un hemispasm facial sau "grimasa facial" caracteristic, n timpul creia asistm la o contracie brusc i repetat a muchilor pleoapelor, muchilor penorali, a muchilor pieloi fr ns contracia frontalului. Dup afirmaia unor bolnavi, grimasa facial ar fi un mod de
117 reacie antalgic. Dintre tulburrile simpatice, se constat manifestri vaso-motorii (roeaa feei) i secretorii (hipersecreie lacrimal, sudoripar i chiar salivar). Aceste tuiburri rmn pe plan secundar fa de accesul dureros. - durerea din nevralgia trigeminal nu cedeaz la antalgicele obinuite. - crizele dureroase nu apar n timpul nopii i atunci cnd apar ele suni provocate de atingeri involuntare sau alte acte mecanice necontrolate, n timpul somnului. - durerile au un caracter recidivant, ele survin ciclic dup perioade de linite de sptmni, luni sau ani de zile, avnd acelai caractere cu crizele precedente, putnd varia doar intensitatea i durata crizelor. Se poate observa c c frecven mai mare a cazurilor se nregistreaz n clinic primvara i toamna ceea ce ar putea sugera influena factorilor climatici i atmosferici n declanare; i accentuarea episoadelor dureroase. - dup perioade lungi de suferin sau de rspunsuri terapeutice nesatis fctoare unii dintre bolnavi pot prezenta tulburri psihice de diferite grade prezentnd un facies i avnd un comportament care trdeaz o mare suferin. Teorii patogenice ale NTP: Nevralgia trigeminal are o patogenie complex i prezint unele caracte ristici care o difereniaz de alte tipuri de dureri nevralgice. n majoritatea cazurilor stimulii sunt de natur mecanic, un rol deoseb: avnd receptorii tactili i proprioceptivi. De obicei ,n toate situaiile exist o zon declanatoare n teritoriul crei acioneaz stimulii. PATRICK, n 1914, a denumit aceast zon declanatoar zon "dolorigen" sau "trigger area" n prezent "trigger zone". Nevralgia trigeminal este caracterizat de o perioad refractar cai urmeaz fiecrei crize, perioad n care durerea nu mai poate fi provocat de ni< un stimul adecvat sau preferenial. Lungimea acestei perioade este direct proporional cu intensitatea durata crizei precedente. Nevralgia esenial a nervului trigemen ar putea fi produs de ctre con presiuni la nivelul unghiului ponto-cerebelos, ceea ce justific susinerea teori mecanice n patogenia ei. De asemenea, s-a discutat influena factorilor mecanici de tip iritativ nivelul rdcinii senzitive a nervului trigemen Printre aceti factori sunt anevrismele, angioamele, contactul dintre rdcina senzitiv i artera sau vena petroas. O alt teorie patogenic const n susinerea existenei ntinderii trige-minale datorit coborrii fosei cerebrale posterioare. Acest proces s-ar produce n mod fiziologic, cnd datorit vrstei discurile
118 vertebrale cervicale se reduc prin atrofie senil, scurtnd coloana cervical i cobornd coninutul fosei cerebrale posterioare. Situaia aceasta determin o angulaie mai accentuat a nervului trigemen la nivelul stncii osului temporal. mpotriva acestei teorii vine, ca argument, faptul c dac ntradevr aa s-ar ntmpla, ar nsemna ca marea majoritate a vrstnicilor s sufere de nevralgie de trigemen. S-a emis teoria c angulaia nervului poate s apar i datorit nclinaiei anormale a stncii, datorit unui proces de demineralizare. Nevralgia trigeminal secundar (NTS) Nevralgia trigeminal secundar, se refer la toate cazurile care, dei reproduc fidel caracterele durerii din nevralgia trigeminal prezint totui foarte discrete semne de suferin organic a nervului (discrete zone de hipoestezie, diminuarea reflexului corneean, etc), precum i n acele cazuri n care dei exist tabloul clinic al nevralgiei trigeminale, la interveniile chirurgicale pentru suprimarea durerii se gsete un factor de iritaie trigeminal (bride leptomeningeale, anse vasculare, formaiuni chistice sau tumori extratrigeminale). Nevralgia trigeminal secundar este caracterizat prin simptomul durere care se datorete unor stri patologice n teritoriile de receptivitate ale trigeme-nului, fiind tipice pentru o serie de afeciuni dento-parodontale, mucoase, osose, sinusale, tumorale, traumatice, etc. Durerea se prezint particular de la individ la individ n funcie de reactivitatea acestora. Examenele clinice i paraclinice pot pune n eviden: pulpite, parodontite, incluzii dentare, erupii dentare dificile, alveolite postextracionale, infecii nazale, infecii sinusale, osteite, osteomielite, litiaz salivar, artrite temporo-mandibulare i altele. Cauza nevralgiei, de obicei, este decelabil i tratarea ei suprim fenomenul dureros. Din punct de vedere clinic, durerile trigeminale se pot apropia, n situaii excepionale, de caracterul celor din NTP. Cel mai frecvent, durerile sunt asociate cu semne de suferin organic a nervului trigemen, implicnd funcia senzitiv, reflex, motorie i vegetativ. Factorii etiologici ai NTS: a. Factori tumorali: - schwanoamele sau neurofibroamele ganglionului GASSER sau ale rdcinii trigeminale retrogasseriene; - tumorile paratrigeminale (meningioame, colesteatoame), care sunt rare; - tumorile maligne de vecintate care invadeaz secundar trigemenul la nivel extranevraxial i care determin predominent
119 semne organice de suferin a trigemenului, deci un sindrom trigeminal secundar. b. Factori inflamatori: - herpesul este cel mai frecvent dintre aceti factori i afecteaz n procent de 95% din cazuri ramurile oftalmice. Durerile preced de regul apariia veziculelor herpetice, n unele cazuri durerile survenind tardiv dup apariia veziculelor. Dac sunt afectai nervii lacrimal i nazociliar pot s apar cheratite i ulcere corneene. Durerile pot fi declanate de stimuli exteroceptivi sau stresuri emoionale fr a exista ns un "trigger zone" caracteristic. Sediul procesului virotic se afl, n general, n ganglionul lui GASSER. - inflamaiile periferice pe traiectul ramurilor trigeminale (afeciuni ale pulpei dentare, sinusite, etc.) nu reprezint factori etiologici ci doar factori favorizani. - arahnoiditele de fos cerebral posterioar -reprezint un factor etiologic cert dei este foarte rar. - scleroza n plci: n general, durerile trigeminale apar tardiv n evoluia leuconevraxitei. c. Ali factori etiologici: - traumatismele cranio-cerebrale cu fracturi la nivelul apexului stncii temporalului; crizele malarice; reumatismul; guta; diabetul; luesul. Sindromul dureros trigeminal secundar Sindromul dureros trigeminal secundar definete acele cazuri n care durerea nu este identic celei din nevralgia trigeminal i nu este pe primul plan al simptomatologiei. Sindromul trigeminal este dominat de o suferin organic a nervului trigemen i a structurilor paratrigeminale, aa cum se ntmpl n tumorile de baz de craniu.
ROLUL RECEPTORILOR BUCALI N MENINEREA HOMEOSTAZIEI Homeostazia reprezint meninerea constant a structurii organismului, a acelor valori chimice, umorale, etc, ce sunt indispensabile pentru asigurarea funciilor normale ale individului. Homeostazia nu se refer numai la mediul intern, ci la toate procesele fiziologice. Ca atare, putem vorbi de multiple aspecte ale homeostaziei, ncepnd de la celul i pn la comportamentul ansamblului organismului. La nivelul cavitii bucale au fost evideniate influene generale, metabolice, nervoase, din organism. Dar i procesele metabolice i nervoase ce se desfoar n structurile acestei caviti, pot determina modificri reflexe n diverse esuturi i organe. Deci,
120 putem vorbi de participarea receptorilor bucali, inclusiv a celor la nivelul aparatului dento-maxilar la homeostazia organismului, ct i de o homeostazie ntre componentele cavitii bucale, respectiv ale aparatului dento-maxilar. Homeostazia nu nseamn lipsa de modificare, ci un joc continuu ntre anumite limite cantitative impuse de dinamica echilibrului biologic. Ea are la baz procesul fiziologic de retroaferentare-conexiune invers (feed-back). Prin conexiune invers se asigur o interaciune ntre efect i cauz, corectarea fcndu-se totdeauna de la efect la cauz. Mecanismele de feed-back negativ determin readucerea parametrului deviat, napoi, ntre limitele valorilor ideale, standard. Ele reprezint mecanismele fiziologice de reglare ale tuturor parametrilor funcionali ai organismului. n schimb, mecanismele de feed-back pozitiv sunt declanate de ctre devieri mari ale parametrilor i au ca rezultat accentuarea, agravarea acestor dezechilibre ce se finalizeaz n starea de boal. Cavitatea bucal particip la meninerea homeostaziei generale prin: - eliminarea i la nivelul cavitii bucale a substanelor nocive (corpi cetonici, uree, metale grele), n cazul intoxicaiilor endogene, eliminarea de medicamente, microorganisme; - uscarea regiunii orofaringiene determin senzaia de sete, creterea aportului hidric i restabilirea echilibrului hidric; - meninerea troficitaii mucoasei bucale contribuie att la protecia esuturilor subiacente fa de agenii mecanici, termici, chimici, infecioi, din mediul bucal sau extern, ct i la desfurarea normal a funciilor la acest nivel; - stimularea secreiei salivare, n afara aportului alimentar, asigur meninerea umiditii mucoasei n vederea articulrii cuvintelor n timpul vorbirii. Adaptarea secreiei salivare, cantitativ i calitativ (pH, mucin, componeni cu rol antiseptic) n funcie de natura alimentului, contribuie la realizarea homeostaziei funciei de digestie; - n cursul alimentrii, stimulii compleci solicit mai muli receptori: olfactivi, tactili, termici, gustativi, musculari, care au rol de aprare, sesiznd substanele alterate i mpiedicnd folosirea lor. Receptorii gustativi prin aezarea la nivelul cavitii bucale declaneaz n mod reflex secreiile digestive; - masticaia, secreia salivar, gustaia, deglutiia sunt acte care se desfoar prin intermediul receptorilor bucali, acte care particip la homeostazia funciei digestive, iar aceasta la homeostazia ntregului corp; - fizionomia feei - mimica nu are aspect static ci variaz prin aciunea muchilor subiaceni. Aspectul feei se modific n permanen pentru a exprima veselie, tristee, durere. Prin rolul funcional de exteriorizare, de comunicare, a strilor afective, fizionomia particip att la homeostazia sistemului dento-maxilar ct i la homeostazia ntregului organism.
121 De exemplu: n timpul actului motor al secionrii unui aliment, introdus ntre cele dou arcade, prin comenzi corticale se asigur precizia micrilor mandibulei. n cursul micrilor de secionare, ca rezultat al contraciei musculare, scoara este informat de eficiena contraciilor, iar dac presiunea de secionare este prea mic, se comand intervenia i a altor uniti motorii astfel ca secionarea s reueasc. Fonaia, vorbirea articulat, prin intermediul crei omul i exprim gndurile, reprezint mijlocul de comunicare ntre membrii societii i determin homeostazia social.
PROIECIA CORTICAL A SENSIBILITII SOMESTEZICE ORO-FACIALE I GENERALE Sensibilitatea oro-facial se proiecteaz n poriunea inferioar a lobului parietal i pe marginea superioar a santului lateral. Lobul parietal este preponderent senzorial i de asociaie. Cuprinde ariile 3,1,2 i 43,5 i 7,39 i 40. Scoara somestezic primar este format din ariile 3,1,2. Are o somato-topie precis (homunculus senzitiv), predominant contralateral, diversele regiuni sunt integral reprezentate pe scoar, cea mai mare reprezentare avnd buzele i limba, urmate de mn i police (fig. 30).
122 Mrimea ariilor corticale este direct proporional cu numrul i varietatea receptorilor senzitivi din fiecare arie periferic a corpului. Reprezentarea eorti-cal a percepiei senzitivo-senzoriale din cavitatea bucal i tegumentele fetei depete cu mult jumtatea zonei ocupate de percepia somestezic a ntregului corp. Aceasta datorit importanei fiziologice a senzaiilor produse din cavitatea bucal, densitii i varietii mari a receptorilor din aceast zon. Poriunea inferioar a ariei somestezice unde se proiecteaz gura, buzele, limba cuprinde i aria cortical a gustului (aria 43). Cmpurile 5 i 7 constituie aria somatopsihic de gnozie, ce asigur discriminarea i grupare informaiilor senzitive elementare, de integrare a percepiei presupunnd recunoaterea modelelor. Ariile 39 i 40 fac parte din scoara de asociaie. Aria 40 este un important centru asociativ ce integreaz informaiile ariilor auditive, vizuale i somestezice, iar n emisferul dominat reprezint centrul ideomotor. Sensibilitatea cutanat poate fi suprimat n practica medical, prin anestezie local, rahianestezie i prin narcoz central. n faza instalrii anesteziei ct i n cea a nlturrii ei, se observ disocieri ntre tipurile de sensibilitate cutanat. Astfel, n cazul anesteziei locale, sensibilitatea termic dispare prima i se restabilete ultima. Rahianestezia duce la suprimarea sensibilitii durerose cu pstrarea sensibilitii tactile. Cortexul somestezic este n conexiune cu cortexul motor.
123
SISTEMUL ORO-FACIAL NOIUNI DE FIZIOLOGIE Ediia a doua
Volumul II - ACTIVITATEA MOTORIE
SISTEMUL MUSCULAR ORO-FACIAL
Muchii sistemului oro-facial produc mobilizarea mandibulei, micrile buzelor, limbii i vlului palatin, asigurnd realizarea funciilor motorii specifice acestui sistem.
1.1. ORGANIZAREA FUNCIONAL A SISTEMULUI MUSCULAR ORO-FACIAL
Organizarea musculaturii privind conformaia, dimensiunile, inseriile este astfel constituit ca s realizeze o eficien ct mai mare cu minimum de efort. Randamentul cel mai mare al muchiului se realizeaz cnd fora generat de muchi se aplic perpendicular pe rezistena de deplasat.
124 Sistemul muscular al aparatului oro-facial este organizat pe principiul grupelor musculare antagoniste, ceea ce ofer posibilitatea efecturii unor micri de amplitudini variate i asigurarea unui echilibru dinamic al elementului de deplasat. Grupele musculare din regiunea oro-facial se pot clasifica n: muchii mobilizatori ai mandibulei; muchii pieloi ai feei; muchii limbii; muchii vlului palatului, laringelui, faringelui; muchii regiunilor nvecinate. 1.1.1. Muchii mobilizatori ai mandibulei Cuprind grupele musculare care asigur micrile complexe ale mandibulei, clasificate dup funcia lor i direcia de micare pe care o imprim mandibulei n: muchii ridictori ai mandibulei; muchii cobortori ai mandibulei; muchii propulsori; muchii diductori. 1.1.1.1. Muchii ridictori ai mandibulei sunt: a. muchii temporali: au inserii principale pe scuama oaselor temporale i pe apofizele coronoide. Din punct de vedere funcional, muchiu temporal se mparte n trei fascicule: fasciculul anterior, cu direcie aproape vertical, deplaseaz mandibula n sus; fasciculul mijlociu, are o nclinare oblic spre napoi, realizeaz depla sarea mandibulei n sus i napoi; fasciculul posterior, are fibre cu poziie orizontal, realizeaz retropulsia mandibulei fiind astfel antagonist al pterigoidianului extern. Muchiul temporal particip la: deplasarea mandibulei n sus i posterior; micrile de lateralitate; retropulsia mandibulei, avnd rol n micrile rapide cu contact ocluzal i n masticaia lejer. b. Muchii maseteri se insera pe arcada zigomatic i faa extern a unghiului goniac. Masa muscular a maseterului se difereniaz n trei fascicule: fasciculul superficial al maseterului orientat n jos i posterior; fasciculul mijlociu al maseterului cu traiect vertical; fasciculul profund al maseterului cu traiect vertical. Acest aranjament al fibrelor face ca muchiul maseter s aib cel mai bun avantaj mecanic (eficien mecanic) independent de gradul protruziei mandibulei. Muchiul maseter realizeaz micri puternice de ridicare, propulsie i diducie a mandibulei. c. Muchiul pterigoidian intern are fibrele orientate oblic de sus n jos, dinspre nainte spre napoi i dinuntru ctre nafar,
125 realiznd micri de ridicare, propulsie i diducie a mandibulei. 1.1.1.2. Muchii cobortori ai mandibulei asigur coborrea mandibulei i deschiderea gurii. Pot fi clasificai n: a. muchii cobortori propriu-zii ai mandibulei ce cuprind: muchiul milohioidian care mpreun cu cel din partea opus realizeaz o adevrat diafragm ce formeaz planeul gurii, delimitnd astfel un etaj supramilohioidian i unul submilohioidian; muchiul geniohioidian se prezint ca un muchi aplatizat de sus n jos; muchiul digastric, pntecele anterior. Aceti muchi coboar mandibula dup ce osul hioid a fost fixat de muchii infrahioidieni (subhioidieni). b. muchii cobortori indireci ai mandibulei cuprind muchii care stabilizeaz osul hioid i astfel particip indirect la coborrea mandibulei. Ei sunt reprezentai de: muchiul stilohioidian, ridic osul hioid iar odat cu el i scheletul laringelui, micare deosebit de important pentru deglutiie; muchii subhioidieni: m. sternohioidian (conecteaz sternul, cartilajul tiroid i osul hioid), m. omohioidian (se insera posterior pe omoplat), m. sternotiroidian, m. tirohioidian. Contracia acestor muchi coboar i stabilizeaz osul hioid, favoriznd aciunea de deschidere a gurii, a muchilor suprahioidieni. Muchii hioidieni n raport cu aciunea lor se pot clasifica n: muchiul digastric, pntecele posterior;
126 muchii suprahioidieni: digastric, stilohioidian, milohioidian, geniohioidian, care sunt ridictori ai hioidului cnd se contract simultan i au o inserie fix pe mandibul. Contracia simultan a muchilor milohioidieni ridic planeul bucal i secundar aplic limba pe bolta palatin, care apas, ca un piston, bolul alimentar spre faringe, deci aceti muchi particip direct la timpul bucal al deglutiiei. n cazul cnd osul hioid este fixat de ceilali muchi hioidieni, con tracia simultan a muchilor suprahioidieni (excepie m. stilohioidian) determin coborrea mandibulei. muchii subhioidieni: sternocleidohioidian, sternotiroidian, tirohioidian, omohioidian. Toi muchii subhioidieni sunt cobortori ai osului hioid, cnd se contract simultan i au inserie fix pe stern i clavicul. M. tirohioidieni ridic laringele, cnd captul hioidian este fixat pe m. suprahioidieni, avnd astfel rol n timpul faringian al deglutiiei. Prin intermediul muchilor suprahioidieni, muchii subhioidieni pot deveni cobortori ai mandibulei dac punctul fix al inseriilor lor l constituie sternul i clavicula. Contracia bilateral i simultan a muchilor omohioidieni deplaseaz hioidul napoi. 1.1.1.3. Muchii propulsori ai mandibulei proiecteaz nainte osul mandibular, realiznd micarea de propulsie. In aceast categorie de muchi sunt inclui: m. pterigoidian extern, constituit din dou fascicule: fasciculul superior orientat uor oblic ctre nafar, n jos, napoi i fasciculul inferior cu direcie aproape orizontal. Acesta reprezint principalul muchi propulsor; m. maseter, prin fasciculul superficial; m. pterigoidian intern, cu aciune propulsoare secundar. 1.1.1.4. Muchii retropulsori sunt reprezentai de m. temporali (prin fasci culul posterior), m. maseter (prin fasciculul superficial) i m. pterigoidian intern. Grupele antagoniste ale muchilor mobilizatori ai mandibulei sunt:
127 a. n sens vertical: ridictori: maseter, pterigoidian intern, fasciculul anterior i m. mijlociu al temporalului; cobortori: geniohioidian, milohioidian, pntecele anterior al m. digastricului, pterigoidian extern; b. n sens sagital: propulsori: pterigoidian extern, intern, fasciculul superficial al m. maseterului; retropulsori: fasciculul posterior al temporalului, fasciculul pro m. fund al maseterului, geniohioidian, pntece posterior al digastricului; c. n plan transversal: m. pterigoidian extern, n contracie unilateral, 1.1,2, Muchii pieloi ai feei Muchii pieloi ai feei sunt muchi subiri, aplatizai, au una sau ambele inserii mobile la nivelul tegumentului, prin contracia lor realizeaz expresivitatea feei. Muchii pieloi ai feei localizai n jurul orificiilor (muchii perio-rali, ai narinei, ai fantei palpebrale) acioneaz ca dilatatori sau constrictori ai acestor orificii. Muchiul orbicularul buzelor are un rol deosebit n sugere (succiune), n prehensiunea alimentelor, n masticaie, n pronunia consoanelor Iabiale ("b", "p"), n pronunia vocalelor "o", "u", n scuipat, suflat, fluierat, srutat, etc. 1.1.3, Muchii limbii Muchii intrinseci i extrinseci ai limbii (vezi capitolul Limba din volumul I, pag, 113) sunt considerai mpreun cu muchii oro-faciali ca muchi auxiliari ai masticaiei, intervenind n meninerea alimentelor pe suprafaa ocluzal i deplasarea lor pe o arcad sau alta. 1.1.4. Muchii vlului palatului, laringelui, faringelui Muchii vlului palatului cuprind cinci perechi de muchi care au rolul de a nchide nazofaringele n timpul deglutiiei i de a separa cavitatea bucal de bucofaringe i de laringe n timpul deglutiiei i al fonaiei. Prin con tracia i relaxarea coordonat a muchilor vlului palatin se modific istmurile dintre cavitatea bucal i cea faringian contribuind efectiv la masticaie* deghitiie, fonaie i respiraie, Muchii laringelui pot fi grupai n dou grupe antagoniste: muchii constrictori ai glotei care sunt i adductori ai coardelor vocale, muchi inervai de nervul recurent. Paralizia unilateral a nervului recurent determin o voce bitonal; muchii dilatatori ai glotei care deprteaz coardele vocale. In timpul vorbirii sau cntatuhri, muchii laringelui se contract sau se relaxeaz coordonat asigurnd
128 nlimea sunetelor, intensitatea vocii. n timpul inspirului se contract muchii dilatatori ai glotei. n timpul expiraiei i al emiterii sunetelor nalte (acute), laringele este ridicat n sus i nainte. Laringele este un organ cu mare mobilitate. Laringele este ridicat vertical i puin nainte la fiecare deglutiie, la expiraie forat i n timpul emiterii sunetelor nalte. Laringele coboar n timpul inspiraiei i n emiterea sunetelor joase. 1.1.4,3. Muchii faringelui Faringele este un conduct musculo-membranos cu direcie vertical la om, situat la rspntia dintre cavitatea nazal, bucal, pe de o parte i laringele, gura esofagului, pe de alt parte. Bolul alimentar i coloana de aer, trec obligatoriu prin faringe ncrucindu-i drumurile. Faringele este i o cutie de rezonan. Afeciunile faringiene se nsoesc de tulburri de deglutiie, respiraie, fonaie. Musculatura faringelui este reprezentat de cinci perechi de muchi dintre care trei sunt constrictori i doi ridictori. Contracia succesiv i coordonat a muchilor constrictori determin unda peristaltic ce conduce bolul alimentar spre esofag. Muchii ridictori ai faringelui ridic faringele i cartilajul tiroid n timpul faringian al deglutiiei. Paralizia muchilor constrictori ai faringelui (difterie, leziuni bulbare) precum i contraciile spastice tetanos, histerie, turbare) fac imposibil deglutia, alimentele refulnd spre fosele nazale sau spre laringe. 1.1.5. Muchii regiunilor nvecinate (muchii asociai aparatului dento-maxilar) Muchii regiunilor nvecinate cuprind o serie de grupe musculare, care, i mod secundar i asociat, mobilizeaz mandibula sau particip la efectuare! funciilor principale ale aparatului dento-maxilar (masticaia, deglutiia, fonaia mimica). Din aceast grup de muchi asociai reinem: m. pielosul gtului: ridic pielea gtului exprimnd frica, groaza; m. flexori ai capului, dintre care sternocleidomastoidianul imprima capului micarea de flexie (prin contracie bilateral), nclinarea laterala (prin contracie unilateral) sau rotarea (contracie contralaterala). De asemenea, este un muchi inspirator accesoriu; m. extensori ai capului prin tonicitatea lor menin capul n poziia de echilibru pe coloana cervical. Hipertonia muchilor extensori determin, c timpul, retrognaia mandibular iar hipertonia muchilor flexori ai capuli determin prognaia mandibular. Tabelul 1. Principalii muchi ai aparatului dento-maxilar (dup Boboc, 1996, modificat)
129 Denumirea muchiului Digastric, pntece anterior Digastric, pntece posterior Inervaie N. milohioidian, ramur din n. alveolar inferior Ramur din n. facial Aciune Observaii
Coborrea i retropulsia mandibulei Acine indirect de coborre a mandibulei a) ridic osul hioid i trage nainte laringele; b) coboar mandibula Ridic osul hioid i coboar mandibula Coboar comisura i ridic pielea gtului Propulsor, cobortor i n micarea de lateralitate Propulsia se nsoete i de o coborre n ATM: glisare anterioar i rotaie In masticaie coboar mandibula; In deglutiie ridic hioidul.
Milohioidian
N. milohioidian, ramur din n. alveolar inferior Ramur din n. hipoglos
Geniohioidia n Muchiul pielos al gtului Pterigoidian extern, fascicul superior Pterigoidian extern, fascicul inferior
N. facial
N. mandibular
N. mandibular
Temporal a) fascicul anterior b) fascicul mijlociu c) fascicul posterior
N. temporal profund ant. N. temporal profund mijlociu N. temporal profund posterior
Ridic mandibula Retropulsia mandibulei
Rezultanta celor trei fascicole: n sus i uor napoi. Masticatorii toctori dezvolt fasciculul posterior; Masticatorii frectori dezvolt
130 Maseter a) fascicul superficial b) fascicul profund Influen foarte puternic asupra reg. goniace, condilului, arcadei zigoma-tice i osului malar Ridictor i determin micri de lateralitate de partea opus contraciei Primete fascicule din muchii nvecinai
N. maseterin N. maseterin
Oblic nainte i n sus; Vertical
Pterigoidian intern Orbicularul extern
N. pterigoidian medial N. facial
Direcie oblic n sus, nuntru i nainte nchide orificul bucal
Orbicularul intern (sfincterul gurii) Triunghiular al buzelor Ptrat al buzei superioare Zigomatic
N. facial
N. facial N. facial N. facial
nchide orificiul bucal apropie buzele i Ic aplic pe Coboar comisura Ridic buza superioar Trage comisura n sus i n afar Trage buza inferioar n jos Trage buza superioar n sus Apas buza superioar pe dini Apas buza inferioar pe dini Ridic i face proeminent buza inferioar, ncreete pielea brbiei
Arc dou fascicule ce se ntretaie la comisur M. tristeei i dispreului
Antagonist cu m. triunghiular
Ptrat al buzei inferioare Canin superior Incisiv superior Incisiv inferior Mentonier
N. facial N. facial N. facial N. facial N. facial
131 Buccinator N. facial a) cnd gura e goal, trage de comisur i apropie buzele, le apas pe dini, lrgete fanta labial b) gura plin cu aer: contracia comprim aerul i-1 expulzeaz cu presiune Rol n rs i plns; antagonist cu musculatura limbii privind dezvoltarea transversal a arcadelor dentare
Receptorii musculari sunt de tip mecano-receptori cuprinznd: fusurile neuro-musculare i organele tendinoase Golgi, n principal, ca i corpusculii Pacini, terminaiile libere nemielinizate. 1.2.1. Fusul neuro-muscular Fusul neuro-muscular este o formaiune proprioceptiva aezat n paralel cu fibrele musculare striate extrafusale. 1.2.1.1. Structura fusului neuro-muscular Fiecare fus neuro-muscular conine aproximativ zece fibre musculare nvelite de o capsul de esut conjuctiv. Aceste fibre musculare cu caractere embrionare, ce prezint striaii numai la capete, se numesc fibre intrafusale. Fibrele intrafusale sunt situate n paralel cu restul fibrelor musculare, cu proprieti contractile, numite fibre extrafusale. Fibrele intrafusale se insera la un capt pe tendonul muchiului, iar la cellalt capt pe locul de inserie al fibrelor extrafusale. Fibrele intrafusale, la mamifere, sunt de dou tipuri numite: fibre cu sac nuclear: care prezint nucleii adunai ntr-o zon central, dilatat, iar la capete prezint striaii; fibre cu lan nuclear, la care nucleii sunt aezai central, ntr-un ir unic.
1.2. RECEPTORII MUSCULARI
132
Fig. 1. Schema fusului neuromuscular Ambele tipuri de fibre intrafusale prezint activitate contractil numai n zona capetelor striate. 1.2.1.2- Inervaia senzitiv a fibrelor intrafusale Inervaia senzitiv a fibrelor intrafusale este asigurat de: a. fibre senzitive primare sau anulo-spirale, sunt terminaii ale fibrelor aferente de tip Ia, cu vitez de conducere rapid (70-120 m/sec.)- Acestea se nfoar n jurul zonelor cu sac nuclear, cu nuclei n lan, transmit informaiile kinestezice musculare privind gradul de ntindere al muchiului, permind adaptarea tonic de postura i micare, la variaia lungimii i vitezei de ntindere muscular; b. fibre senzitive secundare, sunt terminaii "n buchet" ale fibrelor senzi tive de grup II, sunt localizate numai la capetele fibrelor cu nuclei n lan. Au rol n sesizarea i semnalarea alungirii instantanee a muchiului.
133 1.2.1.3. Inervaia motorie a fibrelor intrafusale Inervaia motorie a fibrelor intrafusale este asigurat de fibre nervoase cu diametru de 3-6 micrometri, ce constituie aproape 30% din fibrele rdcinii ventrale, aparin grupului A gama din Erlanger i Gasser, se numesc fibre gama eferente sau sistemul nervilor motori mici. Aceste fibre inerveaz exclusiv fusurile neuro-musculare avnd dou tipuri histologice de terminaii: "n plac", pe fibrele intrafusale cu sac nuclear, terminaii ale fibrelor gama "dinamice"; "cuadrilat", pe fibrele intrafusale cu lan nuclear, terminaii ale fibrelor gama "statice". De asemenea, neuronii motori mari, beta inerveaz att fibrele intrafusale ct i fibrele extrafusale. Stimularea axonilor motori gama i beta produce dou tipuri funcionale de rspunsuri i anume: a. rspuns "static": cnd poriunea receptoare a fusului neuromuscular este ntins ncet, numrul impulsurilor transmise prin fibrele senzitive este direct proporional cu gradul de ntindere al fusului i dureaz cteva minute (atta timp ct receptorul este ntins). Rspunsul static este dependent de fibrele cu lan nuclear, fibre inervate att de terminaii anulo-spirale ct i de cele "n buchet". Se realizeaz contracia muchiului pe toat perioada ct muchiul este meninut la o lungime excesiv. b. rspuns "dinamic": se declaneaz o contracie instantanee puternic cnd lungimea fusului crete rapid, brusc, iar numrul de impulsuri transmise prin fibrele senzitive primare este foarte mare, deoarece aceste fibre rspund extrem de activ la o rat rapid de schimbare a lungimii fusului. Chiar dac lungimea fusului se modific numai cu o fraciune de micrometru ntr-un timp extrem de scurt (o fraciune de secund) fibrele primare transmit explosiv numrul impulsurilor n exces. Imediat ce lungimea fusului nceteaz s creasc, rata impulsurilor revine la un nivel inferior celui static (fig. 2).
134
Cnd receptorul fusului se scurteaz, aceast schimbare momentan scade impulsurile prin fibrele senzitive primare. De ndat ce receptorul i stabilizeaz noua lungime, reapar impulsuri n fibrele Ia. Rspunsul dinamic este dependent de stimularea fibrelor cu sac nuclear, fibre inervate numai de terminaii primare Ia. 1.2.1.4. Controlul rspunsului dinamic i static prin neuronul motor gama Neuronii motori gama ai fusului neuro-muscular se pot mpri n dou tipuri: a. neuroni gama dinamici, care inerveaz numai captul fibrelor cu sac nuclear inducnd un rspuns exploziv muscular n timp ce rspunsul static este puternic afectat- Stimularea eferenelor dinamice crete sensibilitatea fusului la rata variaiei de lungime. b. neuroni gama statici , care inerveaz fibrele cu nuclei n lan inducnd rspunsul static, care are mica influen asupra rspunsului dinamic. Stimularea eferenelor statice crete sensibilitatea fusului la lungime constant, de repaus. In condiii normale cnd exist un numr redus de excitaii prin neuronul gama, fusurile neuro-musculare trimit impulsuri continue prin fibrele senzitive, ntinderea fusului determin creterea ratei de impulsuri, iar scurtarea induce scderea ratei de impulsuri senzitive. Astfel, fusurile neuro-musculare pot trimite la mduva spinrii: semnale "pozitive", adic un numr crescut de impulsuri, indicnd creterea lungimii muchiului; semnale "negative", cnd numrul de impulsuri este sub nivelul normal semnificnd c muchiul nu este ntins. Ambele tipuri de rspunsuri ale fusului neuro-muscular sunt importante pentru controlul contraciei musculare. Controlul descrcrilor gama eferente Neuronii motori ai sistemului gama eferent sunt reglai de tracturile descendente cu origine n diferite arii corticale. Prin aceste ci, sensibilitatea fusurilor neuro-musculare poate fi ajustat i modificat n funcie de necesiti, n vederea realizrii controlului postural. Ali factori pot, de asemenea, influena descrcrile gama eferente. Astfel, anxietatea crete descrcrile n neuroni iar reflexele tendinoase sunt hiperactive. Stimularea cutanat, mai ales de ageni nocivi, crete descrcrile gama eferente la fusurile neuromusculare din flexorii ipsilaterali cu scderea impulsurilor ia extensori. 1.2,1.5, Reflexul miotatic Cea mai simpl manifestare a funcionrii fusurilor neuromusculare este reflexul muscular la ntindere numit i reflexul miotatic (fig. 3). Principiul reflexului miotatic: ntinderea muchiului determin excitarea fusurilor neuro-musculare inducnd reflex contracia unui mare numr de fibre din acelai muchi sau n muchii sinergici
apropiai. Circuitul neuronal al reflexului miotatic Fibrele senzitive de tip Ia, cu origine n fusurile neuro-musculare, intr n rdcina dorsal a mduvei spinrii i trec direct n cornul anterior al substanei cenuii. Aici, fac sinaps cu neuronul motor care se termin n muchiul de origine al fusurilor neuro-musculare, printr-o plac motorie. Deci, este o cale mono-sinaptic, prin care semnalul determinat de excitarea fusurilor neuro-musculare se ntoarce napoi la muchi, ntrzierea sinaptic fiind minim. Mediatorul chimic al sinapsei centrale este glutamatul.
Neuron motor
Fus neuromuscular
Fig.3 Circuitul neuronal al reflexului miotatic Reflexul miotatic Informaiile venite prin fibrele senzitive secundare pot urma dou ci: s se termine, monosinaptic, n neuronii motori anteriori; s fac sinaps cu multiplii interneuroni din substana cenuie a mduvei (neuroni intercalri) conectai cu neuronul motor din cornul anterior. Aceast cale este multisinaptic, prezentnd o ntrziere a transmiterii semnalului (fig. 4). Funciile fusurilor neuro-musculare Reflexul miotatic pozitiv induce contracia muchiului declanat de ntinderea sa, deci se opune ntinderii muchiului, iar reflexul miotatic negativ se opune scurtrii muchiului. Deci, reflexul miotatic tinde s menin lungimea constant a
muchiului, aa numitul "status quo" al muchiului. Fusurile neuro-musculare i conexiunile reflexe realizeaz un feed-back ce conlucreaz Ia meninerea lungimii muchiului: dac muchiul este ntins (activ sau pasiv) se genereaz poteniale de aciune n terminaiile senzitive, care prin intermediul neuronului motor alfa induc contracia muchiului, respectiv scurtarea sa. Cnd muchiul este scurtat, scad potenialele de aciune n terminaiile senzitive, motorii i muchiul se va relaxa, respectiv se va ntinde, iar ciclul se reia. Fusurile neuro-musculare au rol i n activitatea motorie voluntar-Semnalele transmise din cortexul motor determin simultan att stimularea
neuronilor motori alfa ct i a neuronilor gama printr-un mecanism numit coactivare. Aceasta produce contracia simultan a fibrelor intrafusale ct i extra-fusale. Scopul contraciei, fibrelor intrafusale n acelai timp cu fibrele extra-fusale, din muchiul scheletic este dublu i anume: aceasta pstreaz lungimea poriunii receptoare a fusului din muchiul n schimbare n opoziie cu contracia muchiului (muchiul se contract, se scurteaz, dar lungimea poriunii receptoare rmne aceiai);
menine funcia de amortizare a fusului, indiferent de modificarea lungimii muchiului, evitndu-se astfel variaiile de lungime a fusului n sensul unei suprantinderi sau adunri, (plieri) excesive, situaii n care fusul nu ar funciona n condiii optime. Sistemul gama eferent este excitat prin semnale din regiunea facilitatoare, bulbo-reticulat i secundar de impulsuri transmise n aria bulbo-reticulat din cerebel, nueleii bazali, cortex cerebral. Sistemul facilitator bulbo-reticulat este n principal responsabil de contraciile muchilor antigravitaionali care au o mare densitate de fusuri neuro-musculare. n aceste condiii, mecanismul gama eferent are importana n amortizarea micrilor diferitelor pri ale corpului n timpul mersului sau alergrii. Una din cele mai importante funcii ale fusurilor neuro-rnusculare este de a stabiliza poziia corpului. Pentru aceasta sistemul facilitator bulbo-reticulat transmite semnale prin fibrele nervoase gama la fibrele musculare intrafusale determinnd contracia capetelor striate i ntinderea regiunii centrale receptoare cu inducerea de poteniale de aciune n fibrele receptoare. Activarea n acelai timp a fusurilor n ambele pri ale unei articulaii determin contracia reflex a muchilor de o parte i de alta a articulaiei rezultnd stabilizarea puternic a articulaiei. In continuare orice for ce tinde s mobilizeze articulaia din aceast poziie este anihilat printr-un reflex miotatic de mare sensibilitate. Aspectele clinice ale reflexului miotatic In examinarea clinic curent explorarea reflexelor osteotendinoase are o deosebit importan. Exemplu: reflexul rotulian percuia, cu un ciocan de reflexe, a tendonului muchiului cvadriceps iniiaz un reflex miotatic dinamic urmat de extensia gambei pe coaps (prin contracia cvadricepsului). Reflexul este sczut sau absent cnd impulsurile facilita torii sunt deprimate sau abolite. Compresiunea de Muchi natur tumorala, distrugerea unor zone din cor-texul motor contralateral determin exacerbarea reflexului rotulian. 1.2.2, Organul tendnos Golgi Organul tendinos Golgi cuprinde o reea de terminaii nervoase butonate, ncapsulate, printre fasciculele tendonului (fig. 5). La fiecare organ tendinos Golgi sunt conectate n serie ntre 10-15 fibre musculare. Fibrele senzitive din organul tendinos Golgi fac parte din grupul Ib, sunt mielinizate, au diametrul 16 micrometri, conduc rapid informaiile senzitive. Organul tendinos Golgi este stimulat de tensiunea realizat n muchi, deci detecteaz tensiunea muscular spre deosebire de fusurile neuro-musculare care detecteaz lungimea i variaiile lungimii muchiului. Aceste fibre senzitive (Ib) transmit impulsurile att local n
mduva spinrii ct i pe cile ascendente spre cerebel (prin fracturile spino-Fig. 5. Organul tendinos Golgi cerebeloase cu o vitez de 120 m/sec.) sau spre (dup Guyton, 1996, modificat) cortexul cerebral. Semnalul local medular excit
FIG. 6. Reflexul de flexie si reflexul de extensie I ncrucisata un interneuron inhibitor, care la rndul su inhib motoneuronul din cornul anterior al mduvei spinrii. Deci, stimularea fibrelor Ib dintr-un muchi este urmat de producerea unor poteniale de inhibiie postsinaptic n neuronul motor care inerveaz muchiul a cror fibre Ib au fost excitate (fig. 6). Acest circuit local inhib direct un anumit muchi fr sa afecteze muchii adiaceni. Pentru c organul Golgi este situat n serie cu fibrele musculare, este stimulat att de ntinderea pasiv a tendonului ct i de contracia activ a muchiului. Gradul de stimulare prin ntindere pasiv nu este mare pentru c fibrele elastice preiau o mare parte din ntindere. Stimularea obinuit este produs prin contracia muchiului i astfel organul Golgi funcioneaz ca un transductor n circuitul de feed-back care regleaz fora muscular. Organul tendinos Golgi are dou tipuri de rspunsuri: rspunsul dinamic: cnd tensiunea muchiului crete rapid, instantaneu; rspunsul static: de repaus sau bazai care este direct proporional cu tensiunea muchiului la un moment dat. Creterea tensiunii muchiului stimuleaz organul Golgi, semnalul fiind transmis n mduva spinrii determin reflexul inhibitor al muchiului respectiv. Acest reflex acioneaz ca un mecanism de feed-back negativ care previne dezvoltarea unei tensiuni prea maris n muchi. Cnd tensiunea n muchi j respectiv n tendon devine
extrem, efectul inhibitor declanat de organul Golgi poate fi aa de mare nct s induc o relaxare instantanee a muchiului n totalitate. Acest efect se numete reacia de ntindere (alungire). Reflexul de alungire este un mecanism protectiv de prevenire a ruperii muchiului sau a desprinderii tendonului din inseria osoas. Flexia pasiv a cotului induce imediat un reflex de ntindere n muchiul triceps. ntinderea activeaz reflexul invers ntinderii. Rezistena la flexie induce colapsul instantaneu i braul se flecteaz. Flexia pasiv ntinde muchiul din nou i ciclul se poate relua. Un alt rol al organului tendinos Golgi este de a egaliza forele contractile a fibrelor musculare separate: fibrele care realizeaz o tensiune n exces vor fi inhibate reflex, iar cele care realizeaz o tensiune prea mic vor deveni mai excitate pentru c reflexul de inhibiie este absent. Astfel se previn leziunile n anumite zone ale muchilor unde un numr mic de fibre sunt supratensionate, Rezistena muchiului la ntindere este definit ca tonusul muchiului. Un muchi, a crui nerv motor a fost secionat, are o mic rezisten la ntindere i se spune c este flasc. Muchiul hipertonic (spastic) este acel muchi n care rezistena la ntindere este mare datorit reflexelor de ntindere hiperactive. ntre aceste dou aspecte (flasc i spastic) este starea de tonus normal. n general, muchii sunt hipotonici cnd rata descrcrilor pe neuronul gama eferent este mic i sunt hipertonici cnd rata este mare. 1.2.3. Corpusculii Pacini Corpusculii Pacini sunt inervai senzitiv de fibre nervoase cu o grosime de 1-4 micrometrij sunt stimulai la presiune, micri rapide, vibraii i ntr-o mic msur la ntindere. Sunt veritabili detectori de acceleraie. 1.2.4. Terminaiile libere nemielinizate Terminaiile libere nemietinizate sunt de fapt receptori ai durerii la nivelul muchiului i tendoanelor. Aceti receptori sunt activai n cursul oboselii, spasmului, ischemiei prin contracii musculare prelungite. 1.2.5. Receptorii din musculatura sistemului oro-facial n muchii ridictori ai mandibulei exist un numr important de fusuri neuro-musculare n schimb muchii cobortori ai mandibulei au un numr redus de fusuri neuro-musculare.Inervaia senzitiva a fusurilor neuro-musculare din muchii masticatori este asigurata de neuronii unipolari mari din nucleul mezencefalic al trige-menului. Acetia trec far s fac sinaps n ganglionul Gasser. Prelungirile acestor neuroni dau colaterale care formeaz conexiuni cu neuronii motori alfa ai muchilor ridicatoTi, din nucleul motor al trigemenului. Se constituie, astfel, arcuri reflexe monosinaptice ce reprezint suportul morfologic al reflexului miotatic mandibular (maseterin). Existena receptorilor Golgi n muchii masticatori este o problem
controversat. Stimularea receptorilor Golgi ar induce inhibiia motoneuronilor alfa ai muchilor ridictori, n mod similar cu ceea ce se produce n cazul reflexelor miotatice ale membrelor. Receptorii fusali din muchii gtului intervin indirect n controlul micrilor masticatorii. Poziia capului are o influena important asupra ritmului i forei de masticaie.
1.3. TONUSUL MUSCULAR
Tonusul muscular reprezini starea de uoar contracie (semicontracie) a muchiului striat n repaus. Tonusul muscular este o stare fiziologic fundamental a muchilor scheletici i poate avea mai multe forme: tonus de repaus; tonus de postur; tonus de atitudine; tonus de comportament. Tonusul muscular caracterizeaz muchii n organismul intact, are o mare variabilitate n funcia de o multitudine de factori. 1,3.1. Mecanisme tonigene Rolul principal n realizarea mecanismelor tonigene l are sistemul nervos. La originea mecanismului tonigen sta reflexul medular miotatic (de ntindere). Arcul reflex miotatic cuprinde: receptor: fusul neuro-muscular; ci senzitive: aferentele de tip la i II; centrii medulari; ci eferente: motoneuroni alfa pentru fibrele extrafusale i motoneuroni gama pentru fibrele intrafusale. Fusul neuro-muscular intervine n controlul nervos, incontient al contraciei musculare, n cursul micrii i al contraciei statice susinute. Reflexul miotatic este influenat de centrii supramedulari din trunchiul cerebral, nucleii bazali, formaiunea reticulat, cortexul frontal i cerebel. Importana funcional a tonusului muscular este complex:
tonusul de fond, de postur asigur fixitatea articulaiilor i amortizarea elastic a micrilor; creterea tonusului muscular este un element important n procesul de termogenez; strile emoionale stressante are loc o cretere a tonusului n mus cular de fond. 1.3.2. Tonusul muchilor masticatori Tonusul muscular caracterizeaz att starea de repaus mandibular ct i starea de micare. n starea de repaus i de postur mandibular, tonusul muchilor masticatori are rol de a menine o anumit relaie spaial ntre diversele segmente scheletice mobile ale extremitii cefalice. 1.3.2.1. Rolul motoneutonului alfa Tonusul muchilor masticatori este realizat n principal de ctre motoneuronul alfa din nucleii motori ai nervilor cranieni corespunztori, care prezint un ritm tonigen propriu de descrcare a impulsurilor. Fiecare axon al neuronului alfa din nucleul motor al trigemenului, inerveaz un numr variabil de fibre striate, existnd o anumit topografie bine stabilit a nucleului motor trigeminal. De asemenea, exist o alternan a activitii unitilor motorii ce asigur tonusul maseterului, n sensul c unele descarc impulsuri tonigene, iar altele sunt n repaus. Avantajele acestei activiti alternante sunt; muchiul nu obosete; activitatea tonic nu este obositoare pentru muchi; consumul energetic este mic. Motoneuronul gama, din nucleul trigeminal, intervine indirect n realizarea tonusului muscular prin aciune asupra fibrelor intrafusale ale fusului neuro-muscular. 1.3.2.2. Reglarea activitii tonice a motoneuronului alfa n reglarea activitii tonigene a motoneuronului alfa intervin mai multe circuite: a. circuitul neuronului intercalar Renshaw care prin neuronul Renshaw realizeaz un mecanism de inhibiie recurent fiecrui neuron motor trigeminal. Neuronul intercalar Renshaw are o sinaps axo-somatic cu motoneuronul alfa iar acesta trimite o colateral recurent pe corpul neuronului intercalar. Acest circuit asigur o frecven constant de descrcare a impulsurilor n motoneuronul alfa. Cnd crete frecvena de descrcare iar tonusul muchilor masticatori tinde s creasc, intervine neuronul intercalar Renshaw care inhib motoneuronul alfa.
Circuitul Renshaw funcioneaz astfel ca un limitator de frecvent. b. Aferentele corticale sunt asigurate prin fracturile corticonucleare care duc comanda la nuciei motori ai nervilor cranieni V, VII, XII, dar i prin intermediul fibrelor cortico-reticulare. c. Aferente hipotalamice din hipotalamusul posterior cresc frecvena de descrcare a neuronului gama. d. Aferentele reticulare: substana reticuiat intervine n reglarea tonusului muscular prin intermediul buclei gama. Activarea substanei reticulale facilita toare induce creterea descrcrilor n motoneuronul gama urmat de creterea tonusului muscular. e. Aferentele cerebeioase: cerebelul are conexiuni directe cu motoneuronii alfa i indirecte cu motoneuronii gama (prin intermediul substanei reticulate). Cerebelul, prin intermediul substanei reticulate, excit sau inhib circuitele tonic reglatoare. f. Aferentele periferice venite de la fusurile neuro-musculare realizeaz reflexul miotatic de reglare automat a tonusului striat. Muchii ridictori ai mandibulei prezint mai multe fusuri neuromusculare dect muchii cobortori. Repartiia fusurilor neuromusculare: nm. temporal - 217, n maseter- 150, n pterigoidianul intern - 155, n pntecele anterior al digastricului - 6. Inervaia senzitiv a fusurilor neuro-musculare este asigurat de fibrele Ia i II ce sunt dendrite ale neuronilor localizai n nucleul mezencefalic ai trigeme-nului. Modularea sensibilitii fusurilor neuro-musculare se realizeaz prin motoneuronul gama care pune n tensiune fibrele intrafusale, similar unei ntinderi brusce. 1.3.3. Reflexul miotatic maseterin Reflexul miotatic reprezint contracia reflex a unui muchi indusa de propria sa ntindere. Mecanismul reflexului miotatic st la baza contraciei musculare att statice (tonigene) ct i dinamice, fazice, active. ntinderea brusca a muchiului provoac un rspuns scurt numit reacia fazic a reflexului miotatic. O ntindere lent, de durat mai mare determin un rspuns sub forma reaciei statice la ntindere, cu o frecvent de descrcare joas de 7-20/sec. Reaciile statice miotatice sunt capabile s asigure starea de postur antigravitaional, pe cnd reaciile fazice apar n timpul micrii. Reflexul miotatic maseterin are aceleai caracteristici statice i fazice ca i oricare alt reflex miotatic, ntinderea maseteruiui prin coborrea mandibulei, induce reflex contracia maseterului cu ridicarea mandibulei. 1.3.3.1- Elemente structural-funcionate ale reflexului miotatic maseterin Arcul reflexului miotatic maseterin cuprinde: - receptorul: reprezentat de fusurile neuro-musculare din muchiul maseter;
calea aferent: fibrele anulo-spirale, prelungiri dendritice ale neuronilor din nucleul mezencefalic al trigemenului, care fac sinaps n nucleul motor al trigemenului cu motoneuronul alfa; centrul nervos: nucleul motor al trigemenului; calea eferent: ramura motorie a trigemenului; organ efector: muchiul maseter (n principal) mpreun cu ceilali muchi ridictori ai mandibulei. Examinarea reflexului maseterin: percuia mentonului, cu gura ntre deschis, induce rapid contracia maseterului i nchiderea gurii, n condiii normale maseterul se afl ntr-o permanent solicitare prin aciunea continu a gravitaiei, ce se exercit asupra lui. Reflexul miotatic de ntindere este un fenomen de adaptare neuromuscular la solicitrile antigra-vitaionale. Reflexul miotatic maseterin servete la meninerea constant a lungimii maseterului i a tonusului su care, alturi de ceilali muchi ridictori, particip la pstrarea mandibulei ntr-o poziie de postur convenabil. Slbirea tonicitii i coborrea mandibulei duc la ntinderea fusurilor neuro-musculare i excitarea fibrelor anulospirale, informaie ce ajunge n nucleul mezencefalic al trigemenului. De aici, stimulul ajunge la nucleul motor al trigemenului iar prin motoneuronul alfa determin creterea tonusului maseterului i redresarea mandibulei, n sensul ridicrii ei, 1.3.3.2. Reglarea reflexului miotatic maseterin Reflexul miotatic maseterin este influenat de: a, aferentele senzoriale tonigene, de la proprioceptorii musculari, articulari, exteroceptori din muchii cefei, respectiv regiunea cefalic; b. aferentele vestibulare care semnalez, n orice moment, poziia capului n spaiu, deviaiile sale n raport cu poziia de referin i deplasrile capului. Aparatul vestibular, prin receptorii si de poziie i de micare, participa la distribuia echilibrat a tonusului n funcie de echilibrul static i dinamic al organismului. Aferentele vestibulare influeneaz reflexul miotatic prin: realizarea unui tonus de fond crescut, care crete amplitudinea rspunsului; prin conexiuni vestibulo-motorii; prin substana reticulat care modeleaz bucla gama. Excitarea labirintului modific tonusul maseterin prin conexiunile directe vestibulo-motorii ce influeneaz neuronul alfa din nucleul trigeminal i substana reticulat facilitatoare (care prin bucla gama crete tonusul de ambele pri, dar mai ales-de partea labirintului excitat). Stimulii vestibulari induc modificri de tonus ale muchilor cefei dar i ale muchilor din sistemul oro-faciat Poziia capului influeneaz ritmul i fora de masticaie,
fenomene demonstrate pe trasee electromiografice. Contracia tonic a muchilor cefei poate contrabalansa greutatea capului n poziie de veghe. De fapt, prehensiunea alimentelor, masticaia, vorbirea, mimica nu se pot desfura dect n poziia de veghe a capului.
FIZIOLOGIA ARTICULAIEI TEMPORO-MANDIBULARE
Articulaia temporo-mandibular (ATM) este considerat cea mai complex articulaie din organismul uman care prin structura particular particip la realizarea funciilor motorii de baz ale sistemului oro-facial: masticaia, deglutiia, fonaia. ATM stabilete legtura mobil dintre baza craniului i mandibul pe de o parte, iar pe de alt parte, permite micrile suprafeei articulare dentare inferioare pe cea superioar n vederea efecturii funciilor eseniale ale aparatului dento-maxilar. Relaiile dintre ATM i articulaia dentar sunt amortizate de sindesmozele alveolo-dentare i sistemele traiectoriale dentare, care preiau toate presiunile exercitate de dini pentru a le dispersa pe toat suprafaa cranian. ATM poate realiza micri foarte variate datorit laxitii ligamentelor i absenei de creste osoase n jurul cavitii glenoide. Micrile ATM au o predominant n direcia sagital, sau transversal, sau vertical, n raport cu tipul dinamic masticator. Micarea ATM caracteristic pentru specia uman este o micare de translaie a condilului i a meniscului, asociat cu micarea de rotaie a lui n timpul deschiderii i al micrilor de lateralitate. Malocluziile dentare determin tulburri n sistemul de reglare, care se materializeaz n leziuni parodontale sau articulare. 2.1. MORFOLOGIA ATM ATM este singura articulaie mobil a aparatului dento-maxilar care vine n raport cu o alt articulaie, mult mai mobil, realizat de dini. 2.1.1. Suprafee osoase Suprafaa osoas articular cranian este reprezentat de o poriune convex (tubercul articular) i de o poriune posterioar concav numit cavitate glenoid acoperit de un fibrocartilaj. Cavitatea glenoid are o direcie oblic spre napoi i spre linia median. Diametrele cavitii glenoide sunt de: 20 mm anteroposterior, 20-25 mm n sens transversal, 6-7 mm adncime, deci diametrul transversal predomin n raport cu cel antero-posterior. Suprafaa osoas articular mandibular este situat pe faa anterioar a condilului mandibular. Condilul mandibular are o form elipsoidal (oblic napoi i nuntru ca i cavitatea gle-noid), are diametrul mare de 20-25 mm iar diametrul mic de 10 mm. Condilul mandibular prezint 2 versani: unul anterior convex, articular, acoperit cu un strat cartilaginos i unul posterior, extracapsular. Cartilajul de
acoperire a supra feei articulare are o grosime inegal, grosimea maxim fiind la mijlocul suprafeei articulare condiliene, iar prin elasticitatea sa protejeaz osul de ocuri, amortizndu-le (fig. 7). Cartilajul articular prezint cteva caracteristici eseniale: structura este specializat pentru micri la frecare joas; distribuie uniform la ncrcare i transmisie; amortizare controlat a ocurilor mecanice. Proprietile biomecanice ale cartilajului articular sunt: a. compresibilitatea asigur cartilajului rolul de amortizor pentru esutul osos subiacent care s-ar eroda prin frecare. Presiunile intermitente favorizeaz nutriia cartilajului; b. elasticitatea asigur capacitatea cartilajului de a se aplatiza sub efectul unei sarcini ca apoi s revin la grosimea normal dup ce sarcina a fost ndeprtat. Elasticitatea asigur circulaia lichidului sinovial ntre suprafeele
Fig. 7. Articulaia temporo-mandibular, seciune parasagital (dup Atkinson, 1992, modificat) Fig. 7. Articulaia temporo-mandibular, seciune parasagital (dup Atkinson, 1992, modificat) 1 2 3 4
3. Menisc 4. Cavirare glenoida 1. Conduct auditiv extern 6. M.pterigoidian lateral 2. Capsula posterioar
articulare, cartilagii, favoriznd astfel schimburile nutriionale (asemenea unei "pompe peristaltice"). Strile de deshidratare determin o micorare a elasticitii cartilajului, fiind una din cauzele artrozelor senile; c. pofozitatea: structura de "burete" permite mbibarea cartilajului articular cu lichid sinovial asigurndu-se astfel schimburile nutritive dar i amortizarea mecanic a ocurilor; d. rezistena la forele de tensiune i forfecare. Aceste proprieti au importan funcional n nutriia, metabolismul, activitatea static i dinamic articular. 2.1.2. Meniscul articular intercondilian (discul articular) Meniscul articular se afl situat ntre cele dou suprafee articulare (condilul temporal i condilul mandibular) avnd form de lentil biconcav cu marginile ngroate, are o structur fibrocartilaginoas. Meniscul separ cavitatea articular n dou compartimente, suprameniscal i inframeniscal, fiecare cu sinoviala lui (fig. 8), Micrile ce se efectueaz de ctre menise sunt complexe: n etajul inframeniscal se realizeaz micrile de rotaie, iar n etajul suprameniscal, micrile de translaie. n orice micare a mandibulei particip ambele ATM dreapt i stng. n cazul blocrii unei ATM prin anchiloz, ntreaga mandibul nu se va mica. Sistemele de arcade n fbrocartilagiul de pe suprafaa articular prezint o structur funcional:
partea articular menisco-temporal are o funcie de alunecare precis;
partea menisco-condilian realizeaz o micare precis de rotaie. n caz de solicitare defectuoas, aceste sisteme se reorienteaz.
2.1.3. Elementele de unire ale capetelor articulare Capsula ATM este un manon fibros cu aspect de trunchi de con ce are baza mare pe perimetrul cavitii glenoide, iar baza mic pe conturul suprafeei articulare condiliene. Aceast form se datorete mrimii diferite a suprafeelor articulare. Ligamentele ATM sunt reprezentate de ligamentele proprii intrinseci i ligamentele extrinseci (la distan). Ligamentul temporo-mandibular limiteaz micrile de retropulsie a mandibulei. Pe faa intern a articulaiei, n afar de ligamentul extern (slab dezvoltat), exist alte trei ligamente care limiteaz micrile de retropulsie i de lateralitate a mandibulei. Ligamentele stilomandibulare, sfeno-mandibulare i pterigo-mandibulare mpreun cu ligamentele temporo-mandibulare formeaz un aparat de suspensie amandibulei, care poate aluneca n jos, nainte sau lateral (fig. 9). Funciile ligamentelor sunt: ntresc capsula; previn micrile excesive ale condilului; previn dislocarea mandibule;
acioneaz ca receptori ai poziiei articulaiei. 2.1.4. Sinoviala ATM Sinoviala ATM este un esut conjunctivohistiocitar a crui strom prezint fibre de colagen orientate n sensul traciunilor mecanice, sinoviala tapetnd capsula articular. Funciile sinovialei: Fig. 9. Ligamentele ATM, aspect lateral (A) i medial (B) (dup Atkinson, 1992, modificat) este organul sensibil al articulaiei prin terminaiile sale, proprioceptive i dureroase; - are rol de aprare i protecie prin capacitatea macrofagic a celulelor din structura sa; - are rol de membran de filtrare prin care trec n lichidul sinovial substanele din snge (exemplu: proteine plasmatice, medicamente antiinflamatorii); are rol plastic, umplnd golurile dintre suprafeele articulare n repaus i spaiile interarticulare formate n timpul unor micri, permind acomodarea acestora la mobilizare; are capacitate de regenerare, refcndu-se n cteva luni dup extirpare; are rol secretor, formnd lichidul sinovial. Lichidul sinovial reprezint filmul lichidian vscos de pe suprafeele cartilajelor articulare. Se formeaz printr-un proces de ultrafiltrare local a plasmei sanguine. Lichidul sinovial are
urmtoarele funcii: lubrefiant, nutritiv pentru cartilajul articular poros, particip la clearance-ul unor substane ptrunse n lichidul sinovial. Sinoviala prin prelungirile sale n interiorul cavitii articulare i prin lichidul sinovial completeaz spaiile de incongruen static sau dinamic dintre suprafeele articulare. Particularitile ATM: este divizat n dou caviti articulare prin discul articular fibros sau menise; suprafeele articulare ale oaselor sunt acoperite de un fibro-cartilaj, n loc de cartilaj hialin ca la alte articulaii; cele dou ATM nu se pot mica independent una fa de alta: dac o articulaie temporo-mandibular face o micare, cealalt articulaie, obligator, trebuie s fac micarea compensatorie.
2.1.5. Inervaia ATM Inervaia ATM este asigurat de ramuri din nervul auriculo-temporal sau din nervul coarda timpanului. 2.1.6. Receptorii articulari ATM este o important zon reflexogen datorit inervaiei bogate, densitii i varietii receptorilor din aceast zon. Inervaia senzitiv bogat face ca ATM s fie un veritabil organ senzorial periferic. Astfel, se descriu: 2.1.6.1. Corpusculii Ruffini sunt localizai n capsula ATM, sunt constituii din terminaii mielinizate iar extremitatea ncapsulat este nemielinizat. Activarea receptorilor Ruffini depinde de poziia static sau dinamic a articulaiei, se face pe zone de control a poziiei. ntr-o anumit poziie articular ce produce o angulare ntre 15-30 este stimulat un receptor, iar cnd poziia articular se schimb intr n funcie alt receptor, iar primul trece n repaus. Fiecare receptor controleaz micarea n interiorul unghiului de 15-3G. Viteza micrii n interiorul acestui unghi determin inducerea de poteniale de aciune corespunztoare n receptor. Aceti receptori sunt considerai indicatori ai direciei de micare. 2.1.6.2. Organul lui Golgi pentru articulaii este situat n ligamentele articulaiei i este asemntor corpusculilor tendinoi Golgi. Sunt sensibili la angulari cu limite mai largi, sunt indicatori de poziie i micare. Corpusculii Vater-Paccini prezint central o fibra mielinizat, sunt receptori ai micrilor rapide. Clasificarea ntorfo-fiziologic a receptorilor ATM Din punct de vedere morfologic i electro-fiziologic receptorii senzoriali ai ATM se pot mpri n patru categorii (tabelul 2): a. Mecano-receptori articulari de tip I sunt localizai n straturile superficiale ale capsulei ATM, au forma unor corpusculi globuloi ncapsulai, cu diametrul de 100/40 micrometri, ndeplinesc funcia de mecano-receptor ai poziiilor statice i dinamice ale ATM.
b. Receptorii articulari de tip II sunt localizai n straturile profunde ale capsulei i n esutul celulo-grsos localizat pe faa posterioar a capsulei ATM, cuprind corpusculi ncapsulai cu dimensiuni de 280/120 micrometri, n grupuri de 2-4 corpusculi. Au funcie de mecano-receptori dinamici, se adapteaz rapid, au prag de sensibilitate sczut. c. Mecano-receptori de tip III cuprind corpusculi fuziformi ncapsulai, cu dimensiuni de 600/100 micrometri localizai n ligamentele intrinseci i extrin seci ale ATM. Sunt receptori dinamici, cu prag de sensibilitate ridicat i cu adaptare foarte lent. d. Receptor de tip IV sunt reprezentai de terminaii nervoase libere, ce formeaz plexuri localizate difuz: capsula articular, ligamente, esut celulogrsos periarticular, pereii vaselor sanguine. Acetia au rol de receptori ai durerii (algoceptori), nu se adapteaz, au un grad nalt de sensibilitate. Funciile receptorilor articulari sunt complexe: sesizeaz poziia articulaiei n repaus sau n micare, intervin n dirijarea i coordonarea reflex a diferitelor grupe musculare responsabile de mobilitatea mandibulei. n principal receptorii de tip I, II sunt senzori de informare i reglare n condiiile funcionrii normale ale articulaiei, iar receptorii de tip III i IV intervin n situaiile ce depesc parametrii funcionali normali ai ATM. De asemenea, au rol n transmiterea impulsurilor nociceptive. Senzaia de durere n ATM poate inhiba puternic micrile condilului. Datorit configuraiei suprafeelor articulare i a osului din jur, micrile ATM sunt limitate n plan sagital i coronal. Capul condilului poate aluneca n fa sau n spate n plan sagital, sau se poate rota n plan coronal. Micrile laterale ale mandibulei sunt produse prin micri de alunecare (glisare) ale articulaiilor
n acelai plan sagital cu un condil ce se mic n fa iar cellalt face o micare compensatorie de rotaie n jurul axei verticale.
ATM este cea mai complex articulaie din organismul uman. La mamifere, micarea condilului se face cu predominan sagital (propulsie, retropulsie) numai n groapa mandibular, deplasrile fiind ngrdite de proieminenele de pe marginile cavitii glenoide. La om, apare micarea de translaie a condilului, care pare a fi o consecin a staiunii bipede. n micrile uzuale, maxilarele funcioneaz ca o prghie de gradul III la care: ATM este punctul fix, muchii maseter, pterigoidian intern i temporali creeaz fora, dinii reprezint organul efector prin care se transmite fora la obsta colul alimentar. Spre incisivi, prgia este mai puin eficace iar dinii sunt solicitai mai puin. n laterotruzie dreapt condilul activ blocat n cavitatea glenoid devine punctul fix, iar condilul stng este de balans. Dac n aceast micare, dinii de pe partea stng pstreaz un contact prematur, la nivelul contactului se va situa punctul de rezisten. Fora produs de muchii ridictori de pe partea stng va transforma prghia de gradul III ntr-o prghie de gradul II, mai eficace funcional dar mai traumatizant. Dac contactul de pe partea stng este de mai mare importan, el devine punctul de sprijin, iar prghia se transform ntr-o prghie de gradul I, avnd ca rezultat apariia unor fore foarte puternice la nivelul dintelui de contact, fore care se pot repercuta i asupra ATM respectiv asupra muchilor n unele micri de protruzie dac exist un contact molar, acesta va deveni punct fix. n micarea de incizie fora va aciona posterior punctului fix, iar rezistena la nivel incisiv, formndu-se o prghie de gradul I cu eficacitate maxim care ns suprasolicit att molarul cu contact prematur ct i incisivii. Rezultatele suprasolicitrii sunt: uzur sau mobilitate a dintelui, March Robnson susine c ATM nu sufer nici un efort n funcionalitatea ei normal, iar mandibula nu poate fi asimilat unei prghii. Robinson descrie un mecanism reflex care previne orice efort, n limitele normalului, dirijat spre ATM. Fora ia natere n timpul contraciei ridictorilor i se dirijeaz spre ATM. n acest moment se declaneaz contracia reflex a muchilor temporali, a cror for trage mandibula n sus i posterior, anihilnd ocul. Dup alte teorii, efortul se distribuie att la nivelul dinilor ct i la nivelul ATM. n fiziologia ATM un rol fundamental l joac meniscul, mpreun cu muchiul pterigoidian, prin cele dou fascicule ale sale cu inserie pe menise i pe condilul mandibular. Meniscectomia la om duce la pierderea micrilor de propulsie. n repaus lama meniscal se muleaz n cavitatea glenoid pe versantul articular al condilului temporal, sub influena ascensiunii condilului mandibular. Plecnd de Ia aceast poziie, iniial condilul nu poate efectua dect micri de
2.2. DINAMICA ATM
rotaie. Micarea de propulsie este precedat de contracia fasciculului superior al pterigoidianului extern, care pune n tensiune lama meniscal ce se aeaz ca un plan nclinat ntre scizura Glasser i vrful condilului temporal, determinnd n acelai timp o uoar dezanclavare a condilului din glena ctre antero-inferion ntinderea lamei meniscale se datorete elasticitii sale mai ales n poriunea retrocondilian. Sub aciunea continu a contraciei fasciculului superior al pterigoidianului extern, meniscul temporo-mandibular ntins gliseaz spre anterior, dnd posibilitatea condilului mandibular s alunece pn la vrful condilului temporal, alunecare ce se produce sub aciunea contraciei fasciculului inferior al pterigoi-dianului extern. Limitarea micrii anterioare a meniscului se datorete frului posterior al meniscului. Micarea invers, de revenire n glen, se face prin ncetarea mai nti a contraciei fasciculului inferior, condilul revenind n glen, sub aciunea elevatorilor i prin ncetarea contraciei fasciculului superior. Funcia principal a complexului menisco-pterigoidian este de a dezan-clava condilul din glen i de a-i oferi planul nclinat pentru glisare. La pacientul sub narcoz, deschiderea gurii prin forare, se face printr-o rotaie cu condilii n glen, urmat de ieirea brusc a condililor din cavitatea glenoid i deschiderea maxim a gurii. Studii EMG au artat ca fasciculul superior al p teri goidianul ui extern se contract naintea celui inferior n micarea de propulsie i deschidere. Pterigoidianul extern particip nu numai la protracii i diducii ci i n micrile de ridicare a mandibulei. Revenirea meniscului n glen este o micare pasiv, determinat de revenirea condilului sub aciunea elevatorilor i de elasticitatea frului meniscal posterior. n aceste condiii, contracia uoar a fasciculului pte-rigoidian superior are rolul de a preveni o revenire brusc a meniscului n glen sub aciunea elevatorilor i elasticitii frului meniscal posterior Aceste elemente de fiziologie menisco-pterigoidian explic producerea zgomotelor articulare, patogenia luxaiilor temporo-mandibulare, apariia disfun-ciilor temporo-mandibulare n condiia unor mici tulburri de ocluzie. Deplasrile mandibulei sunt necesare n toate funciile sistemului oro-faciat. n timpul masticaiei, daca aceasta se face bilateral, mandibula poate fi considerat c acioneaz ca o prghie de gradul III (punctul de sprijin n condil, fora la mijloc, Ia nivelul gonionului i apofzei coronoide, iar rezistena la nivelul arcadei dentare, deci la cealalt extremitate a prghiei). n aceste condiii se exercit o for puternic la nivelul articulaiei temporo-mandibulare. Cnd masticaia se face unilateral, condilul de partea activ este mai puin solicitat, deoarece particip n cealalt jumtate a mandibulei (de balans n aceast micare) i mandibula este transformat ntr-o prghie de gradul II deoarece se adaug i fora muscular de pe partea de balans, care pe lng faptul c mrete fora, reduce i sarcina condilului de partea opus. Cnd mandibula acioneaz ca o prghie de gradul III, forele sunt egal repartizate de fiecare parte. Cu ct rezistena alimentar este mai posterioar, cu att presiunea exercitat la nivelul condililor este mai mare. n activitatea unilateral a mandibulei (prghie de gradul II), bolul alimentar deplasndu-se dinainte-napoi, presiunea se reduce la condilul de partea activ,
putnd deveni chiar negativ (traciunea pe capsul n loc de presiune), n timp ce presiunea la condil de partea de balans rmne nalt. Deci: ATM - este sediul de recepionare i transmitere ctre scheletul cranian al unor fore puternice. Teoria lui Marsh Robinson susine c la nivelul ATM nu se exercit nici o for. Toate forele se transmit n regiunea arcadelor dentare. Deplasrile mandibulei se fac prin echilibrul forelor musculare antagoniste i protagoniste, realizat prin jocul controlat al reflexelor nervoase, att n poziia de repaus a mandibulei ct i n diversele ei micri. In teoria Robinson nu se ia n considerare rolul pterigoidianului extern. Repciuc, consider ATM ca o articulaie de conducere muscular iar zonele articulare influeneaz elementele micrii. In cursul micrilor fiziologice nu este posibil o pierdere de contact a suprafeelor articulare, vidul articular, datorit presiunii atmosferice asigur deplasarea elementului osos n articulaie dup terminarea micrii. Studii de mecanic muscular arat c presiunea condilului la nivelul cavitii glenoide nu poate fi important. ns, n poziie de ocluzie central, analizele de mecanic muscular au artat c iau natere fore ce sunt ndreptate spre ATM, iar condilul apas mai puternic pe rebordul posterior al fosei articulare care este ngroat, n general, toat circumferina fosei articulare este supus la solicitri, ce pot modela condilul. Studii complexe ale funcionalitii ATM arat c: presiunea cea mai mare este absorbit la nivelul arcadei dentare; o parte din fore pot fi transmise i de-a lungul osului mandibular spre ATM; alte fore se propag prin fasciile musculare, n special fascia muchiului temporal. Conformaia ATM prezint modificri succesive n raport cu vrsta, modificri ce sunt ntr-o strns corelaie cu fenomenele dentiiei, ATM ar fi complet format la 25 de ani. Erupia dinilor temporari se nsoete de modificri n jocul muscular cu instalarea unor noi reflexe. Cu vrsta, au loc dezvoltarea tuberculului articular i a capului condilului. La nivelul ATM pot avea loc modelri funcionale ce includ conformaia cavifii glenoide i a tuberculului articular, dezvoltarea i conformaia condilului mandibular. Tratamentele cu aparate ortodontice pot determina modificri Ia nivelul ATM; resorbii pe suprafeele de contact cu presiune; apoziie compensatoare pe versantele opuse presiunii; restructurarea condilului ca i a restului osului mandibular; modificri ale cavitii glenoide. 2,2.1- Micrile ATM pentru deschiderea gurii In micarea de deschidere a gurii, condilul se deplaseaz nainte mpreun cu meniscul articular, fcnd i o uoar micare de rotaie. Aceast deplasare este efectuat de muchii pterigoidieni externi, prin contracia bilateral: fasciculul superior al muchiului pterigoidian extern deplaseaz meniscul nainte, iar fasciculul inferior deplaseaz condilul. Meniscut alunec pn pe faa inferioar a tuberculului articular unde formez o "glenoid transportabil
i deformabiia" condiiului articular (Rainer) aceasta pentru a rspunde diferenelor de presiune a diferitelor momente mecanice din articulaie. Deschiderea gurii se face n trei etape: Etapa I: const din realizarea unei micri de rotaie a condiiului pe menise n jurul unui ax transversal ce trece prin centrul condililor i corespunde cu inseria inferioar a ligamentului temporomandibular. Caracteristicele acestei etape sunt: micarea este de tip "balama"; deschide gura cu aproximaxiv 2-4 mm; corespunde trecerii de la ocluzia central la poziia de repaus a mandibulei; micarea este ghidat de receptorii din ligamentele interne i externe (temporo-mandibular) ale articulaiei; limitarea micrii se realizeaz de ctre ligamentele pterigomandibulare i sfenomandibuiare. Etapa II: const din efectuarea a dou micri n ATM: una de rotaie condilomeniscal i alta de translaie temporo-meniscal. Caracteristicile acestei etape sunt: nsumarea celor dou micri (rotaie, translaie) determin deschiderea gurii cu 4 centimetri; rotaia condiiului este asigurat de muchii suprahioidieni i pieloi; micarea de translaie este realizat de contracia simetric a muchilor pterigoidieni externi; micrile se realizeaz n jurul unui ax transversal care alunec i el n jos i nainte. Etapa III: se efectueaz suplimentar i voluntar. La sfritul etapei Ii, muchii maseteri i pterigoidieni interni sunt ntini la maximum iar fibrele lor devin paralele cu ramura ascendent a mandibulei, n momentul cnd aceti muchi se relaxeaz brusc, se contract muchii pterigoidieni externi care prin fascicolele inferioare trag napoi de braul mic al prghiei mandibulare iar fascicolele superioare trag meniscul nainte- Consecinele acestor contracii sunt: se produce micarea de translaie nainte a meniscului i micarea de rotaie napoi a condiiului; gura se deschide cu nc 2 centimetri (n total, 6 centimetri). Micarea de deschidere a gurii n trei etape este o micare continu, iar mentonul descrie un arc de cerc cu concavitatea napoi-Disfunciile ATM se caracterizeaz prin: etapa a IlI-a se face sacadat, sau "n baionet"; leziuni aie meniscului; capul condiiului alunec naintea bureletului meniscal; dureri pterigoidiene, crepitaii, cracmente. 2.2.2. Micrile ATM pentru ridicarea mandibulei Micarea de ridicare a mandibulei se face printr-un mecanism invers: relaxarea brusc a muchilor pterigoidieni externi; contracia progresiv a m. maseter, m. pterigoidieni interni i m.temporali; alunecarea meniscului n groapa glenoid;
rotaia i translarea invers a capului condilului; arcadele dentare vin, din nou, n contact; gura se nchide; presiunea produs pe dini se transmite prin sistemele traiectoriale, la mandibul sau masivul facial. 2.2.3. Micrile ATM pentru propulsia mandibulei Micarea de propulsie a mandibulei se realizeaz prin translaia meniscului i condilului secundar contraciei bilaterale a m. pterigoidieni, m. accesoriu i a fascicolului superficial al maseterului. Micarea de propulsie se nsoete obligatoriu de o micare de coborre a mandibulei. Amplitudinea micrii de coborre a mandibulei depinde de: adncimea glenoidei, nlimea cuspizilor dentari, gradul de supraocluzie incizal, planul de ocluzie. In propulsia maxim (de 2-2,5 cm) versantul anterior al condilului articular apas pe menise i pe tuberculul articular. Aceste presiuni stimuleaz dezvoltarea sagital a mandibulei pn la nchiderea cartilajului de cretere. Dup nchiderea cartilajului de cretere, se pot produce modificri temporare prin propulsia forat cuspidian sau propulsia prin aparate ortodontice. n propulsia mandibulei se realizeaz o cretere a procesului alveolar care are ca rezultat reechilibrarea ocluziei. Micarea de propulsie este limitat de ligamentele stilo-mandibular i temporo-mandibular. Micarea de propulsie a mandibulei prezint variaii n funcie de vrst: la copil, alunecarea meniscului i a condilului se face aproape orizontal, uneori fr s se nsoeasc de coborrea mandibulei (daca dinii sunt uzai); dup vrsta de 7 ani propulsia se asociaz cu o coborre a mandibulei, pentru c se formeaz versantul condilian articular, tuberculul articular i se definitiveaz supraocluzia frontal. Planul de alunecare al meniscului i al condilului face cu planul ocluzal un unghi variabil, ntre 40o-45, 2,2,4. ATM i retropulsia mandibulei Retropulsia mandibulei este realizat de contracia fascicolelor posterioare, mijlocii ale temporalului, de m. digastric, m. geniohioidieni i accesoriu de m. milohioidieni. Alunecarea napoi a meniscului i condilului se face de-a lungul unui plan orizontal sau oblic care face un unghi de 40-45 cu planul ocluzal. Cnd alunecarea condilului depete limita posterioar a capsulei articulare, versantul posterior al condilului comprim esuturile retrocondiliene, esuturi care posed o bogat inervaie vegetativ. 2,2,5. ATM i micrile de lateralitate Micrile de lateralitate ale mandibulei se realizeaz prin alunecarea condiluui i a meniscului de o parte (n urma contraciei m. pterigoidian extern de aceiai parte, n principal i accesoriu, a m. pterigoidian intern, m. maseter, fascicol anterior ale m. temporal) iar condilul de partea opus rmne pe loc sau alunec puin napoi (prin contracia fascicolelor posterioare ale m, temporal). M. temporal are rol principal n micarea de lateralitate a mandibulei. In cazul n care mentorul! sau linia interincisiv se deplaseaz spre dreapta, condilul i meniscul de partea stng se deplaseaz nainte, n jos i nuntru,
formnd cu planul medio-sagital un unghi de 15. Condilul de partea dreapt st pe loc sau se deplaseaz foarte puin napoi i nafar i reprezint axul vertical n jurul cruia se face aceast micare de lateralitate spre dreapta. Urmeaz apoi, micarea de lateralitate spre stnga, cnd condilul din stnga joac rol de pivot, n micarea de lateralitate, linia interincisiv se deplaseaz spre dreapta i spre stnga, n interiorul unui unghi de aproximativ 122. Micrile de lateralitate se asociaz cu micri de propulsie, retropulsie, coborre sau ridicare, micri complexe ce apar, de obicei, n masticaie. Amplitudinea micrilor de lateralitate scade cu deschiderea gurii sau cu retropulsia mandibulei. Micrile i forma ATM sunt influenate de vrst, tipul dentiiei, tipul dinamic masticator: la copil predomin micrile de propulsie - retropulsie pn la apariia dinilor frontali permaneni, cnd apare supraocluzia incisiv; n raport cu tipul dinamic masticator se modeleaz corespunztor forma condilului, a cavitii glenoide i se definitiveaz tipul de uzur dentar; supraocluzia incisiv i cuspizii nali favorizeaz apariia unui tip dinamic masticator, de "toctor" la care predomin micrile de ridicare i cobo rre a mandibulei. Consecinele acestor micri sunt: adncirea fosei glenoide, accentuarea pantei versantelor condiliene; cuspizii dentari teri, unii prin creste, favorizeaz predominena micrilor de lateralitate i definitivarea tipului masticator de "frector"; la tipul masticator "propulsor", predomin micrile de propulsie retropulsie, ocluzia realizndu-se cap la cap. n acest caz pantele condiliene i dentare sunt foarte line sau terse complet.
ATM este influenat de modificarea tiparelor de micare articular i micro traumatismele ce apar n cazul unor disfuncii ce au punctul de plecare parodontal, odontal, neuromuscular. Simptomatologia Ia nivelul ATM poate cuprinde: DUREREA ARTICULAR este de intensitate variabil (dureri surde, vagi, junghiuri), apare n repaus, poate fi exacerbat la micare. Durerea pe care pacientul o acuz n ATM poate fi: durere local (datorit patologiei locale: traumatisme, infecii, neoplasme etc), durerea referit (durerea apare n ATM dar este produs de procese patologice n alte zone: ureche, parotid, muchi), durerea psihogen. ZGOMOTELE ARTICULARE pot fi: cracmentul, crepitaiile. Cracmentul este un zgomot unic, puternic, uni sau bilateral, datorat artroliilor, osteoftelor, deformrilor degenerative ale meniscului. Crepitaiile sunt zgomote multiple de intensitate mai mic. Mecanismele principale de producere a zgomotelor articulare sunt: hiperlaxitatea ligamentar i capsular; oboseala muchilor, care permit micri largi; dezechilibre musculare, n special ale pterigoidianului extern.
2.3. DISFUNCIILE ATM
SALTUL ARTICULAR reprezint micarea sacadat uni- sau bilateral, ce deranjeaz pacientul care se va adresa timpuriu medicului. DEVIEREA MANDIBULEI se nsoete de creterea amplitudinii deschiderii gurii, mandibula fcnd micri de zig-zag, brusc sau lent. SUBLUXAIA se datorete creterii laxitii elementelor periarticulare, unisau bilateral, de obicei este reductibil prin manevre executate de pacient. LIMITAREA DESCHIDERII GURII se produce datorit fibrozrii i sclerozrii esuturilor periarticulare, poziiilor antalgice, spasmului muscular. BLOCAJUL ARTICULAR se produce datorit artroliilor, osteofite fracturate, menise plicaturat, spasm muscular mergnd pn la trismus.
MICRILE MANDIBULARE
Dinamica matidibular poate fi sistematizat dup numeroase criterii. Astfel micrile mandibulei pot fi clasificate n: micri mandibulare elementare: micri simetrice pure realizate fr contact interdentar ce cuprind: micri de coborre, ridicare, propulsie, retropulsie, retracie forat; micri efectuate cu contact interdentar dar fr interpoziie de alimente; micri de masticaie cu dini naturali; micri de masticaie cu dini artificiali. O alt clasificare a micrilor mandibulei este: micri limit n plan frontal, sagital, orizontal; micri funcionale ce cuprind: micri de masticaie, de deglutiie, de fonaie, etc. Dup Posselt micrile mandibulare de baz sunt: micrile de deschidere i nchidere; micarea de protruzie cu contact dentar i revenire; micarea de retruzie i revenire, cu contact dentar; micarea de lateralitate i revenire, pstrnd contactul dentar.
Micrile fundamentale (micri pure) ale mandibulei sunt: micarea de rotaie i micarea de translaie, ce se pot realiza n cele trei planuri: sagital, orizontal i frontal. Micarea de rotaie se realizeaz cnd fiecare punct al mandibulei parcurge o traiectorie curba a crei lungime crete proporional cu distana de la centrul
3.1. MICRILE FUNDAMENTALE ALE MANDIBULEI
de rotaie la un punct luat n considerare, Micarea de translaie se realizeaz cnd fiecare punct al mandibulei se va deplasa pe o traiectorie aproximativ liniar, parcurgnd distane egale faa de punctul de origine.
n dinamica funcional mandibulara numai micrile de balama i mi. carea de propulsie se ntlnesc ca micri pure, celelalte tipuri de micri fiind ncadrate n micrile combinate pe care Ie execut mandibula. Micrile mandibulei se coreleaz cu micrile n articulaia temporomandibular care realizeaz att micri pure (de rotaie i translaie) ct i micri combinate. Micrile mandibulare sunt oglinda configuraiei anatomice i funcionale a determinanilor anatomici posteriori (ATM), anatomici anteriori (ocluzia), funcionali (muchii mobilizatori ai mandibulei). 3.2.1, nregistrarea micrilor mandibulei Micrile principale ale mandibulei (coborre, ridicare, antero-posterioare, de lateralitate dreapta-stnga) se integreaz armonios n funcia masticatorie, conform unui tipar individual, specific fiecrui individ. Aparatura de nregistrare a micrilor mandibulei trebuie s ndeplineasc unele condiii: aparatura s nu deranjeze i s nu modifice micrile mandibulei; sa redea cu acuratee datele i sub o form compatibil analizei computerizate; nregistrrile s fie complete i s redea ntreaga desfurare a procesului; datele culese s fie prezentate sub o form accesibil interpretrii clinicienilor. Dintre metodele multiple de nregistrare amintim cele ce folosesc: traductoare mecanoelectrice fixate prin gutiere pe arcadele dentare; traductoare electronice cuplate cu fotocelule legate de incisivii inferiori; filmarea micrilor mandibulei; traductori feromagnetici cu magnet ataat incisivilor inferiori sau n vestibulul labial mandibular, etc. Indiferent de modalitatea de nregistrare se impun cteva remarci: masticaia urmeaz un traseu ciclic, n direcie predominant vertical-oblic; exist o component de lateralitate transversal i oblic, nsoit de glisare la nivelul ocluziei; deci, se realizeaz micri ciclice n toate cele 3 planuri sagital, frontal,orizontal. 3.2.1.1. Anvelopa micrilor limit n plan sagitat Anvelopa micrilor limit reprezint traseul ce delimiteaz o figur geometric n interiorul creia se nscriu micrile funcionale
3.2. MICRILE MANDIBULARE COMBINATE
mandibulare.Anvelopa micrilor limit n plan sagital cuprinde: coborrea, ridicarea mandibulei; deschiderea, nchiderea gurii; propulsia mandibulei. Cu ajutorul unui traductor plasat la nivelul punctului incisiv inferior se nscrie n plan sagital diagrama POSSELT pe care se descriu urmtoarele poziii succesive (fig- 10): poziia de intercuspidare maxim (IM); poziia de propulsie cu dinii frontali (incisivii) cap la cap (PP); poziia de propulsie maxim (PPM); traiectoria de deschidere din propulsie maxima (PPM-M); poziia de deschidere maxim (M) cea mai anterior posibil; poziia de nchidere cea mai posterior posibil (M-H-IM); h=traiectoria de nchidere (deschidere) obinuit. Anvelopele micrilor funcionale n plan sagital: se ncadreaz n anvelopa micrilor limit; ciclurile masticatorii au la nceput amplitudini mai mari, care se reduc pe msura ce alimentul este frmiat i amestecat cu saliva; ridicarea mandibulei este mai rapid i nu urmeaz ntotdeaunatraiectoria coborrii.
3.2.1.2. Anvelopa micrilor limit n plan frontal; cuprinde deschiderea maxim a gurii cu deplasarea extrema de lateralitate dreapta-stnga a mandibulei. Imaginea se aseamn cu un "scut" "pescru"; ciclurile funcionale se nscriu n interiorul anvelopei micrilor limit. 3.2.1.3. Anvelopa micrilor limit n plan orizontal: realizeaz nregistrarea micrilor laterale extreme, dreapta i stnga. Aspectul curbei este de romb. 3.2.1.4. Procesul masticator Undele masticatorii sunt compuse din: panta descendent - coborrea mandibulei; -vrful undei; panta ascendent - ridicarea mandibulei; palierul ocluzal - linie dreapta, rotunjit, cu denivelri, corespunde contactului ocluzal. Procesul masticator reprezint timpul necesar masticaiei unui aliment de la introducerea lui n cavitatea bucal, pn la deglutiie. Procesul masticator are urmtoarele faze: Faza I: - corespunde poziiei de ocluzie; se nregistreaz ca o linie dreapt. Faza II: - introducerea alimentului n gur se nscrie ca o pant descendent, de amplitudine i nclinaie variabil n funcie de:
deschiderea mai mare sau mai mic a gurii; i viteza de coborre a mandibulei. Faza III: - corespunde adaptrii masticaiei la consistena alimentului; se nregistreaz unde cu paliere ocluzale mai lungi; Faza IV: - "faza de regim a masticaiei", cnd eficiena funcional este maxim, are loc o stabilizare a amplitudinii i frecvenei undelor; este faza care poate caracteriza un anumit individ i un anumit tip masticator; Faza V: - de formare a bolului; se nregistreaz unde cu pante mai line i cu pauze mai mari ntre ele; deglutiia se nregistreaz sub forma unei linii orizontale.Pentru nregistrarea unui proces masticator n Laboratorul de Fiziologie UMF Timioara s-a folosit un sistem mecano-electric (imaginat de Dr. Goia Smaranda) ce cuprinde: dispozitivul de susinere a capului; 2 traductori de deplasare conectai de menton; un sistem de nregistrare: inscriptor XY Sensibilitatea sistemului de nregistrare se poate modifica dup necesiti. Pentru nregistrarea unui proces masticator s-au folosit biscui mici, n greutate medie de 3-5g. Sensibilitatea inscriptorului a fost de IV/cm, iar viteza de nregistrare de 2s/cm. nregistrrile s-au efectuat la subieci normali, de ambele sexe, tineri cu vrsta cuprins ntre 19-20 ani (fig. 11; 12), 3.2.2. Micrile funcionale ale mandibulei Micrile funcionale reprezint combinaii ale micrilor fundamentale, realizndu-se concomitent n mai multe planuri. Printre principalele micri funcionale amintim; micrile de masticaies deglutiie, supt, fonaie, vorbire, etc.
Fig. 11, Procesul masticator Ia 1; 2; 3; 4 biscuii, nregistrat la o persoan de sex masculin,19 ani
Fig. 12. Procesul masticator Ia 1; 2 biscuii, nregistrat la o persoan de sex feminin, 19 ani (diagram original din colecia Dr. Goia Smarauda, micorat).
FIZIOLOGIA MASTICAIEI
Masticaia reprezint procesul de prelucrare mecanic a alimentelor, de
insalivare, proces finalizat n formarea bolului alimentar. In efectuarea micrilor masticatorii, punctul de plecare pentru deplasrile mandibulare este dat de poziia de repaus. Mandibula este meninut n poziia de repaus de ctre: echilibrul tonic dintre grupele antagoniste ale muchilor mobilizatori i aciunea gravitaiei. Micrile efectuate prin contracia muchilor mobilizatori ai mandibulei sunt complexe. Aceste micri se fac n vederea prehensiunii, tierii sau sfierii alimentelor, n scopul zdrobirii i triturrii lor.
Fazele mecanice ale masticaiei sunt: incizia alimentelor, sfrmarea i triturarea. Aceste etape sunt precedate de acte motorii ajuttoare: echilibrarea capului i prehensiunea alimentelor. 4.1.1. Echilibrarea capului Echilibrarea capului reprezint prima micare controlat i continu, pregtitoare a masticaiei. Muchii flexori ai capului se contract iar n corelaie cu extensorii menin capul n poziia optim. 4.1.2. Prehensiunea alimentelor Prehensiunea alimentelor se realizeaz n doi timpi prin: - coborrea mandibulei cu deschiderea voluntara a gurii, proces realizat de contracia controlat de ctre sistemul nervos central a m. suprahioidieni, m.subhioidieni i pterigoidieni externi (fascicol inferior) (fig. 13). Coborrea mandibulei este favorizat la om de fora gravitaional. In micarea de coborre a mandibulei, condilul mandibular face o micare de translaie nainte i n jos (tracionat de m. pterigoidian extern) odat cu meniscul interarticular. Hioidul coboar fiind mobilizat napoi de m. subhioidieni. n acest timp m. ridictori ai mandibulei (maseter, pterigoidian intern, fascicolul mijlociu al temporalului) prezint o relaxare controlat, astfel nct coborrea mandibulei se face lent, dirijat i continuu. - propulsia mandibulei efectuat de contracia bilateral i simetric a muchilor pterigoidieni externi i a fascicolelor anterioare ale temporalului, asociat cu relaxarea controlat a muchilor retropulsori (m. suprahioidieni, fascicolul posterior al temporalului).
4.1. FAZELE MASTICAIEI
4.1.3. Secionarea alimentelor (incizia) Ridicarea mandibulei ncheie etapa de prehensiune i alimentele aduse cu mna spre gur sunt secionate de dinii frontali. Are loc contracia puternic a fascicolului mijlociu al temporalului, a muchilor maseteri i a pteri-goidienilor interni rezultnd nchiderea gurii n mod lent, continuu i puternic. Hioidul este mobilizat nainte i n sus de m. suprahioidieni (fig. 14). Micarea de incizie este unic. M. pterigoidieni externi, prezint o contracie lent pentru a imprima mandibulei, n ridicare, o poziie optim pentru articularea cu maxilarul superior, poziia de cap la cap a incisivilor superiori i inferiori. In timpul inciziei, incisivii inferiori alunec pe faa palatinal a celor superiori. Incizia solicit incisivii la basculare n jurul punctului de rotaie, micare creia i se opune rezistena osului alveolar, muchilor buzelor i limbii. Alimentele dure opun rezisten la nchiderea gurii, ceea ce determin o contracie mai mare a m. pterigoidian extern, m. supra-, subhioidieni, a muchilor mimicii i chiar a muchilor cefei. Astfel se echilibreaz mai bine capul i se realizeaz micrile ajuttoare pentru nfrngerea rezistenei alimentelor. Traiectoriile funcionale n incizie variaz dup consistena alimentelor. Pentru alimentele dure este necesar secionarea n poziia cap la cap a incisivilor. Pentru unele alimente, actul de incizie poate fi asociat cu o traciune cu mna a fragmentului exobucal, ce poate aciona ca o tor de basculare vestibular suplimentar. n timpul inciziei exist o mic "dezocluzie ' molar, care are rol n protecia parodontala.
In timpul a micrii de ridicare mandibulei, contracia muchilor ridictori este de tip izotonic. Cnd ridicarea mandibulei este'oprit de obstacolul alimentar, contracia devine izometric, realizeaz o tensiune muscular crescut, suficient pentru a nvinge rezistena alimentului. Canini sunt folosii la sfierea alimentelor mai consistente. 4.1.4. Zdrobirea i triturarea alimentelor Premolarii servesc la zdrobirea alimentelor. Ei prezint cuspizi ascuii, care le faciliteaz ptrunderea n interiorul alimentelor, pentru a le fragmenta n particole relativ mari. Molarii, prin suprafaa ocluzal mare, au rol important n frmiarea n continuare a alimentelor, n triturarea lor, n special, prin micri de late'ralitate care se succed spre dreapta sau spre stnga. Se realizeaz contracii alternative ale muchilor pterigoidieni externi de o parte i a temporalilor de cealalt parte, asociate cu relaxarea controlat a acelorai muchi, care devin antagoniti succesivi. De asemenea, are loc i o deplasare nainte i n jos a meniscului interarticular de partea opus micrii, astfel nct spaiul retrocondilian se mrete alternativ cu fiecare micare de acest fel. n timpul masticaiei: se dezvolt for n muchii ridictori ai mandibulei; se realizeaz contracia de balansare a muchilor direcionali; se produce relaxarea controlat a antagonitilor. n timpul micrilor de mcinare alimentele sunt aezate alternativ ntre arcadele dentare n sectoarele laterale. Alimentele sunt reaezate ntre arcadele dentare prin aciunea coordonat a limbii i obrajilor. Limba, datorit terminaiilor sale senzoriale, separ din masa alimentar acele fragmente care sunt bine triturate i insalivate, le adun la un loc formnd bolul alimentar pe care-1 pregtete pentru deglutiie. Pe msur ce alimentele sunt triturate are loc i amestecarea lor cu saliv, proces denumit insalivare. Limba particip la masticaie i prin frmiarea unor alimente, pe care le preseaz pe palatul dur. Limba execut micri de intensiti diferite: micri fine: de aezare a alimentelor ntre arcadele dentare, de triere a fragmentelor, de insalivare, de realizare a bolului alimentar; micri puternice: de sfrmare direct a alimentelor, de deglutiie, de curire a arcadelor dentare dup deglutiie. Bolta palatin particip la zdrobirea unor alimente mai mari i furnizeaz informaii asupra consistenei alimentelor. Obrazul, prin muchii buccinatori, readuce, pentru triturare, alimentele czute n vestibul. Buzele, mucoasa obrajilor, limba, palatul conin numeroi receptori senzitivi ce au rol n coordonarea dinamicii masticatorii. Buzele menin saliva i alimentele n cavitatea bucal n timpul triturrii i apreciaz temperatura alimentelor.
4.2. FORE MASTICATORII
Fora de contracie a muchilor masticatori este n funcie de gradul de dezvoltare al muchilor care depinde de: ras; regional: la eschimoi, care se hrnesc cu alimente crude, la nivelul tip molarilor forele dezvoltate sunt de trei ori mai mari dect la subiecii ce folosesc o alimentaie preparat; vrst; sex; modul de pregtire al alimentelor Contracia muchilor maseteri poate fi de tip izotonic sau de tip izo-metric. Contracia izotonic realizeaz scurtarea lungimii muchiului, cu meninerea constant a tensiunii. Prin contracie izotonic se pun n contact static suprafeele triturante ale dinilor anta-goniti. Se realizeaz prin contracia m. ridictori ai mandibulei i relaxarea m. cobortori. Contracia izometric se caracterizeaz prin meninerea constant a lungimii muchiului i creterea tensiunii dezvoltat n muchi (fig. 15). Contracia izometric se realizeaz dup "prinderea" alimentelor ntre suprafeele ocluzale i dureaz pn cnd tensiunea dezvoltat n muchi nvinge rezistena (duritatea) alimentelor, n acest moment alimentul este zdrobit iar contracia izometric este urmat de contracia izotonic. Deci, n timpul masticaiei au loc schimbri permanente ale tipului de contracie. n urma contraciei n muchii masticatori se dezvolt o for mecanic, proporional cu numrul de uniti motorii antrenate funcional. Fora de contracie depinde de gradul de ntindere al muchiului, pn la un moment dat. Dac distana iniial dintre dini este mai mare (maseterul este mai ntins) fora de contracie este mare. Aceasta se dato-rete, pe de o parte, stimulrii fusurilor neuro-musculare, iar pe de alt parte, posibilitii de a se realiza un numr maxim de puni acto-miozinice. Lungimea optim a sarcomerului la care fora dezvoltat este maxim se
consider a fi de 2,2 micrometri. Dac distana dintre mandibul i maxilar este prea mic, musculatura nu poate aciona la lungimea optim, iar fora de contracie scade (numrul de puni acto-miozinice active este sczut, se produce o nclecare a filamentelor de actin). Viteza de scurtare a muchilor maseteri (viteza de ridicare a mandibulei) este invers proporional cu consistena alimentelor. Pe msur ce mandibula se apropie de maxilar n timpul masticaiei, alimentele opun rezisten, iar viteza de ridicare scade naintea stabilirii contactului ocluzal. Fora de presiune vertical ce se exercit n timpul masticaiei este transformat datorit structurii parodontale n for de traciune i transmis asupra esutului osos unde se vor produce fenomene adaptativ-constructive. Muchii mobilizatori ai mandibulei pot dezvolta n timpul masticaiei fore foarte mari (tabelul 3). Ali autori dau valori de aproximativ 10 kg for pe o suprafa de 1 cm2. Presiunile verticale suportate de dini: incisivi: 32 kg canini: 35 kg
premolar II: 44 kg molar I: 45-70 kg molar III: 64 kg. n masticaia obinuit se utilizeaz aproximativ 1/3 din fora maxim, adic fiecare micare masticatorie s-ar nsoi de desfurarea unor fore n jui de 35 kg. Intensitatea forei masticatorii depinde de mai muli factori: de puterea de contracie a muchilor; de sex: la femei, fora dezvoltat la nivelul molarilor este 2/3 din cea a brbailor. de pragul de toleran al parodoniului, respectiv de sensibilitatea parodoniului.
4.3. ROLUL FUNCIONAL AL MASTICAIEI
4.3.1. Masticaia i digestia Concomitent cu frmiarea alimentelor are loc amestecarea lor cu saliv i formarea bolului alimentar, pregtit pentru deglutiie. Insalivarea alimentelor permite nceperea digestiei nc din cavitatea bucal. Micrile masticatorii reprezint i un stimul mecanic ce favorizeaz evacuarea salivei din glandele salivare. Insalivarea alimentelor permite dizolvarea constituenilor alimentari i stimularea receptorilor gustativi cu urmtoarele consecine: - realizarea senzaiei gustative; - aprecierea calitii alimentelor; - stimularea secreiei salivare; - stimularea secreiei gastrice, pancreatice, prin declanarea reflexelor necondiionate. Triturarea corespunztoare a alimentelor asigur o digestie optim. Deglutiia unor alimente insuficient triturate ngreuneaz digestia: apar tulburri gastrice prin efortul digestiv gastric crescut i prin ntrzierea evacurii gastrice. Masticaia rapid are dezavantaje: - nu stimuleaz suficient receptorii bucali; - secreia salivar i gastric sunt insuficiente; - determin un efort digestiv gastric crescut; - induce o digestie i absorbie deficitar; - favorizeaz apariia hipotrofiei la copii. Enzimele digestive acioneaz la suprafaa fragmentelor, de aceea la fragmentele mari, netriturate eficiena de digestie este sczut. Particulele mari necesit un travaliu gastric mai intens i mai ndelungat ceea ce suprasolicit stomacul inducnd afeciuni digestive. 4.3.2. Masticaia i olfacia Zdrobirea corespunztoare a alimentelor permite eliberarea substanelor odorifere i percepia olfactiv. Importana percepiei olfactive n timpul masticaiei: - apreciaz calitatea alimentelor; - declaneaz i susine secreiile digestive: salivare, gastrice, pancreatice prin reflexele condiionate. - asociat cu simul gustativ creeaz o stare psihic de satisfacie sau insatisfacie. 4.3.3. Masticaia i structurile buco-dentare O solicitare normal, n timpul masticaiei a esuturilor sistemului oro-facial are un rol pozitiv n meninerea troficitii sistemului, asigurnd dezvoltarea lui. Masticaia viguroas determin: a - stimularea centrilor osteogenetici de cretere; b - intensificarea trabeculaiei osoase; c - oasele maxilare sunt mai groase, mai rezistente; d membrana (structurile) periodontal se ngroa; e - activarea circulaiei n musculatura masticatorie, n structurile osoase, rezultnd creterea aportului de substane nutritive, hormoni; f- favorizarea autocuririi cavitii bucale;
g - cheratinizarea i multiplicarea stratului bazai al mucoasei gingivale; h stimularea reaciilor de aprare; i - scderea depunerii de tartru; 1 - prevenirea formrii cariilor dentare. Activitatea muscular susinut influeneaz oasele maxilare prin: a - intermediul inseriilor osoase - astfel subiecii care fac micri masticatorii susinute au mandibula bine dezvoltat. La persoanele cu rahitism contraciile musculare puternice deformeaz maxilarul superior, b - aplicarea direct a presiunilor musculare pe suprafeele osoase, c - aplicarea indirect a forelor musculare prin intermediul sistemului dentar. In timpul masticaiei, prin intermediul ligamentelor periodontale, se transform forele de frecare n fore de traciune direct asupra osului alveolar. La nivelul oaselor maxilare se exercit fore de traciune, a cror surs este contracia muchilor mobilizatori ai mandibulei. Forele de masticaie se transmit indirect prin intermediul sistemului dentar. Resorbia proceselor alveolare i mrirea spaiului parodontal la dinii nesolicitai, prin lipsa antagonitilor, reprezint o dovad concludent a lipsei forelor stimulatoare ale masticaiei n structurarea oaselor. Poziia dinilor i restructurarea permanent a pereilor alveolari sunt n funcie de solicitrile funcionale generate de masticaie. n unele anomalii dento-maxilare cu malpoziii dentare, micrile masticatorii se fac defectuos. 4.3.4. Masticaia i sistemul nervos Masticaia are i un rol sedativ, calmant asupra sistemului nervos Contracia prelungit a muchilor masticatori creeaz oboseal i somn la copii La adult, micrile lente continue de masticaie (folosirea gumei de mestecat echilibreaz tonusul muchilor antagoniti i au efect calmant.
4.4. MASTICAIA I ANOMALIILE APARATULUI DENTOMAXILAR
Tulburrile masticatorii sunt factori predispozani pentru producerea anomaliilor dento-maxilare prin: - masticaia insuficient face ca musculatura masticatorie i oasele maxilare s fie mai puin dezvoltate; - orientarea funcional a trabeculelor este mai discret; - parodoniul marginal ofer o implantare dentar mai puin robust; - abraziunea fiziologic lipsete sau este foarte redus; - nu se face mezializarea fiziologic a mandibulei; - secreia salivar mai sczut ntreine o autocurire deficitar, favorizeaz instalarea de procese distructive la nivelul coroanei; - malpoziii, ectopii dentare; - crete frecvena cariilor; - solicitarea insuficient a parodoniului, favorizeaz apariia parodontopatiilor. Anomaliile dento-maxilare instalate micoreaz eficiena masticatorie prin
mai multe mecanisme: - reducerea suprafeei de contact ocluzal; - absena sau dificultatea n efectuarea unor micri masticatorii; - limitarea micrilor masticatorii prin blocaje; - lezarea direct a mucoasei de pe maxilarul antagonist, de ctre dini, determin senzaia dureroas care reduce i mai mult fora de contracie. 4.5. REGLAREA MASTICAIEI Masticaia este un act complex, parial voluntar, parial reflex prin care se asigur coordonarea activitii musculare n vederea prelucrrii mecanice a alimentelor. Masticaia are o complexitate care este unic n tot organismul. Reglarea masticaiei se face prin mecanism nervos, la care particip zone din aria cortical motorie, din trunchiul cerebral, corelate cu informaiile senzitive din zona oro-facial. Masticaia ncepe cu deschiderea voluntar a gurii i coborrea mandibulei induse de ctre impulsurile motorii corticale venite pe ci piramidale i extrapiramidale la nucleii motori ai nervilor cranieni V, VII, XII, din trunchiul cerebral. Actul motor al masticaiei este modulat i de impulsuri cerebeloase. Ablaia cerebelului produce ataxie, modificri motorii ce afecteaz i prehen-siunea hranei. Ridicarea reflex a mandibulei se realizeaz prin reflexul miotatic maseterin dinamic cu urmtoarea succesiune: - deschiderea gurii determin ntinderea muchilor ridictoii ai mandibulei; - se stimuleaz fusurile neuro-musculare din maseter; - fibrele anulo-fusale transmit direct excitaia la nucleul senzorial mezencefalic al trigemenului; - dup o ntrziere de 0,7 ms informaia este transmis monosinaptic la nucleul motor al trigemenului, din punte; - impulsurile motorii iau calea neuronilor motorii alfa spre muchii ridictori ai mandibulei; - contracia m. ridictori determin ridicarea mandibulei, nchiderea gurii, realizarea contactului ocluzal pe alimentele din cavitatea bucal. n continuare se declaneaz reflexul de coborre coordonat a mandibulei, coborre ce permite rearanjarea alimentelor ntre suprafeele ocluzale ale dinilor. Succesiunea fenomenelor de coborre reflex a mandibulei: - alimentele nchise n cavitatea bucal stimuleaz presoreceptorii din: mucoasa gingival, ligamente periodontale, pulp, dentin, mucoas bucal, lingual, palatul dur, etc; - impulsurile generate de stimularea presoreceptorilor sunt transmise nucleului senzorial al n. V din punte, prin n. dentari i linguali; - din nucleul senzorial, principal, al trigemenului impulsurile ajung la nucleul motor al n. V, dup o ntrziere de 1,7 msec. - n nucleul motor se genereaz un potenial de excitaie postsinaptic n motoneuronii muchilor cobortori i un potenial de inhibiie postsinaptic n neuronii motori ai muchilor ridictori;
- se trimit comenzi efectorii pentru muchii cobortori i inhibitorii pentru muchii ridictori; - rspunsul motor este promt, asigur deschiderea gurii, protejeaz arcadele dentare de o suprasolicitare mecanic. Impulsurile proprioceptive de la ATM controleaz activitatea motorie a mandibulei, limiteaz micrile, readapteaz masticaia dup intervenii ortodontice, transmit i stimuli de durere. Reflexul necondiionat masticator are centrul n nucleul masticator din protuberanta (punte). Acest centru primete aferente tactile proprioceptive, gustative pe calea nervilor V, VII, IX. Nucleul masticator pontin este coordonat de centrii corticali n vederea finalizrii masticaiei i a realizrii bolului alimentar. Leziunile ariei motorii corticale suprim reflexul masticator ca act coordonat i face imposibil realizarea bolului alimentar, deoarece persist numai micri stereotipe de ridicare i coborre a mandibulei fr coordonare lingual. orofacial. Sistemul limbic, hipotalamusul, formaia reticulat, cerebelul contribuie la reglarea contraciei musculare n timpul masticaiei. Durata unui ciclu masticator este de aproximativ o secund. n afeciuni dentare, plasarea de proteze n gur, stereotipul masticator se modific contient, elaborndu-se un nou tip de masticaie. Activitatea motorie a centrilor masticatori este influenat i de statusul umoral al organismului: n hipoglicemie se amplific potenialul motor al muchilor ridictori.
FIZIOLOGIA DEGLUTIIEI
Deglutiia reprezint totalitatea fenomenelor care concur la trecerea bolului alimentar sau a salivei din cavitatea bucal n faringe i esofag (fig, 16). Deglutiia este o funcie permanent ce se desfoar att n perioadele de veghe, ct i n cele de somn. Numrul de deglutiii n 24 ore variaz ntre 590 pn la 1600, dup diferii autori.
5.1. TIMPII DEGLUTIIEI
Se descriu trei timpi sau etape ale deglutiiei i anume: timpul bucal, timpul faringian i timpul esofagian. 5.1.1. Timpul I, bucal al deglutiiei Este un timp voluntar, dureaz 1 secund i const din transportarea bolului alimentar din cavitatea bucal n faringe prin micri coordonate ale limbii, palatului moale, mandibulei i complexului hioidian. Succesiunea fenomenelor timpului bucal al deglutiiei: - bolul alimentar este plasat pe faa dorsal a limbii; - muchii intrinseci dau limbii forma de "jgheab" n care se muleaz bolul
alimentar - vrful limbii se sprijin pe bolta palatin i se rsfrnge napoi; - contracia muchilor milohioidieni ridic planeul bucal odat cu limba, care apas cu toat fora pe bolta palatin, mpingnd bolul spre faringe; - prin contracia m. hiogloi i stilogloi se trag napoi prile laterale ale bazei limbii, modelnd-o "n jgheab" prin care bolul alimentar alunec spre faringe, existnd i o und peristaltic a limbii. Limba exercit pe bolta palatin o presiune ntre 41 i 70 g/cm2, presiune dependent de consistena alimentelor. Aceast presiune se exercit mai ales pe prile laterale ale boitei. Presiunea excesiv a limbii n poriunea posterioar a bolii palatine determin vestibulizarea dinilor. Timpul I al deglutiiei se schimb cu vrsta.
5.1.2. Timpul II, faringian al deglutiiei Timpul faringian al deglutiiei este scurt (1 secund), autonom. Contactul alimentelor cu diverse pri ale gurii i faringelui declaneaz o succesiune de reflexe care au drept scop asigurarea condiiilor pentru ca bolul alimentar s coboare n esofag i s nu ptrund n laringe sau n nazo-faringe. Zonele reflexogene cele mai sensibile sunt reprezentate de: - mucoasa pilierilor anterior; - peretele posterior al faringelui;
- stlpii posteriori ai amigdalelor; - amigdale; - baza limbii; - lueta; - palatul moale. Aceast zon se numete zona reflexogen Vasilieff. Progresiunea bolului alimentar n faringe este asigurat de contracia reflex a constrictorului superior al faringelui deasupra bolului, obligndu-1 s treac n esofag. naintea bolului, n faringe i esofag, se creeaz o presiune negativ, de aproximativ 35 cm3 ap. Alte ci posibile de angajare a bolului alimentar n timpul faringian al deglutiiei sunt: - napoierea bolului alimentar n cavitatea bucal este prevenit prin contracia m. palatoglos i palatofaringian, care ngusteaz istmul bucal. Bolul poate fi readus n gur printr-un reflex special de tuse, de curire a faringelui; - ptrunderea alimentelor n cavitatea nazal este prevenit prin nchiderea orificiului nazofaringian realizat de contracia m. peristafilini externi i a muchilor ridictori i tensori ai vlului palatin; - ptrunderea alimentelor n laringe este mpiedicat prin separarea cii digestive de cea respiratorie. Orificiul laringofaringian se nchide prin: ridicarea mandibulei, a hioidu-lui, laringelui i a limbii i prin coborrea epiglotei pe laringo-faringe. La aceste micri complexe particip: - m. ridictori ai mandibulei; - m. suprahioidieni, care sprijinindu-se pe mandibul, ridic hioidul i apoi cartilajul tiroid; - hioidul mobilizat n sus i nainte, trage n aceai direcie baza limbii; - epiglota se deplaseaz n jos, prin micri foarte rapide, nchiznd calea aerian, ca un capac; - ridicarea laringelui (prin contracia m. tirohioidian), face ca el s ajung sub protecia epiglotei i bazei limbii. n cazul n care laringele este fixat printr-un proces patologic, deglutiia este imposibil. La sugar, laringele este ntr-o poziie mai nalt i de aceea este posibil s se efectueze simultan suptul i respiraia, laptele curgnd pe pereii laterali ai faringo-laringclui; - laringele se nchide i prin alturarea corzilor vocale. 5.1.3. Timpul III, esofagian al deglutiiei Timpul esofagian ncepe cu deschiderea orificiului superior al esofagului prin mobilizarea nainte a peretelui anterior i iniierea unei unde peristaltice. n poriunea cervical a esofagului (pe o lungime de 6-7 cm) care are muchi striai, deplasarea bolului se face foarte rapid (o secund). n poriunea toracic a esofagului (alctuit din fibre musculare striate i netede) deplasarea se face mai lent, n 1,5-2 secunde. n poriunea inferioar, esofagul este alctuit numai din musculatur neted, iar deplasarea dureaz 3 secunde. n total timpul esofagian al deglutiiei
dureaz 6-7 secunde. Orificiul cardia se deschide reflex, lsnd bolul alimentar s treac n stomac.
Coordonarea micrilor necesare deglutiiei se realizeaz prin mecanisme nervoase (fig. 17). Declanarea deglutiiei se poate face reflex (cu punct de plecare de la receptorii din regiunea bucal) dar i voluntar (cu participarea scoarei cerebrale). 5.2.1. Calea senzitiv, aferent este asigurat de nervii: trigemen, glosofaringian, vag, care transmit impulsuri din cavitatea bucal i zona reflexogen Vasilieff. 5.2.2. Centrul deglutiiei este localizat n bulb, n substana reticulat, n vecintatea centrilor respiratori, salivatori, cardio-vasculari. n timpul deglutiiei sunt inhibai centrii respiratori i centrul masticator. Deglutiia unui bol alimentar voluminos induce reflexe salivare, lacrimale, vasomotorii. Centrul deglutiiei are legturi cu nucleii: masticatori, al nervilor faciali, hipogloi. 5.2.3. Calea eferent este reprezentat de ramurile motorii ale nervilor: - trigemen, asigur ridicarea mandibulei i nchiderea gurii; - facial, controleaz muchii buzelor i ai mimicii; - hipogloi, asigur micrile coordonate ale muchilor limbii; - glosofaringian, pentru musculatura faringian; - vag, pentru musculatura esofagian. In partea superioar a esofagului, inervaia vagal declaneaz contraciile eristaltice i asigur progresiunea lor n funcie de impulsurile descrcate de centrul bulbar. In esofagul mijlociu i inferior, propagarea undelor este asigurat e inervaia intrinsec a plexului mienteric Auerbach. Progresiunea undelor este controlat de fibrele vagale care stimuleaz peristaltismul, relaxeaz cardia i de fibrele simpatice care inhib peristaltismul i contract sfincterul cardia. Sfincterul cardia are o activitate tonic i fazic sub aciunea vagului, care crete tonusul i deschide fazic sfincterul, n faa fiecrei unde peristaltice. Deglutiia infantil este coordonat de nervii faciali i hipogloi, iar, deglutiia adultului este controlat de trigemen i hipoglos. La edentatul total, pierderea dinilor duce la o revenire ctre deglutiia controlat de facial, n care orbicularul buzelor joac rol primordial, evitndu-se stressul ATM.
5.2. REGLAREA DEGLUTIIEI
Tulburrile deglutiiei: - disfagia, reprezint dificultatea de a nghii. Se ntlnete n: amigdalite, leziuni ale limbii, aparate ortodontice, lezarea centrilor nervoi; - paralizia deglutiiei: n come uremice, hepatice, narcoz profund, intoxicaii cu morfin, barbiturice.
5.3. PARTICULARITI ALE DEGLUTIIEI
5.3.1. Deglutiia la nou nscut se caracterizeaz prin pstrarea maxilarelor deprtate, contraciile buzelor i obrajilor fiind puternice. Deglutiia cu arcadele deprtate se numete deglutiie de tip infantil (ce dureaz pn la 6 luni). Nou nscutul duce limba nainte, prinznd mamelonul ntre ea i creasta alveolar superioar. Partea dorsal a limbii formeaz un jgheab prin care laptele curge spre faringe ajutat de contraciile muchilor linguali. Buzele asigur nchiderea etan a cavitii bucale. n acest timp, maxilarele sunt deprtate avnd limba aezat n partea anterioar, iar n prile laterale se interpune mucoasa jugal i musculatura obrazului. n repaus, crestele maxilare fiind foarte puin dezvoltate permit ptrunderea limbii ntre cele dou maxilare, meninndu-le deprtate. 5.3.2. Etapa tranziional In jurul vrstei de 6 luni odat cu erupia incisivilor se modific treptat comportamentul limbii: limba ia o poziie mai posterioar (napoia incisivilor), se lete, poriunile laterale ptrund ntre crestele celor dou maxilare, nc edentate. In timpul deglutiiei, limba rmne napoia incisivilor, buzele se nchid, au loc contracii ale muchilor mimicii. In timpul erupiei caninilor i molarilor temporari, limba n repaus, ptrunde ntre arcadele dentare n regiunile laterale, contribuind la fixarea unei poziii mandibulare, deci se schieaz, deja, "inocluzia fiziologic". Acest stadiu tranziional, dureaz de la 6 luni pn la, aproximativ, 2 ani. 5.3.3. Deglutiia adultului Deglutiia de tip adult este o deglutiie cu arcadele n contact i se ealizeaz n urmtoarele condiii: - ingerarea de alimente solide sau pstoase; - ingerarea de lichide n cantiti mici; - deglutiia salivei. Ingerarea de alimente zemoase, de cantiti mari de lichide se realizeaz, i la adult, n condiiile pstrrii arcadelor dentare deprtate. 5.3.4. Deglutiia n prezena aparatelor ortodontice Aparatele ortodontice acioneaz mpiedicnd interpunerea limbii ntre arcade ceea ce determin: - nlturarea forelor anormale pe care limba le exercit la nivelul arcadelor dentare; - normalizarea morfologic; - reeducarea deglutiiei; - mecanismele neuro-musculare orale se adapteaz noii situaii.
5.4. DEGLUTIIA I ANOMALIILE APARATULUI DENTOMAXILAR
Prelungirea i permanentizarea stadiului de deglutiie infantil reprezint o incapacitate a copilului de a-i adapta micrile la noile condiii, create de
erupia dentar, incapacitate care este consecina unei ntrzieri n maturarea nervoas i muscular. Succesiunea fenomenelor n imaturitatea neuro-muscular: - permanentizarea stadiului de deglutiie infantil sau stadiului tranziional datorit imaturitii neuro-musculare; - se genereaz fore cu influen negativ asupra dezvoltrii aparatului dento-maxilar; - apar anomalii dento-maxilare. Deglutiia infantil se poate instala ca un fenomen de adaptare la modificrile produse de anomalii dento-maxilare. Revenirea la deglutiia infantil se poate produce i: - datorit obiceiului vicios de sugere a degetului; - n perioada nlocuirii dinilor temporari cu cei permaneni; - n caz de macroglosie. Persistena unei deglutiii de tip infantil se poate datora unei "imaturitai" neuro-musculare datorit : - unor cauze locale: sugerea degetului, afeciuni rino-faringiene, fenomene locale legate de nlocuirea dinilor; - unor cauze generale: acromegalia asociat cu macroglosia. De asemenea, deglutiia de tip infantil poate fi rezultatul unor anomalii ale aparatului dento-maxilar.
ALIMENTAREA NATURAL I ARTIFICIAL A SUGARULUI
Imediat dup natere, copilul poate s sug datorit unui reflex nnscut, automat, care exist la toate mamiferele.
Alimentaia natural la sn sau suptul la sn se caracterizeaz prin micri ritmice: - de nfundare i umflare a obrazului; - de ridicare i coborre a hioidului; - de propulsie i retropulsie a mandibulei; - ale limbii, obrajilor; - de nclinare sau extensie a capului; - de strngere i relaxare a sfincterului labial. Acestea se asociaz cu micri ale minilor de prindere a mamelei i contracia celulelor mioepiteliale indus de oxitocin. Mecanismul suptului cuprinde dou micri distincte: - una de succiune, care necesit realizarea unui vid n cavitatea bucal, iar - alta de presiune care const din aciunea de exprimare a laptelui din mamelon. In timpul suptului mamelonul i o parte a areolei sunt atrase n gura
6.1. SUPTUL LA SN
copilului, mamelonul fiind mult alungit. Baza mamelonului este prins ntre gingia superioar i vrful limbii. Limba formeaz un adevrat jgheab i realizeaz o micare ondulatorie (ca o und peristaltic), de la vrf spre baza limbii, ce preseaz asupra mamelonului. Micrile mandibulei asigur modificrile de presiune (10-15 cm H20 sub presiune atmosferic), n interiorul cavitii bucale. Buzele asigur att nchiderea ermetic a cavitii bucale ct i compresiuni ritmice la nivelul areolei.
- asigur creterea aparatului dento-maxilar; - realizeaz funcionarea armonioas a aparatului dento-maxilar; - solicit muchii masticatori; - sunt solicitate toate grupele musculare ale aparatului dento-maxilar (ADM), avnd loc ntr-o succesiune ritmic, contracia unui grup de muchi urmat de contracia antagonitilor; - activitatea muscular determin dezvoltarea echilibrat i proporional a elementelor osoase pe care se insera; - vrful limbii exercit pe bolt o presiune de 50-60 g/cm2; - micrile ritmice ale limbii fac ca ea s aib rolul unei pompe aspiratoarerespingtoare pentru jetul de lapte din mamelon; - micrile ritmice ale limbii au repercursiuni directe i asupra debitului sanguin din mucoasa boitei palatine, ceea ce determin o dezvoltare armonioas a boitei palatine i a foselor nazale; - limba, stimuleaz funcia osteogenetic de cretere a suturii incisivocanine i intermaxilare, bolta dezvoltndu-se normal n lime i lungime; - are loc insalivarea laptelui i pregtirea digestiei optime; - sunt antrenai i se echilibreaz aciunea grupelor musculare antagoniste, care vor fi ulterior solicitate n cazul alimentaiei mixte; - aciunea trofic i stimulatoare a muchilor pentru celulele osteogenetice de cretere contribuie la dezvoltarea echilibrat a elementelor ADM ca i a elementelor sistemului de susinere a capului; - prin efortul muscular realizat n timpul suptului viguros se induce un somn reconfortant i linititor.
6.2. AVANTAJELE SUPTULUI LA SN
6.3. ALIMENTAREA ARTIFICIAL A SUGARULUI
Alimentaia prin biberon a sugarului determin adaptarea contraciilor musculare corespunztor noilor condiii. Buzele nu mai nchid ermetic fanta labial, pentru c nu este necesar, deoarece laptele curge singur prin greutatea lui. Orbicularii buzelor nu sunt solicitai, nu se mai "antreneaz" iar dezvoltarea lor deficitar se va manifesta mai trziu prin hipotonia buzelor. De asemenea, nu au loc nfundarea i umflarea obrajilor, nefiind nevoie de o presiune oral, care s aspire laptele i n consecin buccinatorul se va dezvolta insuficient, va fi hipoton i cu for de contracie redus.
Micrile de ridicare i coborre ale hioidului efectuate de m. suprahioi-dieni i ridictorii mandibulei nu mai sunt necesare, laptele din biberon curgnd singur, fr s fie obligatorie o presiune negativ aspiratoare n cavitatea bucal. Lipsa de solicitare a grupelor musculare antagoniste, ntr-o aciune echilibrat, se repercuteaz asupra dezvoltrii insuficiente a mandibulei n raport cu maxilarul i arcadele alveolare superioare. Lipsa de solicitare a grupelor musculare antagoniste, ntr-o aciune echilibrat, se repercuteaz asupra dezvoltrii insuficiente a mandibulei n raport cu maxilarul i arcadele alveolare superioare. Nu se mai efectueaz propulsia i retropulsia mandibulei pentru c nu mai este necesar presarea mamelei prin mentonul sugarului. Va rezulta o dezvoltare deficitar a mandibulei n aceast perioad, cnd are loc o cretere intens. Cnd sugarul este hrnit cu biberonul, vrful limbii se modeleaz "n jgheab" pentru a asigura trecerea uoar a laptelui spre faringe. n aceast situaie, vascularizaia boitei palatine i centrele ei osteogenetice nu mai sunt stimulate i dezvoltarea nu se mai realizeaz armonios. Se ajunge la formarea unei boite adnci, strmtat (ogival) iar planeul foselor nazale este ngustat, perturbnd respiraia nazal.
6.4. DEZAVANTAJELE ALIMENTRII ARTIFICIALE A SUGARULUI
- micrile de propulsie sunt reduse pentru c laptele curge uor i n cantitate mai mare; - aerofagia, adic ingestia mpreun cu laptele a unei cantiti de aer; - reducerea secreiei salivare; - insalivarea insuficient a laptelui i tulburri de digestie; - prin absena stimulilor funcionali de propulsie a mandibulei se perturb dezvoltarea mandibulei i se pericliteaz aezarea corect a primelor elemente dentare; - se pot constitui disfuncii n dezvoltarea ADM cu repercursiuni asupra dezvoltrii generale a copilului.
6.5. REGLAREA NERVOAS A SUPTULUI
Suptul este un act reflex nnscut, cu origine bulbo-protuberanial. Cile centripete, aferente ale acestui reflex sunt incluse n ramurile senzitive ale trigemenului. Cile centrifuge, eferente sunt reprezentate de nervii trigemeni, faciali, hipogloi, prin intermediul crora este asigurat contracia muchilor corespunztori i ai limbii. Micrile mandibulei necesare suptului prezint la nceput un control tactilokinestezic, iar ulterior, odat cu dezvoltarea componentelor sistemului orofacial i a experienei alimentare se instaleaz un control reflex tactilogustativo-kinestezic.
Trecerea de la alimentaia la sn la alimentaia diversificat impune adaptarea ADM la noile situaii: - micarea obrajilor, limbii, buzelor, mandibulei se complic; - micrile dirijate de sistemul nervos se adapteaz la consistena alimentelor; - formarea bolului alimentar, insalivarea lui i mpingerea spre faringe se nva n fraged copilrie; - acest lan de reflexe condiionate, nvate vor deveni automate; - odat cu apariia dinilor, copilul ncepe s nvee s mestece; - dup apariia primilor molari se constat prezena aproape a tuturor micrile mandibulei ca i la adult.
FONAIA
Laringele face parte din structura cilor aerifere permind trecerea bidirecional a aerului, n procesul ventilaiei pulmonare. De asemenea, este elementul de baz n producerea sunetelor.
7.1. STRUCTURA FUNCIONAL A LARINGELUI
7.1.1. Muchii laringelui Muchii laringelui pot fi grupai n dou grupe musculare antagoniste: - m. constrictori ai glotei care au funcie de adductori ai corzilor vocale sunt: m. cricoaritenoizi anteriori, laterali, m. tiroaritenoizi, m. interaritenoizi, m. anteroepiglotic. Contracia acestor muchi determin apropierea cartilajelor aritenoide ntre ele i ngustarea orificiului glotic. Muchii sunt inervai de nervii recureni. Paralizia unilateral a nervului recurent determin vocea bitonal. - m. dilatatori ai glotei determin deprtarea corzilor vocale, sunt m. cricoaritenoizi posteriori. 7.1.2. Corzile vocale 7.1.2.1. Structura corzilor vocale Corzile vocale sunt dou formaiuni musculo-tendinoase aezate orizontal. Lungimea medie a corzilor vocale variaz cu: - sexul: - 15 mm la sexul masculin; - 11 mm la sexul feminin; - vrsta: - la copil = 7 mm. La pubertate, corzile vocale se alungesc la fete cu 1/3 (10-11 mm) iar la biei se dubleaz (14-15 mm). Fiecare coard vocal este alctuit n principal din 2 muchi, ce au un metabolism de tip anaerob, ceea ce determin o semifatigabilitate a muchilor vocali. Marginile libere ale corzilor vocale sunt alctuite din esut conjunctiv elastic acoperite de un epiteliu stratificat. Deasupra corzilor vocale se gsesc "benzile ventriculare" sau falsele corzi vocale ce conin numeroase glande mucoase. Mucusul acestor glande, lubrefiaza corzile vocale adevrate. In timpul respiraiei obinuite, corzile vocale sunt deprtate, lsnd ntre ele
un spaiu, numit orificiu glotic. Corzile vocale sunt ntinse de muchii cricotiroidieni (m. intrinseci) inervai de nervul laringeu superior. n paralizia nervului laringeu superior corzile vocale sunt flasce, iar vocea este rguit. Corzile vocale sunt relaxate de ctre m. tiroaritenoizi interni. 7.1.2.2. Rolul corzilor vocale n fonaie Corzile vocale au rol activ n fonaie. Ele se apropie reciproc i se deprteaz ca urmare a contraciei muchilor proprii i muchilor limitrofi, contracie dependent de impulsurile nervoase venite pe calea nervilor recureni, n salve. Moulanguet n 1953 a pus n eviden homoritmia dintre activitatea nervilor recureni i deschiderile glotice. n fonaie, contracia activa a corzilor vocale determin deschideri i nchideri ritmate ale glotei, secionarea coloanei de aer i producerea sunetelor, dac presiunea subglotic este suficient. n cursul fiecrei deschideri, trece o anumit cantitate de aer. nchiderile ritmice realizate de corzile vocale secioneaz coloana de aer, imprimndu-i un caracter de intermiten. Frecvena sunetului este dependent de deschiderile glotice care se coreleaz cu frecvena salvelor din nervii recureni. Deplasarea corzilor vocale se face n sens orizontal cu 4 mm maximum iar n sens vertical cu 0,2-0,5 mm, deci, corzile vocale descriu o micare elipsoidal, cu axul mare orizontal. Micarea complet a corzilor vocale, sau ciclul corzilor vocale se realizeaz n patru faze: - faza de deprtare a corzilor vocale, deschiderea glotic sau abducia realizat de muchii proprii mpreun cu m. cricoaritenoid posteriori; - faza de elongaie maxim, n care corzile vocale rmn deprtate (un timp variabil), sub aciunea m. cricotiroidieni; - faza de apropiere, adducie sau de nchidere a glotei, n poziia poste-rioar (prin contracia numai a m. aritenoidieni), anterioar (prin contracia numai a m. cricoaritenoidieni laterali), pe toat lungimea corzii (prin contracia ambilor muchi). Astfel, corzile vocale vin n contact prin marginile lor libere (ce conin esut conjunctiv elastic). n micarea de adducie, corzile vocale pot prezenta modificri de form (ngrori pariale), n special n vocea cntat; - faza de acolare, n care corzile vocale rmn alipite pn la declanarea salvei recureniale urmtoare. Ciclii de activitate ai corzilor vocale n fonaie se realizeaz cu o mare frecven, astfel: - n vorbire: - 90 cicli/secund, la un brbat cu voce profund; - 300 cicli/secund, la o femeie cu voce nalt; - n cntatul de oper: 65-1300 cicli/secund. 7.1.3. Activitatea muchilor laringelui (mobilitatea laringelui) n timpul vorbirii sau cntatului muchii laringelui se contract sau se relaxeaz coordonat. Muchii inspiratori i cei expiratori coordoneaz introducerea aerului n plmni, respectiv expirarea lui sub presiune. Presiunea
aerului expirat d intensitatea vocii. In timpul inspirului se relaxeaz muschii constrictori i se contract muschii dilatatori ai glotei. nlimea sunetelor este dependent de ntinderea i vibrarea reglat a corzilor vocale, sub influena muchilor tensori sau muchilor relaxani ai corzilor. n timpul expiraiei i al emiterii sunetelor nalte (acute), laringele este ridicat n sus i nainte. n timpul inspiraiei sau n emiterea sunetelor joase laringele coboar. Extensia capului se asociaz cu ridicarea laringelui, iar flexia capului se nsoete de coborrea laringelui. n timpul somnului, este absolut necesar echilibrul dintre m. constrictori i dilatatori ai glotei, pentru a se permite o respiraie linitit. Poziia laringelui este meninut prin: - continuitatea cu faringele (care este fixat prin m. ridictori i constrictori ai faringelui); - muchii i ligamentele care l fixeaz de hioid. Cnd hioidul este fixat, laringele este ridicat de m. tirohioidieni i de ridictorii faringelui iar cnd mandibula este fixat, laringele este ridicat de m. suprahioidieni. Laringele este cobort de m. sternotiroidieni.
7.2. PAVILIONUL FARINGO-BUCAL
Aerul expirat ntmpin din partea pereilor o anumita rezisten numit impedan de scurgere. Coloana vibratorie de aer traverseaz partea superioar a tubului fonator adic pavilionul faringo-bucal, n pavilionul faringobucal are loc o reducere a intensitii sunetului. Funciile pavilionului faringo-bucal: - exercit asupra "valurilor" acustice absorbii selective dnd vocii timbru caracteristic; - pereii elastici rein o parte din vibraii; - aerul capt o micare de turbion, energia cinetic necesar este sustras din energia acustic; - o parte din energia vibratorie este transmis la masele aeriene semi-nchise: sinusurile faciale, cavitile aeriene subglotice, esofag; - are o influen retroactiv asupra efectorului laringian (feed-back-ul mtoreglrii n fonaie): comanda din cortexul motor transmis muchilor fonatori nduce realizarea unui sunet care prin conducere aerian sau conducere osoas ajunge ia receptorul auditiv. Pe cile specifice auditive informaia ajunge ncortexul senzorial realiznd senzaia auditiv. De aici, prin conexiunii* cortexul motor se moduleaz corespunztor activitatea muchilor fonatori. - particip la modularea vocalelor; - particip la formarea consoanelor.
7.3. MECANISMUL FONAIEI I AL ARTICULRII FONOMENELOR
Fonaia este ansamblul de fenomene fiziologice care concur la producerea vocii sub influena stimulul nervos (Clement Munier). Se consider c fonaia are dou faze: - faza subglotic, n cadrul creia aerul comprimat de plmn este expulzat prin laringe fcnd s vibreze corzile vocale; - faza supraglotic cnd sunetul ptrunde n pavilionul faringo-bucal n care vlul palatin, limba, buzele i arcadele alveo Io -dentare i modific caracteristicile, transformnd sunetul laringian n foneme sau vorbire articulat. Fonaia este precedat de inspirarea aerului n plmni. Aerul expirat cu presiune trece prin strmtoarea laringian inducnd vibrarea corzilor vocale. Forma i grosimea corzilor vocale sunt asigurate de contracia difereniat a m. tiroaritenoizi. Modificarea lungimii corzilor vocale se face prin contracia sau relaxarea muchilor cricoaritenoizi posteriori. Aceste micri (contracii musculare) sunt reglate de n. recurent. Contraciile musculare asigur gradul de ntindere a corzilor vocale determinnd emiterea unor sunete mai nalte sau mai joase (grave). Sunetul fundamental nedifereniat produs de vibrarea corzilor vocale are dou lungimi de und: - ntre: 375 la 825 unde/sec. - ntre: 800 la 2.400 unde/sec. Sunetul emis printr-un numr de vibraii pe minut este modelat n cutia de rezonan a cavitii bucale, unde se realizeaz, mai nti "compresiunea aerului" prin nchiderea orificiilor bucale i contracia buccinatorilor. Urmeaz apoi "explozia", cnd coloana de aer vibratoare i comprimat "scap'-' prin alte orificii create de: - cele dou buze; sau - de buza superioar cu dinii frontali; sau - de limb cu dinii frontali; sau - de limb cu bolta palatin; sau - prin modificri ale lumenului nazofaringian sau laringofarmgian. FONEMA reprezint cea mai mic unitate sonor care are funcia de a diferenia cuvintele ntre ele precum i formele gramaticale ale aceluiai cuvnt..
7.3.1 Vocalele Vocalele sunt produse direct la nivelul laringelui prin modificri variabile ale poziiei corzilor vocale ( deprtare, acolare succesiv ) care ntrerup
coloana de aer,imprimndu-i o micare vibratorie specific pentru fiecare vocal. Dar i pavilionul faringo-bucal prezint modificri n timpul emisiei vocalelor. Astfel au loc : deplasri ale limbii, mandibulei, buzelor n cursul emisiei vocalelor: pentru pronunia vocalei a se ngusteaz spaiul dintre epiglot i peretele posterior al faringelui; pentru pronunia vocalei I se reduce spaiul de trecere dintre faringe i baza limbii; vocala u se pronun prin mpingerea napoi a bazei limbii; vocala o se formeaz prin ngustarea n ansamblu a faringelui; vocala e se pronun prin ngustarea moderat a spaiului dintre palat i faa dorsal a limbii. 7.3.2Producerea consoanelor Pentru producerea consoanelor coloana de aer preparat fonetic de cicluri activitilor corzilor vocale sufer secionri de form i ntindere variabile la nivelul pavilionului faringobucal, limba, buzele, lueta efectund diverse micri de inflexiune (fig. 18) Dup nivelul la care se face ntreruperea coloanei de aer consoanele se pot mpri n: consoane labiale (b, m, p) ce sunt produse prin nchiderea i redeschiderea brusc a buzelor; consoane labio-dentale (f, v): buza inferioar vine n contact cu incisivii superiori i se rentoarce repede n poziia iniial; consoane lingvo-dentale (d, t, n) sunt produse prin retragerea vrfului limbii de pe incisivi pe palat, imediat napoia acestora; consoane lingvo-palatale (g, c): pentru pronunia crora limba stabilete contacte cu regiunea mijlocie sau chiar posterioar a palatului dur. De asemenea, consoanele se pot clasifica dup gradul de ntrerupere i forma devierii coloanei de aer n: consoane explozive: p, b, t, d, g, c; consoane fricative: f, v;
consoane consoane consoane consoane
africative: s, j; nazale: m, n; laterale: l; rulate sau vibrate: r.
7.3.3 Tulburrile de fonaie Pronunia fenomenelor este condiionat de integritatea buzelor, limbii, luetei, a sistemelor neuro-musculare care le mobilizeaz i de o dezvoltare psihic corespunztoare. Tulburri de fonaie pot fi: dislalia: imposibilitatea de a pronuna unele fenomene; paradislalia: pronunarea unor fenomene n locul altora; Corectarea tulburrilor fonetice se face prin: plastia defectelor de buz, palat; tratamentul anomaliilor dento-maxilare; reeducare fonetic. Inocluzia verticl se asociaz cu pronunia defectoas a consoanelor: f, v, b, p, m, n, deoarece buzele nu se pot apropia ntre ele sau cu dinii superiori. Retrognaia mandibular se asociaz cu pronunia defectoas a consoanelor: b, p, m, n, s, z pentru c bolnavii nu pot aduce n poziia corect buzele i limba napoia dinilor frontali superiori. Lipsa dinilor frontali, n perioada de schimbare a dinilor sau dup traumatisme, creeaz dificulti n pronunia: f, v. Tulburrile de vorbire din anomaliile dento-maxilare pot avea consecine nefavorabile din punct de vedere al sntii psihice i al acceptabilitii sociale. De aceea, se impune imperios cunoaterea i tratamentul anomaniilor dento-maxilare ca i corectarea prin reeducare fonetic a tulburrilor de vorbire. Terapia ortodontic poate influena fonaia: aparatele ortodontice creind condiii noi pentru activitatea articulatorie i declannd mecanismele de adaptare funcional. 7.3.4 Caracteristicile vocii umane INTENSITATEA VOCII este proporional cu amplitudinea vibraiilor corzilor vocale, n medie 40-50 decibeli. Depinde de: dezvoltarea toracelui i fora muchilor respiratori. TIMBRUL VOCII: este condiionat de numrul i calitatea sunetelor armonice supraadugate tonului laringian fundamental ca i de forma cavitilor de rezonan. TONUL depinde de frecvena vibraiilor corzilor vocale, de lungimea, grosimea i tensiune coardelor. n vorbirea obinuit frecvena vibraiilor este
de 100-200 cicli/sec, la brbat i 250-300 cicli/sec, la femeie. Fonaia este influenat de factori endocrini i umorali. Astfel STH i hormonii sexuali influeneaz direct caracteristicile vocii. Mutaia fiziologic a vocii n perioada pubertii se datorete n special hormonilor sexuali care induc dezvoltarea organului fonator cu modificarea calitativ a vocii.
7.4. VORBIREA
Vorbirea constituie principala modalitate de exprimare i comunicare a diferitelor forme de activitate neuropsihic uman. mpreun cu exprimarea grafic i gestic vorbirea asigura limbajul oral, scris sau gestual, ca manifestri specifice de comunicare interuman. Vorbirea evolueaz de la simpla emitere de sunete cu modulaii reduse i uniforme, ca rezultat al existenei unui reflex motor nnscut, pn la stadiul de vorbire articulat, coerent, cu particulariti individuale i caracteristici locale, perfectibile n tot cursul vieii. Funciile limbajului, n general i vorbirii, n particular, sunt: comunicativ, cognitiv, emoional-expresiv. Vorbirea ncepe spre sfritul primului an de via prin imitarea cuvintelor auzite, ceea ce presupune integritatea sistemului auditiv. n acest sens pledeaz faptul c surdul din natere rmne mut i c surditatea survenit sub 5 ani determin pierderea limbajului articulat. Dac pronunia nu este fidel, prin compararea auditiv i corectri repetate de articulare se ajunge la pronunarea corect a cuvintelor. nsuirea vocabularului se realizeaz prin condiionarea reflex. Limbajul vorbit se mbogete progresiv prin nvare. Substituindu-se primului sistem de semnalizare cuvintele devin dup expresia lui Pavlov "semnale ale semnalelor", iar vorbirea reprezint cel de al doilea sistem de semnalizare. Limbajul scris ncepe s se dezvolte la vrsta de 5-6 ani, elaborarea sa realizndu-se prin mecanisme reflex condiionate analoge celor ale limbajului oral. Bazele neuro-fiziologice ale vorbirii sunt incomplet elucidate. Aferentele senzitivo-senzoriale auditive, vizuale, tactile, kinestezice etc, ajunse sub forma mpulsuui nervos la nivelul zonelor corticale de proiecie specific sunt integrate, analizate i decodate dnd natere percepiilor primare respective. De la nivelul zonelor corticale specifice, informaiile decodate sunt dirijate prin intermediul ariilor interpretative din jur la zonele de alegere, formare i exprimare a cuvintelor din emisferul dominant. Zonele corticale ale vorbirii sunt reprezentate de aria vorbirii Broca situat la baza circumvoluiei frontale ascendente (aria 44-Brodman) i de aria interpretativ general sau gnozic descris de Wernicke la nivelul zonei postero-superioare a lobului temporal. De fapt ntreaga suprafa cortical fronto-temporo-parieto-occipital dintre centrul vorbirii (Broca) i aria interpretativ Wernicke constituie dispozitivul nervos al limbajului vorbit. Componenta cortical a limbajului vorbit acioneaz prin intermediul cilor nervoase motorii asupra musculaturii organelor fonatorii,
de producere i articulare a sunetelor. Numeroase observaii clinice confirmate i de cercetri clinico-experimentale au demonstrat lateralizarea vorbirii: emisferul stng este dominant la persoanele dreptace i invers, la cele stngace. ntre limbaj i vorbire exist o veritabil unitate dialectic. Limbajul ca expresie material a ideilor, este elementul formator al gndirii umane. Exist o strns unitate ntre limbajul gndit (interior) i cel exprimat verbal (exterior). Substratul neuro-fiziologic al relaiilor existente ntre gndire i limbaj este numai parial comun, ntruct gndirea are sediul predominant n lobul frontal, n timp ce componenta central a limbajului se ntinde de la baza circumvoluiei frontale ascendente (aria Broca) pn n partea postero-superioar a lobului temporal din emisferul dominant.
7.4.1. Afaziile Afaziile sunt definite ca tulburri de articulare, de scriere i de nelegere a cuvintelor auzite i scrise, n condiiile integritii funcionale motorii, auditive, vizuale. Afazia motorie sau de exprimare se manifest prin imposibilitatea subiectului de a articula cuvintele (anartrie) i de a putea exprima ceea ce gndete (afazie de exprimare). Bolnavul nelege cuvintele auzite sau scrise dar este incapabil s pronune cuvintele de rspuns, dei nu prezint nici o paralizie. Aceast afazie se produce prin leziuni ale piciorului celei de a treia circumvoluii frontale stngi i ariei 44 Brodmann. De obicei se nsoete de hemiplegie dreapt i de alte tulburri de vorbire. Afaziile senzoriale se manifest prin lipsa de nelegere a cuvintelor. Bolnavul aude ce i se spune dar nu nelege (surditate verbal) i citete cuvintele scrise dar nu le nelege semnificaia (cecitatea verbal sau alexie). Aceste dou forme de afazie senzorial se nsoesc i de scderea capacitii intelectuale. Surditatea verbal se produce prin leziuni ale ariei 22 Brodmann iar cecitatea verbal prin leziuni ale cortexului parastriat (ariile 19 i 39 Brodmann). In afazia senzorial tip Wernicke, produs prin leziuni ale zonei temporo-parietale, bolnavul este incapabil de a nelege un text scris sau vorbit, dar poate nc pronuna silabe i cuvinte, poate scrie unele caractere. Bolnavul nu mai prezint ns raporturile normale dintre idei i cuvintele exprimate (afazie prin deficit intelectual), vorbirea este incoerent.
CONTROLUL NERVOS AL ACTIVITILOR MOTORII
Activitatea motorie a organismului, n ntreaga sa complexitate, se realizeaz pe baza unor circuite neuronale interreactante, ce asigur realizarea unor deprinderi motrice de importan funcional major. Desfurarea
diferitelor activitii motorii, adaptate circumstanelor de viaa ale organismului necesit un control riguros realizat de numeroase structuri din sistemul nervos.
8.1. CENTRII MOTORI
8.1.1. Cortexul motor Cortexul motor este implicat n elaborarea, controlul micrilor voluntare precise, fine i al micrilor automate reflexe posturale. La nivelul sau se descriu patru tipuri de arii: a) ARIA MOTRICITATII VOLUNTARE este situat la nivelul frontalei ascendente (aria 4 Brodman, cortex motor primar), de unde pleac cile piramidale cortico-nucleare i cortico-medulare. Reprezentarea motorie a diferitelor segmente ale corpului este inegal realiznd "homunculus motor". Cea mai mare reprezentare o are mna i capul, segmente ce realizeaz un numr mare de micri, de mare finee i complexitate. Humunculus motor are o form asimetric cu mna i capul de proporii exagerate, aezate n jos, la nivelul ariei 4 (fig. 19). Teritoriul inferior, corespunztor muchilor laringelui, limbii, gtului, feei se proiecteaz n fasciculul cortico-nuclear, iar restul n fasciculul corticomedular al sistemului piramidal. b) ARIILE MOTRICITATII SEMI VOLUNTARE AUA UTOMATE particip la realizarea activitilor motorii automate i posturale. Aceste arii sunt: - ARIA PARA PIRAMIDA LA (aria motorie secundar) este situat anterior de aria 4, iar prin legturile cu nucleii bazali n formaiunea reticulat mezencefalic asigur coordonarea micrilor de nsoire ale gestului voluntar. - ARIA MOTORIE SUPLIMENTAR este localizat n zona anterioar a lobului paracentral al frontalei ascendente, particip la controlul micrilor automate ale trunchiului, centurii, rdcinii membrelor.
- ARIA MOTORIE INHIBITOARE (aria supresoare) - aria 4S este n conexiune cu nucleii bazali, cu formaiunea reticulat talamic, avnd rol de inhibiie motorie, de modelare a micrilor. - ARIILE OCULOMOTORII sunt implicate n micrile conjugate ale capului i ochilor. c) ARII DE COORDONARE CORTICO-CEREBELOASE implicate n coordonarea dinamic a activitii motorii, locul de origine al fasciculelor frontoparieto-occipito-temporo-ponto-cerebeloase. d) ARIILE PSIHO-MOTORII sunt localizate n regiunea prefrontal (aria 6, aria 8), cuprind principalii centrii ai elaborrii unei micri, sunt sediul fenomenelor de "praxie" cum ar fi: vorbirea, scrierea, etc. Aceste arii prezint conexiuni cu aria 4 somato-motorie. Funciile ariilor motorii corticale sunt: declaneaz diversele activiti notorii, asigur elaborarea, memorizarea i controlul succesiunii actelor motorii. 8.1.2. Formaiuni motorii subcorticale Structurile nervoase subcorticale particip la meninerea i asigurarea
reciziei, duratei, amplitudinii activitilor motorii. Aceste structuri sub-orticale sunt: a) NUCLEU BAZALI controleaz micrile de ansamblu ale corpului care acompaniaz micrile voluntare (nucleul caudat, putamenul) sau micrile diferitelor segmente ale corpului cnd se realizeaz o micare complex (globus pallidus). b) TRUNCHIUL CEREBRAL intervine n activitile motorii: prin prezena nucleilor nervilor cranieni, a unor formaiuni extrapiramidale, a unor centrii refleci, prin trecerea unor importante ci motorii, etc. Astfel trunchiul cerebral particip la: - activitatea motorie reflex: deglutiie, reflexul corneean i auditiv de clipire, auditivo-oculogir. masticator, reflexe pupilare, posturale de redresare, statokinetice; - integrarea neuro-motorie prin centrii tectali, tuberculuii cvadrigemeni; - reglarea reflex postural i la facilitarea reflexelor medulare miotatice prin cile descendente vestibulare; - reglarea reactivitii tonice posturale prin formaiunea reticulat ce are att componente activatoare ct i inhibitoare. Formaiunea reticulat i exercit efectele prin intermediul motoneuronilor alfa i gama, integrai funcional ntr-un sistem de feed-back. Mduva spinrii Mduva spinrii particip la activitile motorii voluntare i reflexe deoarece este punctul final al cilor descendente piramidale i extrapiramidale, unde aceste ci fac sinaps cu neuronii efectori alfa i gama. Funciile cuplului de neuroni alfa-gama: - adapteaz continuu tonusul bazai i postural; - declaneaz, menine i finalizeaz actele motorii reflexe, cu punct de plecare medular sau supramedular; - declaneaz i finalizeaz micrile voluntare comandate cortical; - particip la realizarea stereotipurilor motorii, automate, etc.
8.1.4. Cerebelul Cerebelul din punct de vedere filogenetic i anatomofiziologic se mparte n arhicerebel, paleocerebel i neocerebel. ARHICEREBELUL este un centru al echilibrului gravitaional, deoarece particip la coordonarea reflexelor de redresare labirintice i statokinetice. PALEOCEREBELUL - particip la reglarea tonusului muscular, a contraciei musculare ce asigur echilibrul corpului sau a unor segmente ale corpului (n anumite atitudini posturale). Realizarea acestor funcii este posibil pentru c, la acest nivel, converg cile sensibilitii proprioceptive incontiente, contiente i informaiile de la receptori superficiali stimulai de anumite "posturi" sau "poziii".
NEOCEREBELUL regleaz i coordoneaz micrile generale ale corpului i extremitilor; armonizeaz micrile; asigur, indirect, motricitatea voluntar. Ataxia reprezint lipsa de coordonare a contraciei diferiilor muchi. Se descrie "ataxia cerebeloas" n leziuni ale cerebelului, manifestat prin: tulburri de tonus muscular, dispariia reflexelor de susinere i redresare, disarmonia micrilor, etc. CILE MOTORII DE CONDUCERE Schema clasic a cilor motorii de conducere cuprinde calea piramidal, cu doi neuroni pentru motricitatea voluntar i calea extrapiramidal, multineuronal pentru motricitatea semivoluntar, automat. 8.2.1. Cale piramidal Calea piramidal i are originea principal n cortexul motor, aria 4, dar i n aria premotorie i n aria motorie post central (fig. 20).
FIG.21. CAI DESCENDENTE MOTORII (dupa Guyton, 1996, modificat) n structura cii piramidale se difereniaz fasciculele: - cortico-nucleare pentru nucleii motori din trunchiul cerebral; - cortico-medulare (spinale) pentru motoneuronii din coarnele anterioare ale mduvei spinrii (fig. 21). Conectarea cii piramidale la motoneuronii (alfa i gama) se realizeaz contralateral fie indirect, prin neuronii intercalri (pentru musculatura flexoare proximal a membrelor) fie direct (pentru muchii degetelor sau ai sistemului oro-facial). Calea piramidal format din 2 neuroni (corticali i medulari) conduce rapid comanda pentru motricitatea voluntar dar i pentru micrile automate ce nsoesc un act motor voluntar. Calea extrapirarnidai nervos: Calea extrapirarnidai i are originea n diverse zone ale sistemului
- centrii corticali sunt: cortexul prefrontal (ariile 6 i 8), aria supresoare 4S, cortexul parietal (ariile 1,2,3,5), cortexul temporal (aria 21), cortexul occipital (aria 19); - centrii tectali sunt localizai la nivelul tuberculilor cvadrigemeni ce sunt n conexiune cu corpii geniculai; - centrii optostriai situai la nivelul corpilor striai; -nucleii trunchiului cerebral: nucleul rou, oliva, complexul vestibular i substana reticulat ce intervin major n reglarea tonusului muscular; - cerebelul - prin care se asigur coordonarea static i dinamic a activitii motorii. Cile extrapiramidale au rol n: - transmiterea mesajelor tonice posturale; - conducerea comenzilor pentru micrile automate i asociate; -coordonarea micrilor semivoluntare sau voluntare. Pentru realizarea acestor funcii sunt create circuite multineuronale: - cortex - nuclei bazali-cortex: controleaz tonusul i intensitatea micrii; - cortex-cerebel-cortex: asigur coordonarea dinamic a micrilor corticale voluntare sau a motricitatii subcorticale; - cortex-formaiunea reticulat-cortex: care intervine n sens activator sau inhibitor n realizare unor acte motorii, etc. Comenzile pentru motricitatea automat nnscut (masticaie) sau ctigat (mers, scris, cntat la instrumente) se declaneaz n ariile prefrontale 6,8, aria temporal 21 i nucleii bazali. Cile oculomotorii Cile oculomotorii au origine dubl, somatic i vegetativ, intervin n coordonarea micrilor conjugate ale capului i ochilor ca i n reactivitatea i acomodarea pupilar.
INTEGRAREA FUNCIONAL A ACTIVITILOR MOTORII
Realizarea actelor motorii voluntare necesit integritatea morfofuncional a tuturor structurilor nervoase i a efectorilor musculari. In realizarea actelor motorii voluntare se parcurg mai multe etape: - conceperea sau "proiectarea" cortical a micrii; - stabilirea secvenelor micrii i prospectarea condiiilor n care se efectueaz; - transmiterea comenzilor la sistemul cfcctor neuro-muscular, adecvat scopului micrii; - controlul temporo-spaial al execuiei micrii; - reglarea i adaptarea forei de contracie musculare la nevoile actului motor; - modelarea activitii motorii voluntare la condiiile aprute pe
parcursul desfurrii aciunii; - stabilirea continurii sau ntreruperii activitii motorii. Deci, pe tot parcursul activitii motorii voluntare, scoara cerebral asigur coordonarea privind declanarea, susinerea i ncetarea actului motor. Prin intermediul scoarei cerebrale, unele deprinderi motorii capt un caracter de automatism sau "stereotip dinamic" ca urmare a repetrii actului motor de un numr foarte mare de ori. Prin conexiunile dintre diferitele arii corticale motorii, senzitivosenzoriale se realizeaz acte motorii specializate cu caracter praxic (scrisul, vorbirea, cntatul, etc).
8.4 CORTEXUL MOTOR I SISTEMUL ORO-FACIAL
Analiznd reprezentarea motorie cortical a corpului, aa-zisul "homun-culus motor" constatm c proiecia este rsturnat i existena unor zone motorii excesiv de mari corespunztoare frunii, ochilor, buzelor, maxilarului, limbii, deglutitiei i a zonelor responsabile de vocalizare. Reprezentarea nu este n raport cu mrimea segmentului ci cu importana funcional, fiind cu att mai ntins, cu ct funcia motorie este mai fin i mai difereniat. n partea inferioar a circumvoluiei frontale ascendente (aria 4 Brodman) se gsete: - aria primar masticatorie; - aria secundar masticatorie (lateral de cea primar) ce controleaz muchii feei i muchii masticatori de aceeai parte; - aria suplimentar (medial de girus cinguli) care controleaz micrile mandibulei n timpul fonaiei i armonizeaz micrile mandibulei cu micrile capului, feei, gtului. Scoara motorie declaneaz dinamica mandibular, a limbii, a laringelui pentru realizarea micrilor specifice n timpul masticaiei, deglutitiei, respectiv fonaiei i vorbirii. Impulsurile din scoara motorie prin cile piramidale, fibrele corticonucleare, controleaz muchii extremitii cefalice i ai sistemului oro-facial. Fibrele cortico-nucleare au un traiect direct sau ncruciat, pe linia median, la nivelul trunchiului cerebral. Dup ncruciare, fibrele corticonucleare ajung la nucleii motori ai nervilor cranieni, unde fac sinaps cu neuronii motori ai trigemenului, facialului, hipoglosului. Neuronii motori se distribuie, apoi, muchilor corespunztori. Cile descendente cortico-nucleare au pe traiectul lor doi neuroni. - primul neuron cu origine n scoar; - al doilea neuron, localizat n nucleii motori ai nervilor cranieni. Remarcm, n nucleii motori ai nervilor cranieni, o anumit somatotopie a neuronilor ce induc contracia unor anumite fibre musculare, bine delimitate. i n cadrul activitii motorii a sistemului oro-facial se pot realiza unele
stereotipuri masticatorii. Controlul nervos realizat de scoara cerebral, este ajustat, "programat" conform cu morfologia sistemului oro-facial i cu necesitile funcionale, stabilindu-se un stereotip de masticaie: de tocare, de frecare, de propulsie, mixt. Cu punct de plecare de la receptorii parodontali se adapteaz fora de contracie a muchilor masticatori dependent de consistena alimentului i experiena personal, ntrerupndu-se, la nevoie, masticaia. Controlul cortexului cerebral este absolut necesar pentru finalizarea actului masticator, de realizare a bolului alimentar.
9.ELECTROMIOGRAFIA (EMG)
EMG reprezint totalitatea procedeelor obiective de studiu ale activitii electrice musculare n cursul contraciei reflexe sau voluntare. Traseul obinut prin nscrierea grafic a biopotenialelor generate n muchi, n activitate, reprezint electromiograma. In 1851, Dubois Reymond a msurat curenii emii de un muchi n contracie, realiznd prima detectare electromiografic, ulterior Piper (1907-1912) folosind galvanometrul cu coard nregistreaz primele trasee electromiografce. Biopotenialele musculare, n timpul contraciei se pot nregistra la nivelul unei uniti motorii sau a muchiului n ansamblu. Potenialele de aciune musculare genereaz cureni electrici, de amplitudine redus care se propag la distan de locul de producere permind detectarea i culegerea lor cu ajutorul aparatului specializat numit electromiograf. Electromiograful, n prinicipiu, este format din: - electrozi ce culeg potenialele de aciune musculare; - un sistem de amplificare a biopotenialelor; - im sistem de nregistrare i/sau afiare a rezultatelor. Pentru culegerea biopotenialelor se utilizeaz urmtoarele tipuri de electrozi: - electrozi de suprafa: sunt plci metalice, de dimensiuni variabile, plasate pe tegumente, deasupra muchiului examinat, ce ofer informaii referitoare la activitatea electric global a muchiului; - electrozi de tip ac - permit cercetarea unor poriuni foarte mici de muchi i studiul potenialelor de unitate motorie. Culegerile pot fi efectuate prin derivaii monopolare, bipolare sau multipolare. Folosirea acelor-electrod are inconvienentul c d fenomene dureroase, care scad n intensitate dac pacientul realizeaz o bun relaxare muscular. Electrozii de suprafa pentru muchii implicai n funcionalitatea aparatului dento-maxilar sunt plasai astfel: - pentru fasciculele anterioare ale muchilor temporali: la 2 cm superoextern fa de unghiul extern al orbitei; - pentru maseteri, n zona unghiului mandibulei unde se palpeaz cel
mai bine muchii contractai; - pentru pntecele anterioare ale digastricilor: paramedian, submentonier; - penru sternocleidomastoidieni: la jumtatea lungimii muchiului. nregistrarea semnalelor culese de la muchi i amplificate corespunztor se poate realiza: - grafic, prin conectarea amplificatorului la un sistem de penie inscriptoare; - folosind ecranul unui osciloscop conectat la sistemul de amplificare; - pe band magnetic "audio", nregistrndn-se semnalele amplificate ia vitez mare; - cu ajutorul unor sisteme computerizate care digitalizeaz semnalul, sto-cnd traseele sub forma unor fiiere grafice n memoria de mas a calculatorului. 9.1. INTERPRETAREA (ANALIZA) TRASEELOR EMG: I. - durata potenialelor de unitate motorie (UM) reprezint timpul necesar nscrierii electromiografice a modificrilor electrice ce au loc ntr-o unitate motorie cu ocazia unei singure contracii. Durata potenialului de UM d indicaii privind: - capacitatea cuplului neuro-muscular de a sincroniza ntr-o anumit perioad de timp activitatea contractila a tuturor fibrelor musculare dependente; - ritmul desfurrii fenomenelor electrice i chimice din fiecare fibr muscular n parte. Valorile normale ale duratei potenialelor de UM, variaz, dup diferii autori, ntre 3-6 msec., sau 4-8 msec. considerndu-se o valoare medie de 5 msec. Pentru muchii feei valorile normale sunt 5-6 msec. Durata potenialului de UM. prezint variaii dependente de vrst, temperatur, stare de oboseal; II. - amplitudinea unui potenial de UM exprim fora de contracie a fibrelor musculare a cror activitate este culeas de electrod. Amplitudinea unui singur potenial relev fora de contracie a grupului de fibre musculare cu care electrodul vine n contact. ntruct se culege activitatea electric a unui grup de fibre musculare,, amplitudinea potenialului cules nu reprezint curentul de aciune a unei singure celule musculare ci cmpul electric realizat de nsumarea curenilor de aciune ai tuturor fibrelor din grup. De aceea, acul-electrod trebuie nserat chiar n centrul acestui cmp i astfel amplitudinea potenialului este maxim. Valorile normale ale amplitudinii potenialelor de UM variaz ntre 100500 uV i chiar 2 mV (fig. 22). III. - forma potenialului de UM: poate fi bi-, tri-, sau polifazic, Apariia potenialelor polifazice se datorete: - poziiei defectuoase a electrodului plasat n zona de ntreptrundere. : fibrelor musculare a dou sau trei UM vecine; - sincronizrii defectuoase a fibrelor musculare ale UM din care se
culege axonului;
- diferenelor n lungimea i diametrul ramurilor terminale ale
- ntrzierilor sinaptice la plcile motorii din cadrul aceleiai UM; - vitezei diferite de propagare a potenialului de aciune n fibrele:muculare. IV - frecvena potenialelor de UM: pericarionul neuronului motor periferic descarc gradat poteniale de aciune n raport cu intensitatea activitii funcionale a neuronului. Frecvena acestor poteniale, care se succed repetitiv, reflect intensitatea stimulului sosit de la neuronii motori centrali. Frecvena de descrcare a UM crete paralel cu efortul muscular. Efortul muscular este controlat prin mecanisme de feed-back declanate de contracia muchiului, n fusurile neuro-musculare i organele tendinoase Golgi. n timpul unei contracii maximale, ritmul de descrcare al motoneuronului crete la 60-100 cicli/sec n cteva zeci de milisecunde. In contracia susinut ritmul de descrcare este de 20 cicli/sec. Pe traseul EMG frecvenele de descrcare se reflect sub form de poteniale ce se succed foarte rapid i la intervale mici de timp. Aceast cretere a activitii pe unitatea de timp reprezint fenomenul de sumaie temporal. La sumaia temporal se adaug sumaia spaial (se nregistreaz simultan i progresiv i activitatea unitilor motorii vecine).
9.2. ACTIVITATEA EIVIG LA GRADAIA CONTRACIEI MUSCULARE O contracie muscular uoar determin pe traseul EMG un traseu simplu, care este constituit din poteniale de aciune mono- sau bifazice, cu amplitudinea de 200-400 \jlV, durata de 3-4 msec. i frecvena de 4-10 cicli/sec. Pe msura creterii forei de contracie, se produce i mbogirea traseului EMG, graie fenomenelor de sumaie temporal i spaial. La o contracie moderat, activitatea unitii motorii aflate chiar la vrful acului-electrod nscrie pe traseul EMG o succesiune de biopoteniale cu frecvena i amplitudinea mai mari dect pe traseul simplu, ajungnd la 15-25 cicli/sec. i amplitudine de 500-600 u.V. Alturi de aceste poteniale pe traseul EMG mai apar i activitile electrice ale unitilor motorii vecine. Acesta este traseul intermediar. Intricarea potenialelor de aciune ale unitilor motorii se accentueaz progresiv; amplitudinea lor crescnd paralel cu mrirea forei de
contracie, activitatea UM de Ia vrful acului-electrod nu mai poate fi distins de activitile unitilor motorii vecine. n acest mod se constituie traseul interferential. Amplitudinea general a traseului interferential este de 10001200 u,V, dar poate fi considerat normal pn Ia 2000 u.V. Examenul EMG efectuat unui individ normal, n stare de repaus muscular nu deceleaz existena unei activiti bioelectrice, pe ecranul aparatului reproducndu-se aspectul de traseu izoelectric. Exist ns i activiti electrice spontane care nu au semnificaie patologic i care trebuie foarte bine cunoscute, pentru a evita regretabile greeli de diagnostic: - potenialele electrice spontane de inserie; aceste activiti spontane apar la implantarea electrodului n muchi sau la modificarea poziiei acestuia; - potenialele de nerv, apar dac la implantarea electrodului se lezeaz o fibr nervoas intramuscular. 9.3. VARIAII FIZIOLOGICE ALE TRASEELOR EMG Traseele EMG pot prezenta variaii n funcie de o serie de factori fiziologici, variaii de care trebuie s se in seama la interpretarea traseelor. Astfel de factori sunt: vrsta, temperatura, oboseala muscular, circulaia sanguin, etc. Vrsta: se consider c amplitudinea i durata potenialelor de UM prezint valori cu att mai mici, cu ct subiecii sunt mai tineri. Amplitudinea potenialelor UM crete cu vrsta, n primii 2 ani de via diferena fiind de -35% faa de aduli. Se pare c durata scurt a potenialelor de aciune a unitilor motorii infantile se datorete suprafeei reduse a sinapselor neurornusculare la aceast vrst. Temperatura corpului constituie, de asemenea, un factor care influeneaz buna desfurare a activitii neurornusculare. Este necesar ca temperatura din laboratorul EMG s fie 23-24, cnd se realizeaz nregistrarea. Cu ct temperatura este mai sczut, cu att procentajul potenialelor polifazice este mai crescut, amplitudinea general a traseului este mai sczut, iar capacitatea de a realiza interferena traseului EMG la contracie maximal este diminuat. Pentru fiecare grad de temperatura n minus, viteza de conducere motorie scade cu 3 m/sec. Oboseala muscular: fenomenul de oboseal musculara influeneaz (ca i hipotermia) funcionalitatea neuromuscular, n sensul c un muchi obosit genereaz trasee EMG cu amplitudine mai sczut. Pe traseele de tip simplu se constat scderea frecvenei biopotenialelor pe unitatea de timp, iar ritmul de succesiune a descrcrilor devine neregulat. De asemenea, se constat o tendin de sincronizare a activitilor neuronale, ceea ce creeaz dificulti n realizarea traseului interferential. Att hipotermia ct i oboseala muscular influeneaz funcionalitatea unitilor motorii prin intermediul tulburrilor de debit circulator muscular.
9.4. EXPLORAREA ELECTROMIOGRAFIC A SISTEMULUI ORO-FACIAL 9.4.1. EMG-ul musculaturii feei Activitatea EMG a muchilor feei reflect particularitile morfofuncionale ale acestor muchi, de dimensiuni reduse i cu inserii pieloase. Parametri potenialelor de UM prezint amplitudinea redus, durata scurt i forma adesea polifazic. Traseele sunt bogate, interfereniale. Examenul EMG al musculaturi feei i are indicaia n paraliziile i contracturile faciale. Astfel, n paraliziile faciale, examenul EMG efectuat la 15 zile. dup debutul bolii constat, de obicei, o bogat activitate electric spontan, sub forma de poteniale de fibrilaie i adesea poteniale lente de denervare. Examenul EMG n cazurile de paralizie facial mai veche de 8 sptmni prezint o deosebit valoare n ceea ce privete indicaia operatorie (eliberarea nervului n 1/3 distal a apeductului Falloppe). n contracturile faciale EMG d posibilitatea n primul rnd s se stabileasc originea postparalitic sau primitiv a contractrii. Astfel, n hemispasmele faciale postparalitice se constat existena - de obicei la toi muchii hemifeei - a unei activiti electrice spontane abundente, pe care se suprapun bufeuri sincrone de tip miotonic, cu frecvena de 180-250 cicli/sec. La micrile voluntare apare un traseu srac, cu numeroase poteniale polifazice i cu numeroase sinkinezii. Srcia traseului este cu att mai mare, cu ct elementul paralitic este mai accentuat. n hemispasmele faciale primitive activitatea electric spontan const din bufeurile repetitive sincrone descrise, precum i din potenialele polifazice care survin pe toate derivaiile musculare i care traduc secusele clonice, vizibile clinic. La efectuarea micrilor voluntare, apar trasee caracterizate prin marea lor bogie n biopoteniale, care ating repede interferena i prin extinderea activitii electrice a muchiului care se contract, asupra muchilor nesolicitai n micare. 9.4.2. EMG n musculatura velopalatin, faringian i laringian
Aspectul diferit al traseelor EMG derivate din musculatura velopalatin, faringian i laringian constituie expresia particularitilor morfofiziologice i filoontogenetice ale acestei musculaturi. Musculatura velo-palato-faringo-laringian este prevzut cu receptori de sensibilitate proprioceptiv prin intermediul cror se execut un control miotatic de tip feed-back asupra funciilor de deglutiie i fonaie, care devin astfel acte automate dar contiente, integrate n schema corporal a individului.
Potenialele de aciune ale unitilor motorii au anumite particulariti. Astfel: amplitudinea potenialelor de UM este redus - la muchii intrinseci ai laringelui este n general de 100-300 uV i crete la 500 uV n muchii tiroaritenoidian i crico-tiroidian. n muchii velo-palatini i faringieni amplitudinea potenialelor de UM atinge n timpul deglutiiei 400 uV. Durata este de asemenea foarte scurt, ntre 2 i 4 msec. Forma este n general bi- i trifazic. Frecvena potenialelor de aciune a unitilor motorii difer de la muchi la muchi i n funcie de activitatea pe care o exercit. Astfel, la muchii velopalatini succesiunea potenialelor are frecvena de 10-20 cicli/sec. n timpul fonaiei i ajunge la 20-40 cicli/sec. n timpul deglutiiei. n muchii laringelui i cu deosebire n muchiul vocal (tiro-aritenoidian) frecvena crete pn la 60 cicli/sec. n timpul vorbirii i pn la 100 cicli/sec. n timpul cntatului obinuit. Traseele EMG ale musculaturii velo-palatine: n stare de repaus nu exist activitate spontan, pe trasee aprnd linia izoeleetric. La activitatea fonatorie, frecvena potenialelor electrice realizeaz doar trasee de tip intermediar, cu amplitudine maxim de 300-350 pV i durat total de 10001500 msec. Traseele EMG ale constrictorului superior al faringelui: n stare de repaus se nregistreaz linia izoeleetric. Activitatea muchiului ncepe cu ocazia deglutiiei, cnd se nscrie un traseu interferenial. Traseele EMG ale constrictorului mijlociu al faringelui: in repaus apare linia izoeleetric. Fonaia nu determin apariia de biopoteniale n acest muchi. Activitatea principal apare cu ocazia deglutiiei, cnd se nregistreaz traseul interferenial cu aspect global lenticular, ca i la muchii velo-palatini i constrictorul faringian superior. La nivelul constrictorului mijlociu al faringelui activitatea electric caracteristic apare n timpul reflexului de vom, trecerea undelor antiperistaltice fiind marcat de nscrierea unor faze scurte de traseu interferenial cu amplitudine de 130-300 pV i durat de 300-400 msec., precedate i urmate de unde ample i neregulate, care traduc deplasrile electrodului cu ocazia micrilor puternice ale muchiului n totalitate. Traseele EMG ale muchilor tiro-aritenoidieni: cei doi muchi simetrici se investigheaz simultan, de aceea este necesar un aparat EMG cu dou canale. La m. tiro-aritenoidieni n stare de repaus funcional este foarte greu, chiar excepional, s se obin linie izoeleetric. Exist aproape permanent o activitate legat de respiraie, de deglutiie etc., care genereaz aspecte neregulate de tip simplu sau intermediar foarte srac, cu poteniale de 100-150 uV i durat de 1,5-2 msec. n timpul fonaiei, pe traseele EMG se pot nregistra dou feluri de activiti. In general apare o activitate de tip intermediar bogat sau interferenial, constituit din poteniale cu durat de 2-5 msec., amplitudine de 200-400 uV, form bi- i trifazic. Aceasta este activitatea bioelectric muscular, n care potenialele reproduc homoritmic succesiunea de stimuli sosit de la centrii nervoi. Uneori apare o activitate cu aspect repetitiv, pseudomiotonic, care se datorete - dup Isch - trecerii jetului de aer peste corzile vocale i vibrrii acestora homoritmic cu sunetul emis. Muchii tiro-aritenoizi intr n activitate nu numai n timpul vorbirii rostite ci i n timpul vorbirii interioare (silent
speech). La invitaia de a rosti "n gnd" propoziiunea spus mai nainte "cu voce tare", se nregistreaz trasee asemntoare din punct de vedere al aspectului general, ns cu amplitudine i frecven mai mici. Examenul EMG al musculaturii velo-palatine, faringiene i laringiene este important n cazul diferitelor afeciuni ale laringelui, mai ales n paraliziile laringiene, n diagnosticul unor sindroamc mioclonice, n fonasteniile psihogene, n afoniile isterice, pentru stabilirea diagnosticului diferenial al afeciunilor bulbare i pseudobulbare, al unor forme de mutism, afazii sau disfonii de origine miopatic, miastenic, prin atrofie, agenezie sau fibroz muscular.
9.4.3. EMG n masticaie Muchii implicai n masticaie (m. ridictori i cobortori ai mandibulei) realizeaz o contracie asincron i asimetric. Fora maxim de contracie apare dup maximul evideniat pe EMG, la 0,09 secunde de la stabilirea contactului n poziia de intercuspidare maxim i dureaz 0,11 secunde. Amplitudinea maxim a traseelor EMG la nivelul muchilor maseteri i temporali se nregistreaz n PIM. n masticaie, amplitudinea biopotenialelor variaz n funcie de muchi, valoarea medie procentual fiind: - la muchii temporali: 35%: - la fasciculul profund al muchilor maseteri: 86%; - la fasciculul superficial al muchilor maseteri: 86%. Potenialele EMG nregistrate au ntotdeauna amplitudine mai mare pe partea lucrtoare la nivelul fasciculelor anterioare ale muchilor temporal i fasciculului superficial al maseterului. In masticaia la edentai: - se reduce activitatea electric n muchii maseteri; - cresc potenialele EMG n muchii circumorali. Protezarea corect determin revenirea la activitatea electric uzual. Poziia de postur a mandibulei ce realizeaz spaiul de inocluzie fiziologic prezint variaii, la acelai individ, n funcie de: - vrst; - poziia corpului: clinostatism, ortostatism; - efort fizic; - starea de veghe/somn; - stri patologice - algii. Poziia de postur a mandibulei este realizat prin uoara contracie a muchilor ridictori evideniat prin prezena potenialelor EMG. 9.4.4. EMG n deglutiie n deglutiie contracia muscular este sincron i simetric. Asimetria n contracia maseterilor este considerat semnificativ pentru disfuncia temporo-mandibular. Edentaia total protezat duce, dup o perioad de acomodare ndelungat de circa doi ani la creterea activitii EMG n timpul deglutiiei numai la nivelul orbicuiarului gurii.
BIBLIOGRAFIE SELECTIVA 1. ADAMS M.M. - Down's syndrome: recent trends in the Uniled States. 758-760 2. AU G.N.; LAUNDL T.M.; WALLACE KT,.; DECARLE D.J.; COOK I.J stimulation on the normal pharyngeal swallow responsc [sec co Wintcr 1996, 11 (1), 2-8 3. ANDRONESCU A. - Anatomia copilului, Ed.didactica i pedagogic Bi 4. ARONSSON A.M.; LIND B.; NYLANDER M.; NORDBERG M. - 1 mercury. Biol Met, 1989, 2 (1), 25-30 5. ASERO R.; MASSIRONI F.; VELATI C. Dctection of prognostic fac syndrome in patients with birch pollen hypersensitivity. J Allerj 1996, 97 (2), 6. ASHER McDADE; SHAW W.C. - Currcnt cleft lip and polalc mana! Kingdom.Br.,1.Plastic Surg.,1990, 43, 318-321 7. ATKINSON M.E., WHITE EH. - Principles of anatomy and oral anaton Churchill Livingslone, ED1NBURGH, LONDON, 1992 8. BARRETT A.W.; SCULLY C. - S100 protcin in oral biology and pat Med, Nov 1994, 23 (10), 433-40 9. BERGLUND A. - Release of mercury vapor from dental amalgam. Swc 85 1-52 10. BERKOVITZ B.K.B. - How teelh erupt. Dental update, 1990, 17. 20f 11. BERRIDGE M.J, IRVINE R.F. - Inositol phosphatcs and cell signallin 341, 197-205 12. BERRIDGE M.J, IRVINE R.F. - Inositol triphosphatc, a novei second signal transduction. Nature Lond., 1984, 312, 315-321 13. BERRIDGE M.J. - Calcium osctllations, .1. Biol. Chem., 1990, 265, i 14. BHASKAR S.N. - ORBANS Oral Histology and Embryology, eleve Mosby Year Book, 1990. 15. BJORK A., SKIELLER V. - Facial development and tooth eruption. Ai 62, 4, 339 16. BLANCHETTE M.E.; NANDA R.S.; CURRIER G.F.; GHOSH J.: NA dinal cephalometric study of the soft lissue profile of short- ar from 7 to 17 ycars. Am J Orthod Dcntofacial Orthop, Feb 19(; 17. BOBOC GHEORGHE - Aparatul dento-maxilar, Formare i dezvolta Medical, Bucureti, 1996 18. BRATU D NICULESCU V, STANCU GH., URTIL E. - Net morfologie clinic, 1993, Timioara 19. BRATU D. i colab. - Aparatul dento-maxilar. Dale de morfologic Helicon, Timioara, 1997.
S-ar putea să vă placă și
- Glandele Anexe Ale Tubului DigestivDocument17 paginiGlandele Anexe Ale Tubului DigestivAndreea Diana Drăgoi100% (1)
- Anomalii DentareDocument103 paginiAnomalii DentareAnisoara100% (1)
- DEGLUTIŢIADocument7 paginiDEGLUTIŢIAmogoanta madalinaÎncă nu există evaluări
- Morfologia Functionala A Sistemului StomatognatDocument24 paginiMorfologia Functionala A Sistemului StomatognatAlex Popovici100% (2)
- Examen ParodontologieDocument35 paginiExamen ParodontologieMl BeÎncă nu există evaluări
- Anatomia Chirurgicala A FeteiDocument16 paginiAnatomia Chirurgicala A FeteiZmeu Cristina100% (2)
- Anatomia Parodontiului .Ciurdila DanDocument21 paginiAnatomia Parodontiului .Ciurdila Danciurdila100% (1)
- Etiologia Anomaliilor Dento-MaxilareDocument30 paginiEtiologia Anomaliilor Dento-MaxilareRoxana Lupu100% (1)
- Sinusul MaxilarDocument29 paginiSinusul MaxilarYoeli SmithÎncă nu există evaluări
- Examen ParodontologieDocument122 paginiExamen ParodontologieAlleste Oren100% (3)
- Osul MaxilarDocument4 paginiOsul MaxilarAdina Neagoe100% (1)
- Pulpa DentaraDocument4 paginiPulpa DentaraTheodoros Katsipanos100% (1)
- Pulpa DentaraDocument55 paginiPulpa DentaraEmilia Gemanaru100% (1)
- Aaa ExamenDocument44 paginiAaa ExamenLilia LiliaÎncă nu există evaluări
- Anatomia DinteluiDocument7 paginiAnatomia DinteluiSuzana Soozy100% (1)
- Elaborarea 7 ChirurgieDocument8 paginiElaborarea 7 Chirurgietatiana0ostahiiÎncă nu există evaluări
- Sectia de Stomatologie Preventiva: Organizarea, Regimul de Lucru, Utilarea Si InstrumenteDocument19 paginiSectia de Stomatologie Preventiva: Organizarea, Regimul de Lucru, Utilarea Si InstrumenteIrina LutcanÎncă nu există evaluări
- Parodontiul MarginalDocument34 paginiParodontiul MarginalCristiana Leca100% (4)
- Tartrul Dentar. Compoziţia Chimică. Mecanismul de Formare A Tartrului DentarDocument20 paginiTartrul Dentar. Compoziţia Chimică. Mecanismul de Formare A Tartrului DentarGabriela CebanÎncă nu există evaluări
- Flegmon 2013 Prel.1Document48 paginiFlegmon 2013 Prel.1Grigori VasileÎncă nu există evaluări
- 1.parte GeneralaDocument14 pagini1.parte GeneralaAda Balo100% (1)
- Anatomia Radiologică A Articulatiei Temporo-Mandibulare: Rezident Stela SDocument33 paginiAnatomia Radiologică A Articulatiei Temporo-Mandibulare: Rezident Stela SstelaÎncă nu există evaluări
- Parodontiul ReferatDocument4 paginiParodontiul ReferatDragos FloreaÎncă nu există evaluări
- Curs 7 Cavitatea BucalăDocument20 paginiCurs 7 Cavitatea BucalăToropoc GeorgeÎncă nu există evaluări
- Elaborarea Metodică Nr. 1Document6 paginiElaborarea Metodică Nr. 1Daniela100% (2)
- Elaborarea Practica NR 14Document5 paginiElaborarea Practica NR 14Melania ZorilaÎncă nu există evaluări
- Nervul FacialDocument7 paginiNervul FacialFamilia Macovei Haralambie-AndreeaÎncă nu există evaluări
- Parodontologie Preclinică, Lucrarea Practică Nr. 1, ParadontologieDocument5 paginiParodontologie Preclinică, Lucrarea Practică Nr. 1, ParadontologieStudentulÎncă nu există evaluări
- Ganglionilor Limfatici Ai CapuluiDocument52 paginiGanglionilor Limfatici Ai CapuluiDana Roxana100% (2)
- Etiologia Cariei DentareDocument9 paginiEtiologia Cariei DentareOctavian CalaraşÎncă nu există evaluări
- Referat Pe Tema: Explorari Paraclinice Utilizate Pentru Depistarea Afectiunilor Oro-Maxoli-FacialeDocument14 paginiReferat Pe Tema: Explorari Paraclinice Utilizate Pentru Depistarea Afectiunilor Oro-Maxoli-FacialeMarin DulghieruÎncă nu există evaluări
- TGDocument8 paginiTGLilia LiliaÎncă nu există evaluări
- Caria DentaraDocument2 paginiCaria DentaraArtiom MutavciÎncă nu există evaluări
- Pulpa DentaraDocument30 paginiPulpa DentaraOana CostanÎncă nu există evaluări
- Lucrare DiplomaDocument70 paginiLucrare DiplomaCătălin Nicolae100% (1)
- OMF1Document20 paginiOMF1tatiana0ostahiiÎncă nu există evaluări
- Topografia Regiunii Anterioare A Fetei, Proectia OrificilorDocument17 paginiTopografia Regiunii Anterioare A Fetei, Proectia OrificilorDima CekalÎncă nu există evaluări
- Materialele de Obturaţie de Bază de Ultimă OrăDocument26 paginiMaterialele de Obturaţie de Bază de Ultimă OrăNicolae Grigoras100% (1)
- Elaborarea Metodica Nr. 1Document5 paginiElaborarea Metodica Nr. 1lozovanudÎncă nu există evaluări
- AT Tema 10 Incizii RationaleDocument31 paginiAT Tema 10 Incizii RationalecomiasanÎncă nu există evaluări
- Abcesul ParotidianDocument12 paginiAbcesul ParotidianAnastasia PopaÎncă nu există evaluări
- Substante Anestezice Locale Si Corective VasoconstrictoareDocument13 paginiSubstante Anestezice Locale Si Corective VasoconstrictoareElena GiuscaÎncă nu există evaluări
- Elaborarea Metodică Nr.3Document13 paginiElaborarea Metodică Nr.3Ojog100% (2)
- Anestezia Nervului Alveolar SuperoDocument1 paginăAnestezia Nervului Alveolar SuperoLucia PopescuÎncă nu există evaluări
- Fiziologie - Medicina DentaraDocument59 paginiFiziologie - Medicina Dentaragabri1001Încă nu există evaluări
- Tema-7 Particularităţile de Varstă. Anomaliile de Dezvoltare A Masivului FacialDocument6 paginiTema-7 Particularităţile de Varstă. Anomaliile de Dezvoltare A Masivului FacialCatalin GodovanciucÎncă nu există evaluări
- Redactat 2 OmfDocument29 paginiRedactat 2 OmfTheodor Tricket100% (1)
- Familiarizarea Studen Ților Cu Componența Catedrei - Particularitățile Chirurgiei Dento-Alveolară Și Secției de Chirurgie OMFDocument30 paginiFamiliarizarea Studen Ților Cu Componența Catedrei - Particularitățile Chirurgiei Dento-Alveolară Și Secției de Chirurgie OMFMarica SirghiÎncă nu există evaluări
- Tema 9: Anatomia Nervului Trigemen.: Raspuns Importanța Temei 1. A 2. B 3. C 4. DDocument10 paginiTema 9: Anatomia Nervului Trigemen.: Raspuns Importanța Temei 1. A 2. B 3. C 4. DalexÎncă nu există evaluări
- Fiziologia Mediului BucalDocument15 paginiFiziologia Mediului BucalciposÎncă nu există evaluări
- FizioDocument79 paginiFizioSebastian BarbulescuÎncă nu există evaluări
- FSOF (AutoRecovered) ...Document11 paginiFSOF (AutoRecovered) ...AndreeaÎncă nu există evaluări
- Parodonto ExamDocument117 paginiParodonto ExamgalinaosocÎncă nu există evaluări
- CAVITATEA ORALĂ, Amigdale, Glande SalivareDocument19 paginiCAVITATEA ORALĂ, Amigdale, Glande SalivareJowan YassinÎncă nu există evaluări
- Saliva CursDocument105 paginiSaliva CursAna-Maria CostacheÎncă nu există evaluări
- Tema 1Document10 paginiTema 1VictorÎncă nu există evaluări
- LP 6 - 7 - Fiziologia Aparatului DigestivDocument68 paginiLP 6 - 7 - Fiziologia Aparatului DigestivioanaÎncă nu există evaluări
- Fiziologia DintilorDocument31 paginiFiziologia DintilorCorina Caragacean100% (2)
- Modul I NutritieDocument204 paginiModul I Nutritieppappi12Încă nu există evaluări
- Biologie - Clasa A 10-ADocument28 paginiBiologie - Clasa A 10-ACristigaming29Încă nu există evaluări