S-ar putea să vă placă și
- Analizatorul Olfactiv Si GustativDocument5 paginiAnalizatorul Olfactiv Si GustativBogdan CiochinaÎncă nu există evaluări
- 4-Spatiul PopliteuDocument4 pagini4-Spatiul PopliteuMadalina NinaÎncă nu există evaluări
- Citoscheletul MembranarDocument1 paginăCitoscheletul MembranarCristina PopÎncă nu există evaluări
- Fiziologia Analizatorului Vestibular Utricula Sacula Şi Canalele SemicirculareDocument1 paginăFiziologia Analizatorului Vestibular Utricula Sacula Şi Canalele SemicirculareDiana LavricÎncă nu există evaluări
- Polaritatea Tesutului EpitelialDocument7 paginiPolaritatea Tesutului EpitelialAriela SăvuţÎncă nu există evaluări
- Prezentare AnticorpiDocument15 paginiPrezentare AnticorpiOanaDorofteÎncă nu există evaluări
- Tesutul EpitelialDocument47 paginiTesutul EpitelialValeria JentimirÎncă nu există evaluări
- Curs 1 Tesutul EpitelialDocument11 paginiCurs 1 Tesutul EpitelialVerginia ShuganiÎncă nu există evaluări
- Analizatorul VizualDocument2 paginiAnalizatorul VizualPaul DorcuÎncă nu există evaluări
- Increngatura Celenterate PROIECTDocument11 paginiIncrengatura Celenterate PROIECTVeronica PlamadealaÎncă nu există evaluări
- Tesut Muscular 26324Document111 paginiTesut Muscular 26324Galina BotnariÎncă nu există evaluări
- Nerv SinapsaDocument57 paginiNerv SinapsaDoina Moisei100% (1)
- Țesuturile Musculare Sunt Adaptate Funcției de ContracțieDocument3 paginiȚesuturile Musculare Sunt Adaptate Funcției de ContracțieAndriana PrisacariÎncă nu există evaluări
- Neurocraniu IDocument3 paginiNeurocraniu Idaniela mortuÎncă nu există evaluări
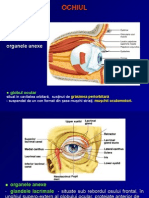
- OchiulDocument31 paginiOchiulRadu Stefan MaricaÎncă nu există evaluări
- Reglarea Nervoasa Si Umorala A Activitatii UrineiDocument20 paginiReglarea Nervoasa Si Umorala A Activitatii UrineiAdelina Cadar100% (1)
- Teme - Anatomia Analizatorului VizualDocument34 paginiTeme - Anatomia Analizatorului Vizualcomanflavius2262Încă nu există evaluări
- Fiziologie Si Patologie-ColocviuDocument25 paginiFiziologie Si Patologie-ColocviugencimoravaÎncă nu există evaluări
- 4) Variatii de Tesuturi ConjunctiveDocument5 pagini4) Variatii de Tesuturi ConjunctiveIonut Valentin StanciuÎncă nu există evaluări
- Embriologie Curs 5 Saptamana 3Document71 paginiEmbriologie Curs 5 Saptamana 3LumyÎncă nu există evaluări
- Teste Cu Imagini Sem II 2017prima TotalizareDocument28 paginiTeste Cu Imagini Sem II 2017prima TotalizareAndr3 w00DÎncă nu există evaluări
- Generalitati Sist OsosDocument59 paginiGeneralitati Sist OsosNoémi ShepherdÎncă nu există evaluări
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Muşchii Gatului-Sternocleidomastoidian, Scaleni, Infrahiodieni+M.PrevertebraliDocument3 paginiMuşchii Gatului-Sternocleidomastoidian, Scaleni, Infrahiodieni+M.PrevertebraliBogdan100% (1)
- Sistemului NeuromuscularDocument54 paginiSistemului NeuromuscularStefi MaryÎncă nu există evaluări
- Citologie MembranaDocument33 paginiCitologie MembranaOsman Daniela100% (1)
- Hormoni 4 PDFDocument61 paginiHormoni 4 PDFAlina MoraruÎncă nu există evaluări
- Analizatorul Acustico VestibularDocument34 paginiAnalizatorul Acustico VestibularAnghel Mihaela-Andreea100% (1)
- Sistemul EndocrinDocument65 paginiSistemul EndocrinMariaBoicuÎncă nu există evaluări
- Anatomie Curs 3 29.10.2013gametogeneza, Spermatogeneza, Spermatozoidul, Etapele Spermatogenezei, Caracterele Fiziologice Ale Spermatozoidului Lichidul SpermaticDocument3 paginiAnatomie Curs 3 29.10.2013gametogeneza, Spermatogeneza, Spermatozoidul, Etapele Spermatogenezei, Caracterele Fiziologice Ale Spermatozoidului Lichidul Spermaticdaniela_piÎncă nu există evaluări
- Sistem Vascular - 2018Document10 paginiSistem Vascular - 2018rodicaÎncă nu există evaluări
- Acizi NucleiciDocument7 paginiAcizi NucleiciMarci GoreaÎncă nu există evaluări
- Organele Genitale MasculineDocument7 paginiOrganele Genitale Masculineginetta_23roÎncă nu există evaluări
- Teste AnatomieDocument43 paginiTeste AnatomiemariutzaÎncă nu există evaluări
- Curs 6 CitologieDocument43 paginiCurs 6 CitologieRoxana Elena StanciuÎncă nu există evaluări
- Curs 1Document29 paginiCurs 1oanaÎncă nu există evaluări
- Seminar 4Document8 paginiSeminar 4Maryana RoseÎncă nu există evaluări
- b.AciziiNucleici 2018Document59 paginib.AciziiNucleici 2018Dragos PrisacaruÎncă nu există evaluări
- Biomoleculele Si CeluleleDocument26 paginiBiomoleculele Si CeluleleAlexandra PoenaruÎncă nu există evaluări
- Ţesutul Epitelial Glandular (Secretor)Document36 paginiŢesutul Epitelial Glandular (Secretor)Andra AgheorghieseiÎncă nu există evaluări
- Regiunea NucalaDocument4 paginiRegiunea NucalaFlavius Ion NicolaeÎncă nu există evaluări
- BacalaureatDocument35 paginiBacalaureatOprea ViorelÎncă nu există evaluări
- Analizatorul Gustativ Si Vestibular - Curs 12Document5 paginiAnalizatorul Gustativ Si Vestibular - Curs 12Ioana Navalici100% (1)
- Examen AnatomieDocument1 paginăExamen AnatomieGambleru IonutzÎncă nu există evaluări
- Anato CursDocument7 paginiAnato CursCebotari IvanÎncă nu există evaluări
- Biologie Clasa-Xi Auxiliar Configuratia Externa A Emisferelor CerebraleDocument3 paginiBiologie Clasa-Xi Auxiliar Configuratia Externa A Emisferelor CerebraleAlexandra ElenaÎncă nu există evaluări
- Organe de Simt GrigorasDocument16 paginiOrgane de Simt GrigorasScolobiuc Emma ElenaÎncă nu există evaluări
- Rolul Ficatului În Biochimia LipidelorDocument6 paginiRolul Ficatului În Biochimia Lipidelorlaurisca937660Încă nu există evaluări
- Met Intermediar Al Proteinelor Simple Si CromoproteinelorDocument96 paginiMet Intermediar Al Proteinelor Simple Si CromoproteinelorIuliana MîrzaÎncă nu există evaluări
- Curs 4 - CitoscheletDocument46 paginiCurs 4 - CitoscheletAdrian UrsuÎncă nu există evaluări
- Curs 2 InimaDocument20 paginiCurs 2 InimaAttila SzakacsÎncă nu există evaluări
- Biofizica Contracţiei MusculareDocument6 paginiBiofizica Contracţiei MuscularenoruletÎncă nu există evaluări
- 31 Filamentele de ActinaDocument10 pagini31 Filamentele de ActinaAlina Calmuc100% (1)
- BiocelDocument40 paginiBiocelAdelina MusaÎncă nu există evaluări
- Curs 5 BiofizicaDocument9 paginiCurs 5 BiofizicacurteanancaÎncă nu există evaluări
- Curs Nr. 8'Document5 paginiCurs Nr. 8'Co BbÎncă nu există evaluări
- 06 Contractia Musculara 2017-12-04Document11 pagini06 Contractia Musculara 2017-12-04costica16100% (1)
- Ciclu CardiacDocument17 paginiCiclu CardiacCiprian JurjeÎncă nu există evaluări
- FIziologia Aparatului NeuromuscularDocument14 paginiFIziologia Aparatului NeuromuscularLoredana Boghez100% (1)
- Fiziologia MuşchilorDocument74 paginiFiziologia MuşchilorXiusa TurcanÎncă nu există evaluări
- Spectre de VibratieDocument111 paginiSpectre de VibratieEnea DanielaÎncă nu există evaluări
- Prezentare Spectroscopie 8aDocument40 paginiPrezentare Spectroscopie 8aKogaionon25Încă nu există evaluări
- SpectroscopieDocument86 paginiSpectroscopieEnea DanielaÎncă nu există evaluări
- SpectroscopieDocument54 paginiSpectroscopieEnea DanielaÎncă nu există evaluări
- Cursul 2 La BiofizicaDocument8 paginiCursul 2 La BiofizicaStancu CristianÎncă nu există evaluări
- SpectroscopieDocument85 paginiSpectroscopieEnea DanielaÎncă nu există evaluări
- Spectre de VibratieDocument111 paginiSpectre de VibratieEnea DanielaÎncă nu există evaluări
- Jurnal ChirurgieDocument115 paginiJurnal ChirurgieCristi Zamfir100% (1)
- Sistemul - Endocrin - Final - Olimpiada. Final OlimpiadaDocument66 paginiSistemul - Endocrin - Final - Olimpiada. Final OlimpiadaAniela DanielaÎncă nu există evaluări