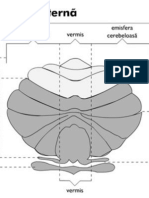
S-ar putea să vă placă și
- Tipuri de ArnDocument6 paginiTipuri de ArnRaluqÎncă nu există evaluări
- Acid RibonucleicDocument4 paginiAcid RibonucleicLarisa HuhuliaÎncă nu există evaluări
- Sinteza Si Secretia CelularaDocument6 paginiSinteza Si Secretia Celularaabcfree2006Încă nu există evaluări
- Prezentare AnticorpiDocument15 paginiPrezentare AnticorpiOanaDorofteÎncă nu există evaluări
- Genetica MolecularaDocument7 paginiGenetica MolecularaClaudia GrosuÎncă nu există evaluări
- 3 Studiul Celulei - Forma Si Dimensiunile CeluleiDocument3 pagini3 Studiul Celulei - Forma Si Dimensiunile CeluleiAndrEea MIronÎncă nu există evaluări
- Poliglucide de RezervaDocument7 paginiPoliglucide de RezervaSh4d0wSt4lk3r50% (2)
- Biologie - Anatomie Si Fiziologoe UmanaDocument8 paginiBiologie - Anatomie Si Fiziologoe UmanaAlice MoisăÎncă nu există evaluări
- Bilanţul Energetic Din Metabolismul GlucidicDocument7 paginiBilanţul Energetic Din Metabolismul Glucidicana1987varmezÎncă nu există evaluări
- Topografia Organelor Şi Sistemelor de OrganeDocument3 paginiTopografia Organelor Şi Sistemelor de OrganeAncuța Rãileanu100% (1)
- ATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialDocument8 paginiATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialPascu Alexandru0% (1)
- Procese MetaboliceDocument14 paginiProcese MetaboliceGabriel Andrei Stan100% (1)
- Metabolismul Materiei ViiDocument9 paginiMetabolismul Materiei ViiInaam BaraziÎncă nu există evaluări
- Nucleii Olivari BulbariDocument2 paginiNucleii Olivari BulbariMihai Alexandru ButnariuÎncă nu există evaluări
- Biologie Clasa A XI-aDocument14 paginiBiologie Clasa A XI-aAlexandra MariaÎncă nu există evaluări
- Glandele EndocrineDocument1 paginăGlandele EndocrineAna Maria ToporascuÎncă nu există evaluări
- PLD Clasa 9 BiologieDocument7 paginiPLD Clasa 9 BiologieAliona Ali100% (1)
- Perturbari Ale MetabolismuluiDocument2 paginiPerturbari Ale MetabolismuluiLoretta IftinchiÎncă nu există evaluări
- Recapitulare AnalizatoriDocument6 paginiRecapitulare AnalizatoriiuliamaghiarÎncă nu există evaluări
- Inervatia Simpatica A Inimii. Baiasu Maria Florina, gr2Document5 paginiInervatia Simpatica A Inimii. Baiasu Maria Florina, gr2mihai mayorÎncă nu există evaluări
- Sistemul Circulator La VertebrateDocument1 paginăSistemul Circulator La VertebrateVic AlÎncă nu există evaluări
- Hidroliza AtpDocument4 paginiHidroliza AtpileanaÎncă nu există evaluări
- MiozinaDocument22 paginiMiozinaOlea Croitor0% (1)
- Biologie Xi CorigentaDocument3 paginiBiologie Xi CorigentaBlajiu BeatriceÎncă nu există evaluări
- Rezumat Curs BotanicaDocument34 paginiRezumat Curs BotanicaispasÎncă nu există evaluări
- Enzime Din Plasma SangvinaDocument8 paginiEnzime Din Plasma SangvinacharoiteÎncă nu există evaluări
- STERIDELEDocument3 paginiSTERIDELEcicku91Încă nu există evaluări
- Digestia Gastrica Intestinala Si AbsorbtiaDocument16 paginiDigestia Gastrica Intestinala Si AbsorbtiaDee Bua100% (1)
- Glandele EndocrineDocument6 paginiGlandele EndocrineAlexandra SimionovÎncă nu există evaluări
- BiomoleculeDocument8 paginiBiomoleculeElena PetreÎncă nu există evaluări
- Fisa de Lucru LacDocument3 paginiFisa de Lucru LacDaniela AvramÎncă nu există evaluări
- LP 1 AdnDocument18 paginiLP 1 AdnGabriela Victoria MartinescuÎncă nu există evaluări
- ProteineDocument5 paginiProteinedandreescu_1Încă nu există evaluări
- CerebelulDocument11 paginiCerebelulCorina Stângă-GrasuÎncă nu există evaluări
- Glandele SuprarenaleDocument2 paginiGlandele SuprarenaleAna Maria ToporascuÎncă nu există evaluări
- Sinteze Și Aplicații Bac - Clasa A Xii-ADocument10 paginiSinteze Și Aplicații Bac - Clasa A Xii-ATeodora Coroi100% (1)
- Relatiile Dintre Structura Si Functiile ADNDocument1 paginăRelatiile Dintre Structura Si Functiile ADNRamona DanyÎncă nu există evaluări
- Digestia Și AbsorbțiaDocument5 paginiDigestia Și AbsorbțiaGigel FroneÎncă nu există evaluări
- Caracteristicile Biochimice Fundamentale Ale Materiei ViiDocument8 paginiCaracteristicile Biochimice Fundamentale Ale Materiei ViidenisemichelaÎncă nu există evaluări
- UroteliulDocument1 paginăUroteliulatd999Încă nu există evaluări
- Emisfere CerebraleDocument13 paginiEmisfere CerebraleMargittai MihaelaÎncă nu există evaluări
- Celula EucariotaDocument15 paginiCelula EucariotaveronicaÎncă nu există evaluări
- Rolul Atp Si AdpDocument18 paginiRolul Atp Si AdpZota Georgiana Andreea100% (2)
- Biologie Clasa A XI A Sem I Toata MateriaDocument27 paginiBiologie Clasa A XI A Sem I Toata MateriaMadalinaBriscoiuÎncă nu există evaluări
- Cursul 2 Metabolismul + Metabolismul ApeiDocument5 paginiCursul 2 Metabolismul + Metabolismul ApeiBalint Ovidiu100% (2)
- MeiozaDocument2 paginiMeiozaEmilia SabauÎncă nu există evaluări
- Compozitia Chimica Si Organizarea Materiei ViiDocument19 paginiCompozitia Chimica Si Organizarea Materiei ViivirtualmorenoÎncă nu există evaluări
- Reglajul Genetic La EucarioteDocument10 paginiReglajul Genetic La Eucariotepantea_elena100% (1)
- ReferatDocument16 paginiReferatMarius VelescuÎncă nu există evaluări
- Procese Corticale. Coscodan Diana. Ora PublicaDocument20 paginiProcese Corticale. Coscodan Diana. Ora PublicaLena Bondari-CebotariÎncă nu există evaluări
- Structura ProteinelorDocument13 paginiStructura ProteinelorA B100% (1)
- BIOCEL Sinteza-1-1Document45 paginiBIOCEL Sinteza-1-1Adelin FrumusheluÎncă nu există evaluări
- (PDF) Referat Lipide - Compress PDFDocument3 pagini(PDF) Referat Lipide - Compress PDFSergiu BeldimanÎncă nu există evaluări
- PlasmalemaDocument1 paginăPlasmalemaAndruşcă LoredanaÎncă nu există evaluări
- Sinteza ProteinelorDocument17 paginiSinteza ProteinelorPavel MexyÎncă nu există evaluări
- Curs 9 - Traducerea Informatiei GeneticeDocument21 paginiCurs 9 - Traducerea Informatiei GeneticeionutÎncă nu există evaluări
- Subiectele 66-80Document16 paginiSubiectele 66-80pelinÎncă nu există evaluări
- Curs 1 Biologie Celulara Si Molecular ADocument19 paginiCurs 1 Biologie Celulara Si Molecular AOana Koppandi100% (1)
- Funcții-Replicația 1Document6 paginiFuncții-Replicația 1Așerita Benea100% (1)
- ArnDocument116 paginiArnGrosu Marius100% (1)
- BacteriiDocument2 paginiBacteriichris_ty_an7256Încă nu există evaluări
- De Cumparat Pentru CopiiDocument1 paginăDe Cumparat Pentru Copiichris_ty_an7256Încă nu există evaluări
- FiziopatDocument7 paginiFiziopatchris_ty_anÎncă nu există evaluări
- Echilibru LeucocitarDocument24 paginiEchilibru Leucocitarchris_ty_an7256Încă nu există evaluări
- TABEL MODEL Nevoi FundamentaleDocument4 paginiTABEL MODEL Nevoi Fundamentalechris_ty_an7256Încă nu există evaluări
- AP. DigestivDocument7 paginiAP. Digestivchris_ty_an7256Încă nu există evaluări
- Dinamica Relatiei Asist-PacientDocument5 paginiDinamica Relatiei Asist-Pacientchris_ty_an7256Încă nu există evaluări
- Calitatea Vietii Si Sanatatea OamenilorDocument15 paginiCalitatea Vietii Si Sanatatea OamenilorDan ChirtoacaÎncă nu există evaluări
- BibliografieDocument11 paginiBibliografiechris_ty_an7256Încă nu există evaluări
- Hemoleucograma CompletaDocument3 paginiHemoleucograma CompletapauastefaniaÎncă nu există evaluări
- Echilibru LeucocitarDocument24 paginiEchilibru Leucocitarchris_ty_an7256Încă nu există evaluări
- A.M.F. III Subiecte Proba Practica 2015Document28 paginiA.M.F. III Subiecte Proba Practica 2015AdelinaGhiorţu100% (1)
- ExamenDocument3 paginiExamenchris_ty_an7256Încă nu există evaluări
- BiohDocument42 paginiBiohchris_ty_an7256Încă nu există evaluări
- Instructiuni de Lucru - Hormoni Si Markeri Tumorali - MaiDocument2 paginiInstructiuni de Lucru - Hormoni Si Markeri Tumorali - Maichris_ty_an7256Încă nu există evaluări
- AneDocument19 paginiAnechris_ty_an7256Încă nu există evaluări
- MetabolismulDocument11 paginiMetabolismulchris_ty_an7256100% (1)
- Southern BlottingDocument10 paginiSouthern Blottingchris_ty_an7256Încă nu există evaluări
- As Lab Bacte1Document8 paginiAs Lab Bacte1chris_ty_an7256Încă nu există evaluări
- As Lab Bacte2Document11 paginiAs Lab Bacte2chris_ty_an7256Încă nu există evaluări
- Antiseptice AnorganicaDocument9 paginiAntiseptice Anorganicachris_ty_an7256Încă nu există evaluări
- As Lab ViroDocument7 paginiAs Lab Virochris_ty_an7256Încă nu există evaluări
- CARACTERIZAREDocument3 paginiCARACTERIZAREGeorgiana Ulea75% (4)
- Chestionar Nivel 2 de BiosigurantaDocument3 paginiChestionar Nivel 2 de Biosigurantachris_ty_an7256Încă nu există evaluări
- Dieta RinaDocument31 paginiDieta Rinachris_ty_an7256Încă nu există evaluări
- New Document Microsoft WordDocument25 paginiNew Document Microsoft Wordchris_ty_an7256Încă nu există evaluări
- PGL - 5.4.4 - Recoltarea Şi Manipularea Probei PrimareDocument111 paginiPGL - 5.4.4 - Recoltarea Şi Manipularea Probei Primarechris_ty_an7256Încă nu există evaluări
- 1 Proiect Didactic Prop CH MonozaharideDocument4 pagini1 Proiect Didactic Prop CH Monozaharidechris_ty_an7256Încă nu există evaluări
- Principii de Baza Ale CercetariiDocument34 paginiPrincipii de Baza Ale Cercetariidanutz18z67% (3)