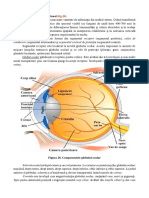
S-ar putea să vă placă și
- Analizatorul VizualDocument11 paginiAnalizatorul VizualMaria100% (1)
- NEUROPSIHOLOGIE: elementele de bază ale problemeiDe la EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiEvaluare: 5 din 5 stele5/5 (1)
- BIOLOGIE Suport de CursDocument34 paginiBIOLOGIE Suport de CursIrina Florentina100% (1)
- REFERAT Analizator VizualDocument5 paginiREFERAT Analizator VizualFlorentina JianuÎncă nu există evaluări
- Sistemul CardiovascularDocument11 paginiSistemul CardiovascularAndreea Stefania100% (1)
- Organele de Simt STRUCTURADocument6 paginiOrganele de Simt STRUCTURAMaria NicuÎncă nu există evaluări
- Analizatorii Sunt Sisteme Morfofunctionale Prin Intermediul CaroraDocument11 paginiAnalizatorii Sunt Sisteme Morfofunctionale Prin Intermediul Caroraisidor_mihaelaÎncă nu există evaluări
- Analizatorul VizualDocument7 paginiAnalizatorul VizualLili BujorÎncă nu există evaluări
- Analizatori PlatformaDocument37 paginiAnalizatori PlatformaMotcoÎncă nu există evaluări
- Analizatorul VizualDocument11 paginiAnalizatorul VizualMiruna Deleanu MariaÎncă nu există evaluări
- Organele de Simț - BiologieDocument1 paginăOrganele de Simț - BiologieBRIANNA LUCAÎncă nu există evaluări
- Proiect Absolvire An 2Document66 paginiProiect Absolvire An 2Mariana Daminescu100% (3)
- OCHIULDocument7 paginiOCHIULIonica Madalina100% (1)
- Organele de Simt.Document7 paginiOrganele de Simt.Obada AnaÎncă nu există evaluări
- Misca Bristena-Igiena Analizatorului VizualDocument21 paginiMisca Bristena-Igiena Analizatorului VizualInu PriescuÎncă nu există evaluări
- C13 Analizatori Analizatorul VizualDocument7 paginiC13 Analizatori Analizatorul VizualDelia CristeaÎncă nu există evaluări
- Analizatorul VizualDocument12 paginiAnalizatorul VizualMîndru Lara100% (1)
- Neurofiziologia Analizatorilor SenzorialiDocument27 paginiNeurofiziologia Analizatorilor SenzorialiSiMonNykÎncă nu există evaluări
- AnalizatoriiDocument54 paginiAnalizatoriiGrecu MădălinaÎncă nu există evaluări
- Analizatorul Vizual OCHIULDocument9 paginiAnalizatorul Vizual OCHIULAna Maria ToporascuÎncă nu există evaluări
- Biosisteme 1Document8 paginiBiosisteme 1Cornelia ZahariaÎncă nu există evaluări
- Analizatorul VizualDocument9 paginiAnalizatorul VizualAlessandra McaÎncă nu există evaluări
- ANALIZATORIIDocument3 paginiANALIZATORIIAna IonescuÎncă nu există evaluări
- Analizatorul VizualDocument2 paginiAnalizatorul VizualJepan MariusÎncă nu există evaluări
- Suport Curs Biologie Clasa A 11-ADocument141 paginiSuport Curs Biologie Clasa A 11-Asonny me100% (1)
- Grile Titu Maiorescu Capitolul III. AnalizatoriiDocument6 paginiGrile Titu Maiorescu Capitolul III. AnalizatoriiManuela ConcioiuÎncă nu există evaluări
- AnalizatoriiDocument11 paginiAnalizatoriiCristina GheraÎncă nu există evaluări
- Analizatorul VizualDocument8 paginiAnalizatorul VizualDankmeisterÎncă nu există evaluări
- Analizatorii Si Glandele EndocrineDocument12 paginiAnalizatorii Si Glandele EndocrineMonica Amalia Fasie Halbac100% (1)
- ANALIZATORIIDocument24 paginiANALIZATORIIcalin.mihaelaÎncă nu există evaluări
- Structura Globului OcularDocument3 paginiStructura Globului OcularNicoleta TuleaÎncă nu există evaluări
- ANALIZATORIIDocument10 paginiANALIZATORIIBosneaga DanielaÎncă nu există evaluări
- Anatomie AMF IDocument149 paginiAnatomie AMF IadelacosereanuÎncă nu există evaluări
- Analizatorul VizualDocument41 paginiAnalizatorul Vizualmariana77100% (2)
- Functiile Organismului UmanDocument35 paginiFunctiile Organismului UmanClaudia Jicmon100% (1)
- BIOLOGIE Suport de Curs Clasa A XI-AFRDocument33 paginiBIOLOGIE Suport de Curs Clasa A XI-AFRMarysaLarysa100% (1)
- Cap. 12 Organele de SimțDocument7 paginiCap. 12 Organele de SimțDragos BirzuÎncă nu există evaluări
- Anatomia Analizatorului VizualDocument51 paginiAnatomia Analizatorului VizualFlorentina Mihaela IonescuÎncă nu există evaluări
- Analizatori BacDocument58 paginiAnalizatori BacCristina StamateÎncă nu există evaluări
- Functiile Organismului UmanDocument34 paginiFunctiile Organismului UmanSebastian GhiurÎncă nu există evaluări
- 2 AnalizatoriiDocument11 pagini2 AnalizatoriiOana GhitleÎncă nu există evaluări
- Ochii - Si - Defectele VederiiDocument45 paginiOchii - Si - Defectele VederiiMiha MihaelaÎncă nu există evaluări
- Analizator VizualDocument6 paginiAnalizator VizualCristiana MorariuÎncă nu există evaluări
- L9. Analizatorul VizualDocument4 paginiL9. Analizatorul VizualLaura FloreaÎncă nu există evaluări
- C7 Analizatorii Ochiul UrecheaDocument8 paginiC7 Analizatorii Ochiul UrecheaAlina FlorentinaÎncă nu există evaluări
- Organele de SimțDocument31 paginiOrganele de SimțPaul vlad 06Încă nu există evaluări
- Proiect OptometrieDocument24 paginiProiect OptometrieIonela-Georgiana Urse100% (7)
- FISA DE LUCRU Anatomie RezolvataDocument5 paginiFISA DE LUCRU Anatomie RezolvataRazvan VizitiuÎncă nu există evaluări
- Analizatorul Vizual A XIDocument10 paginiAnalizatorul Vizual A XITaniaPopescuÎncă nu există evaluări
- Oftalmologie Curs1-Anatomia AP. VizualDocument12 paginiOftalmologie Curs1-Anatomia AP. VizualsymohsnÎncă nu există evaluări
- Eseu AnalizatoriDocument3 paginiEseu AnalizatoriDigei BobitzÎncă nu există evaluări
- AnalizatoriDocument11 paginiAnalizatoriElena ElenaÎncă nu există evaluări
- Analizatorul VizualDocument16 paginiAnalizatorul VizualAny MinoiuÎncă nu există evaluări
- Temă-Analizator VizualDocument3 paginiTemă-Analizator VizualHăpăianu ElisaÎncă nu există evaluări
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- Maximizarea capacității intelectuale: Învățați cum să profitați la maximum de inteligența dumneavoastrăDe la EverandMaximizarea capacității intelectuale: Învățați cum să profitați la maximum de inteligența dumneavoastrăEvaluare: 5 din 5 stele5/5 (1)
- Metode FiziceDocument6 paginiMetode FiziceAndreea StefaniaÎncă nu există evaluări
- DIGESTIADocument4 paginiDIGESTIAAndreea StefaniaÎncă nu există evaluări
- Substante OrganiceDocument9 paginiSubstante OrganiceAndreea StefaniaÎncă nu există evaluări
- METABOLISMULDocument17 paginiMETABOLISMULAndreea StefaniaÎncă nu există evaluări
- Anatomie Curs 5 NouDocument9 paginiAnatomie Curs 5 NouAndreea StefaniaÎncă nu există evaluări
- Anatomie Curs 5 NouDocument9 paginiAnatomie Curs 5 NouAndreea StefaniaÎncă nu există evaluări
- Sistemul NervosDocument10 paginiSistemul NervosAndreea StefaniaÎncă nu există evaluări
- Anatomie Curs 2Document8 paginiAnatomie Curs 2Andreea StefaniaÎncă nu există evaluări
- TESUTURILEDocument4 paginiTESUTURILEAndreea StefaniaÎncă nu există evaluări
- Analizatorul AcusticovestibularDocument5 paginiAnalizatorul AcusticovestibularAndreea StefaniaÎncă nu există evaluări
- ANALIZATORIDocument3 paginiANALIZATORIAndreea StefaniaÎncă nu există evaluări
- Nevoia de A Respira Si A Avea o Buna Circulatie Si Nevoia de A Bea Si A MancaDocument5 paginiNevoia de A Respira Si A Avea o Buna Circulatie Si Nevoia de A Bea Si A MancaAnca Maria GersakÎncă nu există evaluări
- Emisferele CerebraleDocument16 paginiEmisferele CerebraleAndreea StefaniaÎncă nu există evaluări
- Nevoia de A Respira Si A Avea o Buna Circulatie Si Nevoia de A Bea Si A MancaDocument5 paginiNevoia de A Respira Si A Avea o Buna Circulatie Si Nevoia de A Bea Si A MancaAnca Maria GersakÎncă nu există evaluări
- Dieta Echilibrata EseuDocument3 paginiDieta Echilibrata EseuAndreea StefaniaÎncă nu există evaluări
- DInamica Peisajului Comuna Corbeni, Jud - ArgesDocument2 paginiDInamica Peisajului Comuna Corbeni, Jud - ArgesAndreea StefaniaÎncă nu există evaluări
- Nevoile FundamentaleDocument8 paginiNevoile FundamentaleAndreea StefaniaÎncă nu există evaluări
- Dictionar de TermeniDocument20 paginiDictionar de TermeniAndreea StefaniaÎncă nu există evaluări
- Nevoile FundamentaleDocument8 paginiNevoile FundamentaleAndreea StefaniaÎncă nu există evaluări
- Anatomie Curs 2Document8 paginiAnatomie Curs 2Andreea StefaniaÎncă nu există evaluări
- Studiul ChineiDocument12 paginiStudiul ChineiAndreea StefaniaÎncă nu există evaluări
- Fisa Localitatii 2016Document8 paginiFisa Localitatii 2016Andreea StefaniaÎncă nu există evaluări
- Dictionar de TermeniDocument20 paginiDictionar de TermeniAndreea StefaniaÎncă nu există evaluări
- Studiul ChineiDocument12 paginiStudiul ChineiAndreea StefaniaÎncă nu există evaluări
- Nevoile FundamentaleDocument8 paginiNevoile FundamentaleAndreea StefaniaÎncă nu există evaluări
- Dieta Echilibrata EseuDocument3 paginiDieta Echilibrata EseuAndreea StefaniaÎncă nu există evaluări
- Studiul ChineiDocument12 paginiStudiul ChineiAndreea StefaniaÎncă nu există evaluări
- Dieta Echilibrata EseuDocument3 paginiDieta Echilibrata EseuAndreea StefaniaÎncă nu există evaluări