Documente Academic
Documente Profesional
Documente Cultură
Curs Fizio Complet 1
Încărcat de
bucureanionut0 evaluări0% au considerat acest document util (0 voturi)
121 vizualizări54 paginiCurs Fiziologie Anul 1 sem 2
Oradea
Drepturi de autor
© © All Rights Reserved
Formate disponibile
PDF, TXT sau citiți online pe Scribd
Partajați acest document
Partajați sau inserați document
Vi se pare util acest document?
Este necorespunzător acest conținut?
Raportați acest documentCurs Fiziologie Anul 1 sem 2
Oradea
Drepturi de autor:
© All Rights Reserved
Formate disponibile
Descărcați ca PDF, TXT sau citiți online pe Scribd
0 evaluări0% au considerat acest document util (0 voturi)
121 vizualizări54 paginiCurs Fizio Complet 1
Încărcat de
bucureanionutCurs Fiziologie Anul 1 sem 2
Oradea
Drepturi de autor:
© All Rights Reserved
Formate disponibile
Descărcați ca PDF, TXT sau citiți online pe Scribd
Sunteți pe pagina 1din 54
1
Denevrarea muschiului scheletic
Rezulta in urma distructiei motoneuronului Alfa medular provocata de virusul polio,
zdrobirii terminatiei nervoase. Fibra nervoasa provenita din corionul alterat, genereaza
impulsuri care determina contractia unor grupe musculare. Contractia se poate observa cu
ochiul liber sub tegument si poarta denumirea de FASCICULATIE.
Activitatea electrica a muschiului care se contracta se poate inregistra de la suprafata
tegumentului.
Dupa 3-5 zile fibrele musculare insasi prezinta o sensibilitate crescuta fata de
mediatori. Ele produc spontan impulsuri, la inceput 1/secunda, apoi 3-5/secunda,
determinand contractii fine ale fibrelor musculare = fibrilatie musculara. Activitatea electrica
a fibrei musculare poate fi evidentiata prin introducerea unor electrozi sub forma de ac in
muschi (nu se observa la suprafata).
MIASTENIA:
- oboseala permanenta, mai exprimata la extremitatea cefalica, membrelor
superioare.
- imposibilitatea efectuarii unor miscari sau chiar
-paralizia
Cauza este alterarea transmiterii impulsurilor in jonctiunea neuro-musculara. Membrana
postsinaptica are faldurile reduse ca diametru in acest caz. Spatiul sinaptic creste la 20-30 -
50 n . Cauza reducerii dimensiunilor faldurilor este autoimuna. Organismul unor persoane
produce anticorpi fata de receptorii colinergici de pe membrana postsinaptica.
O parte din R sunt distrusi si altii formeaza complexe cu anticorpii si sunt captati prin
ENDOCITOZA. Proportia receptorilor colinergici din membrana se reduce la 20%. In conditiile
eliberarii unor cantitati suficiente de ACO eu nu sunt capabili sa declanseze un potential
postsinaptic care sa se propage.
La 10-20% din bolnavi exista o tumora a timusului. Extirparea tumorii imbunatateste
starea bolnavului. Si la persoanele care nu prezinta tumora de timus s-a observat o
ameliorare a miasteniei prin extirparea timusului. Timusul are o antigenicitate similara cu
cea a membranei postsinaptice.
Frecventa MIASTENIEI = 1:20 000 persoane
Tratament cu fizostigmina si neostigmina -> mareste concentratia ACO in fanta sinaptica,
favorizand contractia.
2
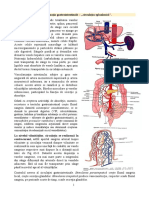
DIGESTIV
Histamina reprezint una dintre substanele endogene cu o puternic aciune excitant
asupra celulelor parietale (oxin-tice). Sinteza unor mari cantiti de histamina are loc n
celulele asemntoare cu cele enterocromafine din mucoasa gastric secretoare de acid, ce
sunt situate n imediata apropiere a celulelor parietale. Celulele productoare conin histidin
car-boxilaza, enzim care particip la formarea histaminei.
Pe suprafaa celulelor productoare de histamina se afl receptori pentru gastrin i pentru
acetilcolin. Gastrina acioneaz ca stimulator al secreiei gastrice, predominant pe celulele
care sintetizeaz histamina. Histamina acioneaz asupra a dou tipuri de receptori
membranari notai cu Hi i H
2
. Fixarea histaminei pe receptorii H
2
determin intensificarea
secreiei gastrice acide. Antihistaminicele folosite n tratamentul strilor alergice blocheaz
receptorii H], ns nu scot din funciune receptorii H
2
, nct aciunea acidosecretant a
histaminei continu. Blocarea receptorilor H
2
cu cimetidin, ranetidin, nizatidin i
famotidin inhib la om efectul stimulator al histaminei, gastrinei i alimentelor. S-a ajuns
astfel la concluzia c histamina constituie un mediator al stimu Iilor acido-secretani din
mucoasa gastric.
Efectul excitosecretor gastric al histaminei exogene apare chiar i dup administrarea s.c. a
unor doze mici de substan (0,2-0,3 mg). Aceeai doz injectat i.v are influen redus
asupra secreiei gastrice.
n mod fiziologic secreia gastric acid poate s fie sporit de ageni coli-nergici
(acetilcolin, carbacol), histamina i gastrin. Pe membrana laterobazal a celulelor parietale
(oxintice) s-au identificat receptori pentru cele trei tipuri de substane. Receptorii pentru
gastrin sunt prezeni pe celulele parietale ale cinilor, obolanilor i altor specii, ns lipsesc
de pe celulele acidosecretoare umane i de iepure. Efectul puternic stimulator al gastrinei
asupra secreiei gastrice acide la om este mediat de histamina.
Intensificarea formrii acidului clorhidric, dup activarea receptorilor, are loc prin
intermediul unor mesageri de ordinul II. Interaciunea acetilcolinei cu receptorii muscarinici
(M
3
) i a gastrinei, cu receptorii specifici (la animalele la care exist), determin creterea
concentraiei ionilor de Ca
++
n citoplasm celulelor parietale. Ionii de Ca** sunt mobilizai
din depozitele intracelulare de inozitol trifosfat (IP
3
) sau provin din lichidul extracelular, n
urma deschiderii canalelor de calciu din membrana celular. Mrirea concentraiei Ca"
intracelu-lar, determin migrarea formaiunilor tubuloveziculare i fuzionarea lor n
canalicule.
Dup fixarea histaminei pe receptori, prin intermediul unei proteine G stimulatoare, are loc
activarea adenilciclazei cu formarea AMPc intracelular. Procesele de fosforilare iniiate de
AMPc, cresc permeabilitatea canalelor ionice din membrana celulelor parietale i nivelul Ca
++
liber intracelular. AMPc mai intervine n activarea cotransportului de KC1. Fr prezena K
+
pe suprafaa luminal, K
+
-H
+
ATP-aza nu funcioneaz (Fig. 24).
Alturi de receptorii pentru histamina, pe membrana laterobazal a celulelor parietale se
afl receptori pentru prostaglandina E2, care prin intermediul unei proteine G inhibitoare
mpiedic formarea AMPc.
3
Secreia de pepsinogen din celulele principale (zimogene) este intensificat de substanele
colinergice i agonitii beta adrenergici. Concentraia HC1 din stomac de asemenea
influeneaz producia de pepsinogen. La persoanele cu aclorhidrie se constat un nivel
extrem de sczut al pepsinei n sucul gastric, dei celulele parietale au un aspect normal.
Fig 24. Reglarea secreiei acide gastrice din celulele parietale.
Fazele secreiei gastrice
Controlul secreiei gastrice a fost divizat n trei faze n funcie de regiunea asupra creia
stimulii secretori i exercit aciunea: faza cefalic, gastric i intestinal.
Faza cefalic
Pe animale cu fistul esofagian i pung gastric inervat, Pavlov a evideniat mecanismul
nervos reflex de producere a fazei cefalice. La asemenea animale, dei alimentele nu ajung n
stomac (prnz fictiv), n perioada masticaiei i deglutiiei se nregistreaz o cretere a se-
creiei gastrice, ce dispare ns dupa secionarea nervilor vagi. Contactul alimentelor cu
mucoasa bucal iniiaz reflexul, a crui cale eferent este format din nervii vagi.
Stimularea secreiei se poate produce, pe lng reflexele necondiionate i prin reflexele
condiionate. Secreia din faza cefalic lipsete cnd alimentele sunt in-gerate fr poft.
n faza cefalic, sub influena impulsurilor vagale se elibereaz att acetilco-lin ct i
gastrin, responsabile pentru secreia a 30-50% a acidului clorhidric,ca rspuns la ingerarea
normal de alimente.
De aceea sucul gastric elaborat n faza cefalic, conine mari cantiti de acid clorhidric i
pepsin. Acelai tip de suc se
obine i dup excitarea nervului vag.
Stimularea hipotalamusului i a poriunilor adiacente scoarei frontale, intensific
activitatea eferent vagal i totodat secreia gastric. Hipoglicemia insulinic, are un efect
similar, mediat probabil prin aciune direct asupra hipotalamusului. Starea de saietate
influeneaz n mare msur secreia gastric. Distrugerea nucleilor ventrolaterali din
hipotalamus (centrii foamei), pe lng producerea unei afagii de durat, duce i la pierderea
capacitii insulinei de a stimula secreia gastric. Excitarea hipotalamusului posterior
determin de asemenea o cretere tardiv a secreiei gastrice printr-un mecanism hormonal
4
hipofizo-suprarenal. Creterea concentraiei glicocorticoizilor circulani la cine, aprut n
urma administrrii de ACTH, mrete secreia gastric de acid i pepsin. Glicocorticoizii
scad cantitatea i schimb compoziia mucusului gastric, de aceea administrarea lor prelungit
predispune la ulcere. Bolnavii cu atrofia glandelor suprarenale (boala lui Addison) prezint de
obicei aclorhidrie, ceea ce pledeaz pentru intervenia hormonilor suprarenali n ntreinerea
secreiei gastrice. Agonitii beta adrenergici, somatostatina, factorul de cretere epidermal
inhib activitatea secretorie a stomacului. Reaciile de furie mresc secreia gastric, iar cele
de team, de nelinite i depresia o inhib.
Faza gastric
Faza gastric a secreiei ncepe odat cu ptrunderea alimentelor n stomac i se continu 3-
4 ore, timp n care alimentele rmn n cavitatea gastric. Introducerea unor alimente direct n
stomac printr-o fistul, declaneaz faza gastric de secreie. Sucul elaborat n faza gastric
are de asemenea un puternic caracter acid i un coninut ridicat n pepsin. Secreia din faza
gastric este mediat reflex, mediatorii fiind acetilcolina i ndeosebi gastrina. Receptorii
reflexului se afl situai n peretele gastric i sunt stimulai de destinderea stomacului sau de
substane chimice, mai ales de aminoacizii rezultai din digestie. De la receptori fibrele
ptrund n plexul submucos al lui Meiss-ner unde sunt localizai pericarionii. Impulsurile
ajung apoi la neuronii postgan-glionari parasimpatici, care se termin pe celulele parietale i
mresc secreia de HC1. Reflexul secretor local descris are toate elementele arcului reflex n
pereii stomacali.
Lansarea gastrinei n circulaie se face att sub influena impulsurilor vagale (nc din faza
cefalic), ct i prin stimularea direct a mucoasei antrale. Stimulii locali pot fi reprezentai de
modificarea pH-ului, de unele substane chimice numite secretagoge ca: etanolul, cafeina,
aminoacizii (glicina, serina, alanina, Uzina),extractele de carne, ionii de Ca" sau de distensia
mecanic a antrului. Simpla cretere a pH-ului regiunii antrale de la 3,5 la 7 sau 8 determin o
mrire semnificativ a concentraiei gastrinei n sngele venos, iar scderea pH-ului o reduce.
Pn la 70% din secreia stomacului se realizeaz n faza gastric.
Gastrina are efecte trofice asupra mucoasei gastrice i duodenale, ceea ce ar putea explica
n parte apariia atrofiei mucoasei gastrice la om, dup rezecie. Aciunea trofic se datorete
activrii de ctre hormon a mitozelor i sintezei de ADN. Gastrina determin i contracia
sfincterului gastroesofagian, mrete secreia de insulina, de glucagon i calcitonin.
Faza intestinal
Existena fazei de secreie intestinal a fost dovedit prin introducerea alimentelor direct n
duoden printr-o fistul, sau prin distensia intestinului subire, manopere ce sunt urmate de
creterea secreiei gastrice. Stimularea secreiei gastrice se face mai ales prin elaborarea local
a gastrinei sau a presupusului hormon - enterooxintin.
Motilina (polipeptid format din 22 aminoacizi), eliberat de mucoasa intestinal ori de cte
ori pH-ul duodenal atinge valoarea de 8,2 stimuleaz secreia i motricitatea stomacului.
Volumul secreiei din faza intestinal este foarte redus i srac n acid, el reprezentnd abia
10% din secreia gastric postalimentar.
Inhibiia secreiei gastrice
5
Coborrea pH-ului sucului gastric sub 3 duce la inhibarea secreiei gastrice, constituind un
important mecanism au-toreglator. Inhibiia provocat de acid nu se produce pe cale nervoas,
ntruct anestezicele aplicate pe mucoasa antral nu o pot nltura. Ionii de H
+
mresc secreia
de somatostatin i acioneaz inhibitor direct asupra celulelor eliberatoare de gastrin.
Grsimile, acidul clorhidric, soluiile hipertonice n contact cu mucoasa duodenal,
elibereaz unul sau mai muli factori umorali cu efecte inhibitoare asupra secreiei gastrice ce
au fost puse pe seama unui presupus hormon numit enteroga-stron (Osaka i Lim, 1930).
Actualmente exist mai multe polipeptide ce candideaz la titlul de enterogastron. Printre ele
se ncadreaz peptidul YY. secretina, colecistokinina, peptidul gastric inhibitor, peptidul
vasoactiv intestinal, somatostatin si prostaglandinele.
Polipeptidul gastric inhibitor (Gastric inhibitory polypeptide - GIP) a fost extras din
mucoasa jejunal de porc. El (este alctuit din 43 aminoacizi) inhib att secreia de HC1 ct
i motilitatea gastric. La 45 minute dup ingerarea de grsimi sau glucide, GIP crete n
snge de cinci ori. Inhibarea secreiei gastrice produs de GIP are drept mediator
somatostatin.
Din mucoasa intestinului subire i chiar a colonului s-a izolat polipeptidul vasoactiv
intestinal (Vasoactive intestinal polypeptide - VIP)(ce conine 28 de aminoacizi). Hormonul
pe lng efectele inhibitoare asupra secreiei acide a stomacului, produce dilataia vaselor
periferice i stimuleaz puternic secreia intestinal de electrolii i ap.
Att GIP ct i VIP au o structur ce se aseamn cu secretina i cu glucagonul.
Hormonul somatostatin, adic inhibitorul STH, evideniat pentru prima dat n
hipotalamus, este un peptid n structura cruia intr 14 sau 28 de aminoacizi (ambele forme
fiind active), ce mpiedic secreia acid gastric, activitatea motorie a stomacului i
elaborarea sucului pancreatic.
n mucoasa gastric a diferitelor specii animale (obolani, cobai, oi) i la om s-au
identificat prostaglandine de tip E i F (PGEi, PGE
2
, PGFi alfa, PGF
2
alfa). Grupa E de
prostaglandine, are efecte inhibitoare asupra secreiei gastrice stimulate de gastrin sau
histamin. Diminuarea secreiei gastrice sub influena PGEi, se explic (aa cum s-a
menionat anterior), prin scderea AMPc intracelular. Aspirina are efecte ulcerigene, n mare
msur prin mpiedicarea sintezei de prostaglandine n mucoasa gastric.
Metode de explorare a secreiei gastrice la om
Recoltarea sucului gastric la om se face prin sondaj gastric. Pentru stabilirea dinamicii
Reflexul de vom
Voma reprezint procesul de evacuare rapid a coninutului gastric prin esofag i cavitatea
bocal. nainte de declanarea vomei, la oameni apare o senzaie intens de grea, asociat cu
o reacie simpatic, exteriorizat prin midriaz (dilatarea pupilelor), paloare, transpiraie,
accelerarea ritmului cardiac i respirator i o reacie parasimpatic cu hipersalivaie,
lcrimare, contracii antiperistaltice duodenale, gastrice i esofagiene, precum i relaxarea
sfincterelor esofagian inferior i superior.
Reflexul de vom ncepe cu cteva inspiraii profunde dup care respiraia se oprete,
laringele se ridic determinnd nchiderea glotei i deschiderea sfincteru-lui esofagian
6
superior. Nasofaringele este blocat prin ridicarea palatului moale. n acelai timp se produce o
relaxare a corpului, fundului stomacului, esofagului i contracia antrului piloric, alturi de
contracia puternic a diafragmului i musculaturii abdominale. Evacuarea coninutului
stomacal n cursul vomei se datorete mai ales presiunii exercitate asupra lui de la exterior i
mai puin prin contraciile proprii. Dup administrarea de curara, care paralizeaz musculatura
striat, voma nu se mai produce. De altfel, F.Magendie (1836) a demonstrat c voma are loc
i dup nlocuirea stomacului cu o vezic urinar de porc.
Voma este un act reflex declanat de stimularea receptorilor rspndii n diferite organe.
Reflexul poate fi iniiat de impulsuri pornite din stomac, viscerele abdominale (apendice,
colecist, intestin, pancreas, ci renale, uter) i din torace (inim, n cazul infarctului
miocardic). Cauza vomei de origine digestiv poate fi iritaia mucoasei, excitabilitatea mrit
a mucoasei, distensia anormal a tractului gastro-intestinal ori a cilor biliare. Mirosurile
dezagreabile, vederea unor alimente neplcute sau stimularea receptorilor vestibulari, n
timpul deplasrii cu anumite vehicule, determin apariia vomei prin mecanisme corticale sau
reflexe.
Fibrele aferente ale arcului reflex cu punct de plecare gastric sau abdominal sunt ataate
nervilor vagi i simpatici.
Centrii reflexului de vom sunt situai n regiunea dorso-lateral a formaiei reticulare din
bulb (n vecintatea centrului respirator). n apropierea centrului vomei, a fost descris o zon
chemoreceptoare "trigger" sau zona trgaci (Borison i Wang, 1953), excitarea creia poate
declana voma.
Zona chemoreceptoare "trigger" este tangent sau chiar inclus n area postrema, o band
de esut cu form de V, situat n pereii laterali ai ventriculului IV, aproape de obex. Ea face
parte din organele circumventriculare i are o permeabilitate vascular mai mare fa de restul
creierului fiind lipsit de barier hematoencefalic. Substanele care produc voma se numesc
emetizante i acioneaz fie prin iritarea mucoasei gastrice, fie prin influenarea zonei
"trigger". Apomorfna, morfina, ipeca, picrotoxina, sulfatul de cupru i pierd efectele
vomitive dup distrugerea zonei "trigger". De aceea se consider c apomorfna i celelalte
substane amintite au numai aciune central i nu periferic. Substanele toxice endogene
cum sunt cele care se acumuleaz n snge n acidoza diabetic, insuficiena renal,
graviditate, sau anumite toxine exogene produc voma att prin mecanism periferic, de excitare
a receptorilor gastrici, ct i prin mecanism central, de stimulare a zonei "trigger".
n intestinul subire, serotonina eliberat din celulele enterocromafine, acionnd asupra
receptorilor 5-HT
3
iniiaz voma. Area postrema i nucleul tractului solitar conin att
receptori 5-HT
3
ct i receptori dopaminergici D
2
. Antagonitii receptorilor 5-HT
3
ca
ondansteronul i ai celor D
2
ca i clorpromazina i haloperi-dolul au efecte antiemetizante.
De la centrii, impulsurile sunt conduse eferent prin nervii frenici, vagi, simpatici, nervii
spinali ai musculaturii abdominale i nervii cranieni ai faringelui i palatului (Fig. 30).
7
Excitarea direct a centrilor vomei se produce prin creterea presiunii in-tracraniene.
Tumorile localizate n ventriculul IV, sau iritaiile meningeale determin vrsturi "n
proiectil", care nu sunt precedate de grea.
Explorarea motricitatii gastrice Motricitatea gastric la om se exploreaz prin examenul
radiologie al stomacului dup ingerarea unei substane de contrast.
Se utilizeaz i metoda viscerografic de nregistrare a micrilor stomacului, prin
intermediul unui balona introdus n stomac i destins, care transmite modificrile de presiune
unei capsule Marey sau cu ajutorul traductorilor de presiune.
ntre metodele recente de explorare a evacurii gastrice se ncadreaz metoda
radioizotopic, care const n adugarea unui izotop (
99
Tc) la un prnz mixt i nregistrarea
radioactivitii din zona epi-gastric.
Metoda ecografic permite urmrirea conturului stomacal dup ingerarea unui prnz mixt
i n timpul evacurii.
Viteza de golire a stomacului la trecerea prin pilor poate fi urmrit prin ecografie Doppler.
Ulcerul gastro-duodenal - Cea mai frecvent afeciune a stomacului este ulcerul. Boala
apare n urma compromiterii mecanismelor de protecie a mucoasei stomacale, sucul gastric
putnd astfel s duc la un proces de au-todigestie. In ulcerele duodenale i prepilorice,
secreia clorhidropeptic mrit contribuie la formarea leziunii.
La distrugerea barierei protectoare gastrice contribuie infecia cu Helicobacter pilori. n
plus, Helicobacter pilori poate elibera o toxin ce determin inflamaia i chiar ulcerarea
mucoasei.
Importana hipersecreiei de HCl i pepsin n geneza ulcerelor duodenale i a celor
prepilorice reiese i din ceea ce se ntmpl n sindromul Zollinger-Ellison, ntlnit la bolnavii
cu tumori secretoare de gastrin (gastrinoame). Tumorile sunt localizate mai frecvent n
pancreas, mai rar n stomac sau duoden. Excesul de gastrin provoac o hipersecreie
prelungit i ulceraii multiple, cu evoluie sever.
8
Refluxul bilei din duoden n stomac ar fi implicat n producerea ulcerului gastric. La
bolnavii cu ulcer gastric regurgitarea bilei n stomac este mai frecvent i mai voluminoas
dect la persoanele sntoase i s-ar datora unui rspuns contracii insuficient al pilorului la
aciunea colecistokininei i secretinei.
Tratamentul medicamentos al bolnavilor cu ulcer duodenal vizeaz reducerea secreiei
gastrice acide i consolidarea mijloacelor de protecie a mucoasei. Pentru neutralizarea
acidului clorhidric produs n exces, se utilizeaz medicamente antiacide, care conin hidroxid
de Al, carbonat de Ca, hidroxid de Mg. O reducere nsemnat a secreiei de HC1 se obine
prin blocarea receptorilor H
2
cu cimetidi-n, ranetidin, nizatidin, famotidin. Receptorii
muscarinici de tip M, pot fi blocai cu atropin sau cu pirenzepin. Omeprazolul poate inhiba
H
+
-K
+
ATP-aza pentru timp ndelungat. O sare bazic de Al a octasulfatului de sucroz
numit Sucralfat, mrete rezistena mucoasei la aciunea corosiv a acidului clorhidric,
formnd o protein aderent la suprafaa ulcerului. n ulcerele duodenale complicate se
recurge la interveniile chirurgicale, care constau n vagotomie, combinat cu rezecia regiunii
antrului pilone, a crei mucoas sintetizeaz gastrina.
DIGESTIA N INTESTINUL SUBIRE
Digestia alimentelor nceput n cavitatea bucal, continuat n stomac, este finisat n
intestinul subire. Enzimele intestinale au pH-ul optim de aciune neutru. Pe parcursul
intestinului aciditatea chimului se reduce n urma schimbului dintre ionii de H
+
i Na
+
prin
peretele intestinal i datorit aciunii neutralizante a sucului pancreatic, bilei i sucului intesti-
nal propriu zis.
In prima poriune a intestinului subire se vars sucul pancreatic i bila.
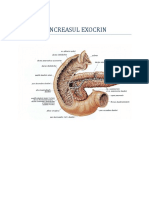
SCREIA PANCRETICA
Pancreasul exocrin este constituit din acini glandulari similari cu cei din glandele salivare.
Celulele glandulare conin granule de zimogen, pe care le descarc prin exocitoz n duetele
pancreatice. Intre celulele acinilor glandulari i lu-menul duetului pancreatic se interpun n
anumite poriuni, nite celule lite numite celule centroacinare, care aparin duetului.
Sucul pancreatic, produsul de secreie extern a pancreasului, se vars n duoden prin
canalul Wirsung i uneori prin canalul accesor al lui Santorini. Persoanele crora le lipsete
canalul accesor sunt mult mai predispuse la ulcer duodenal. Fenomenul se explic prin lipsa
secreiei alcaline pancreatice din partea superioar a duodenului (canalul Santorini se vars n
duoden cu 2 cm mai sus dect canalul Wirsung)(Fig. 31).
In funcie de stimulul aplicat, pancreasul secret dou tipuri de sucuri: 1) un suc apos,
abundent, foarte alcalin, srac n enzime; 2) un suc bogat n enzime, ns cu volum redus.
Secreia pancretica normala reprezint un amestec ntre cele dou tipuri de sucuri.
9
Compoziia sucului pancreatic. Pancreasul secret un suc clar, transparent, uor vscos cu
un volum de 1000-1500 ml/24 ore. Conine 98,5% ap i un reziduu de 1,5%.
a. Substanele anorganice care intr n alctuirea sucului pancreatic sunt: cationii de Na\
K
+
, Ca
++
, Mg++ n concentraii egale cu cele din plasm i dintre anioni, ionul de bicarbonat
ntr-o concentraie mai mare dect n plasm, apoi ionii de clor, fosfat, sulfat. Electroliii sunt
secretai att de celulele duetelor pancreatice ct i de celulele centroacinare. Cu ct debitul
secretor pancreatic este mai mare, cu att crete i concentraia ionului de bicarbonat i scade
concentraia ionului CI". De la 66 rnEq/1 (n plasm 24-25 mEq/1) ionul de bicarbonat poate
s ajung la 140-150 mEq/1. Suma ionilor de bicarbonat i cei de clor rmne constant, adic
pe msur ce sporete concentraia ionilor de HC0
3
-
, diminueaz concentraia ionilor de CI
-
.
Ionul de bicarbonat care mpreun cu Na
+
d natere bicarbonatului de Na, face ca pH-ul
sucului pancreatic s fie alcalin (7,5-8,0) (Fig. 32).
b. n formarea bicarbonatului joac rol anhidraza carbonic, localizat n celulele duetelor
pancreatice de mic calibru, care catalizeaz transformarea C0
2
(sanguin sau produs local) i a
apei n acid carbonic, ce disociaz spontan n ioni de bicarbonat i ioni de hidrogen. Blocanii
an-hidrazei carbonice (acetazolamina) inhib secreia ionilor de bicarbonat. Celulele duetelor
pancreatice sunt ceva mai mici dect celulele acinilor glandulari i nu conin granule de
zimogen. Ele posed ns mecanisme de transport specifice n zona membranei apicale i
latero-bazale.
Fig. 31. Pancreasul i locul de vrsare a
duetelor pancreatice Wirsung i Santorini
10
Ionii bicarbonici pot difuza liber n lumenul duetelor mici. Membrana luminal a celulelor
care alctuiesc duetele de mare calibru, reabsorb bicarbonatul n schimb cu ionul de CI
-
. La un
debit secretor redus, ionii de HC0
3
" eliberai din duetele mici sunt parial nlocuii cu ionii de
CI" n duetele mari, scznd concentraia ionilor de HC0
3
-
n suc. Debitul secretor mare,
reduce timpul de schimb dintre HC0
3
-
i CI
-
i concentraia ionilor bicarbonici n suc rmne
ridicat (Fig. 33).
Clorul din sucul pancreatic provine fie din lichidul secretat de celulele acinului
glandular; fie din celulele ductului, dupa deschiderea canalului de Cl din membrana apicala.
11
Secretina care mareste productia de HCO3 , intensifica sinteza de AMPc. Unul dintre efectele
AMPc consta si in deschiderea canalelor luminale de Cl.
Penetrarea ionilor de Na din lichidul interstitial in lumenul ductului, se face printr-o
cale paracelular, de-a lungul unui gradient electrochimic.
Ionii de H rezultati din disocierea acidului carbonic, sunt expulzati din celule prin
membrana latero-bazala, datorita unui mecanism de Na/H antiport, cuplat cu gradientul
intracelular de Na. Gradientul de Na il mentine Na-K ATP-aza, situata tot in membrana
bazala a celulelor. Cei doi ioni de K patrunsi in celula in schimbul celor 3 ioni de Na
expulzati, difuzeaza in interstitiu prin canalele de K existente in membrana latero-bazala (Fig.
34).
Schimburile de ioni descrise imprima sangelui venos care paraseste pancreasul un pH
mai acid decat cel al sangelui arterial.
Apa este atrasa osmotic in ducte, prin caile paracelulare. Osmolaritatea sucului
pancreatic se apropie foarte mult de cea a plasmei, in toate tipurile de secretie.
Secretia pancreatica are o mare capacitate de a tampona acidul gastric trecut in
duoden, oferind astfel protectie mucoasei impotriva eroziunii si in acelasi timp creeaza un pH
optim de actiune enzimelor digestive.
b. Substantele organice din sucul pancreatic sunt enzimele sintetizate de reticului
endoplasmatic rugos al celulelor din acinii glandulari si depozitate in portiunea apicala sub
forma de granule de zimogen. Secretia enzimelor se face printr-un proces de exocitoza calciu
dependenta.
12
Sucul pancreatic, unul dintre cele mai polivalente sucuri din organism, poate produce
singur descompunerea complet a tuturor tipurilor de alimente. Enzimeie sucului pancreatic
sunt: proteazele, lipazele i amilaza.
Dintre proteazele elaborate n pancreas menionm: tripsina. chimotripsina, elastina,
carboxipeptidaza. ribonucleaza i dezoxiribonucleaza. Ele sunt sintetizate n celulele
glandulare sub form inactiv.
Celulele glandulare ale pancreasului produc o tripsin inactiv numit tripsinogen. .Activarea
tripsinogenului are loc n intestin sub influena enterokinazei. Enterokinaza, enzima produs
de marginea cu perie a celulelor mucoasei duodenale, trece in lumenul intestinal indeosebi
sub influenta sarurilor biliare sau colecistokinine. Avand un continut ridicat de polizaharide
(cca 41%), enterokinaza nu e descompusa in intestin, inainte de a-si exercita efectul activator.
Enterokinaza si ionii de Ca++, transforma tripsinogenul in tripsina, pierzand din molecula sa
un peptid format din 6 aminoaciz. Tripsina (cu o greutate de 21.000-22.000 dal-toni)
ndeplinete rolul de endopeptidaz, ce scindeaz legturile peptidice din zona captului
carboxilic a unor aminoacizi bazici (ex. arginina, lizina) i activeaz celelalte proteaze
pancreatice, inclusiv tripsinogenul, printr-un proces auto-catalitic. Lipsa congenital a
enterokinazei duce la malnutriie proteic.
n sngele circulant se gsesc inhibitori ai tripsinei cum sunt: alfa, antitripsina i alfa
:
macroglobulina.
Chimotripsinogenul n prezena unor mici cantiti de tripsina, n lipsa Ca++, este convertit n
intestin n chimotripsina. (prin pierderea din molecul a 15 aminoacizi). Chimotripsina (cu
greutate de 24.000 daltoni) acioneaz tot ca o endopeptidaz, ce scindeaz legturile peptidice
n care se gsesc angajate gruprile carboxilice ale unor amino-acizi aromatici (fenilalanin,
tirozin, triptofan).
Tripsina coaguleaz sngele, iar chimotripsina coaguleaz laptele.
Elastaza secretat ca proelastaz trece n form activ sub influena tripsinei. Proelastaz se
adsoarbe pe filamentele de elastin. care menin unite fibrele musculare din carne. Enzima
activ (greutate 25.000 daltoni) se ncadreaz ntre en-dopeptidaze i scindeaz legtura pep-
tidic din captul carboxilic ce aparine unor arninoacizi alifatici nearomatici, ca alanina,
valina, glicina. Ea poate hidroliza i hemoglobina, albumina, fibri-na i cazeina.
Carboxipeptidaza este o exopeptidaz care conine n molecul Zn. Are capacitatea de a
desprinde din lanul polipep-tidic cte un aminoacid cu gruparea car-boxilic terminal.
Sintetizat n pancreas sub form de procarboxipeptidaz devine activat n intestin sub
influena uipsinei.
Ribonucleaza i dezoxiribonucleaza descompun acizii nucleici n nucleotide.
Inhibitorii pancreatici ai tripsinei. Activarea tripsinei n interiorul pancreasului (nu n
intestin), poate duce la distrugerea organului. Celulele secretoare de enzime proteolitice ns,
produc n acelai timp inhibitori ai tripsinei.
Cnd pancreasul prezint distrugeri extinse, sau cnd un duet este obstruat, n zona lezat se
adun mari cantiti de suc pancreatic. In asemenea mprejurri, aciunea inhibitorilor tripsinei
poate s fie depit, ceea ce duce la activarea rapid a enzimelor. cu distrugerea n cteva ore
a esutului pancreatic, dnd natere pancreatitei acute hernoragice. Boala se nsoete frecvent
13
cu o stare de oc fatal, iar dac bolnavul supravieuiete, rmne toat viaa cu o insuficien
pancreatic.
Lipaza pancreatic se secret sub form activ, capacitatea ei enzimatic fiind mrit de
arninoacizi. ionii de Ca" i srurile biliare. Srurile biliare emul-sioneaz grsimile, proces
deosebit de important, deoarece lipaza are efecte numai la nivelul interfeei grsimi-ap. Cu
ct emulsionarea grsimilor se face mai fin, cu att sporete eficacitatea enzimei.
Ca structur chimic lipaza este o gli-coprotein (greutate molecular 42.000 daltoni i pH-
ul optim de aciune 6,8). Enzima se degradeaz foarte rapid la un pH acid, la cldur (chiar la
temperatura camerei) i n timpul tranzitului intestinal.
Lipaza nu poate aciona pe picturile de grsime, acoperite de agenii emulsio-nani fr
colipaz. Colipaza, o protein cu molecul mic (9500 daltoni), secretat de pancreas i
activat de tripsin, se fixeaz pe suprafaa picturilor de grsime, dislocnd agenii
emulsionani i ancoreaz lipaza. Acizii biliari adsor-bii pe interfaa grsimi-ap, confer
sarcini negative, care atrag lipaza i colipaza.
Lipaza i colipaza cliveaz molecula de trigliceride pn la monogliceride i acizi grai.
Hidroliza total a grsimilor neutre produs de lipaza i colipaza pancreatic n intestin, se
face n proporie de 40-60%, cea 10% din grsimi sunt neatacate iar restul degradate parial.
Fosfolipazele hidrolizeaz legturile esterice ale fosfogliceridelor (i au fost numite A
u
A
2
, B,
C i D, dup legtura scindat). In sucul pancreatic exist fos-folipaza A
2
cunoscut nc sub
numele de fosfatidil acil hidrolaz. Enzima se secret sub form inactiv de profosfoli-paza
A
2
, fiind activat de tripsin i ionii de Ca
++
.
Fosfolipaza A
2
descompune lecitina ntr-un lizofosfolipid (lizolecitin) i un acid
nesarurat. Lizolecitin lezeaz mem-branele celulare. In pancreatita acut fosfolipaza A; se
activeaz n duete i atac lecitina, unul dintre constituenii normali ai bilei, dnd natere
lizolecitinei. Lizolecitin contribuie la distrugerea esutului pancreatic i la necroza esutului
grsos nconjurtor. Fosfolipaza B scindeaz un acid gras al lizolecitinei pentru a forma
glicerilfosforilcolina.
Amilaza pancreatic este o alfa en-doamilaz, secretat sub form activ, similar amilazei
salivare, ce hidrolizeaz ns i amidonul nefiert sau copt sau glicogenul. (Are greutatea
molecular de 54.000 dai toni i pH-ul optim de aciune 6,9). Paralel cu formarea amilazei o
mic parte a enzimei trece n snge, de aceea dozarea amilazei din snge d indicii asupra
funciei exocrine a pancreasului, n pancreatitele acute hemoragice concentraia amilazei n
snge crete foarte mult, iar n pancreatitele cronice scade.
Amilaza i tripsina supravieuiesc n duoden i ileon un timp mult mai ndelungat. Aa se
explic de ce insuficienele pancreatice cronice se manifest la nceput printr-o malabsorbie a
lipidelor.
Pancreasul secret i glicoproteine ca de exemplu litostarina, ce are rolul de a preveni
precipitarea carbonarului de calciu (CaCO3).
14
Reglarea secreiei pancreatice
Secreia pancreatica este reglat printr-un mecanism nervos i umoral. La fel ca la controlul
secretiei gastrice, n reglarea secretiei pancreatice se pot distinge trei faze: faza cefalica, faza
gastrica si faza intestinal.
Faza cefalica
Reglarea nervoas a secreiei pancreatice se poate demonstra n faza cefalica.
Administrarea unui prnz fictiv la animale, produce o secreie pancreatica bogata n
enzime,ns redus ca volum. Stimulul care iniiaz secreia este deglutiia. Secreia dispare
dac se secioneaz nervii vagi.
Excitarea experimental a nervilor vagi determin o secreie redus ca volum, ns bogat n
enzime. Neuronii parasimpatici postganglionari elibereaz n gland att acetilcolina ct i
polipep-tidul vasoactiv intestinal (VIP). Cei doi mediatori acioneaz sinergie.
Pe membrana latero-bazal a celulelor acinilor glandulari se gsesc receptori pentru
acetilcolina, colecistokinin (CCK) i gastrin. Cuplarea substanelor amintite cu receptorul
specific iniiaz activarea fosfolipazei C, ce hidrolizeaz fosfatidilinozitolul din membran, cu
formarea de inozitol trifosfat (IP3) i diacil-glicerol (DAG). IP
3
deschide canalele de calciu din
reticulul endoplasmic, favoriznd trecerea acestuia n citoplasm. Creterea concentraiei
intracelulare de Ca
++
iniiaz exocitoza enzimelor din granulaii. Cei doi mesageri de ordinul
II, IP
3
i DAG exercit de multe ori efecte asemntoare.
Se presupune c exist nc o cale prin care DAG-ul ar putea modula nivelul calciului
intracelular. Creterea explosiva a concentraiei calciului intracelular, dup stimularea
receptorilor, depete necesitile activrii celulare. Dac nivelul calciului citoplasmatic n-ar
fi controlat, celula ar putea fi distrus. DAG-ul activnd proteinkinaza C, stvilete sporirea
excesiv a calciului din celule.
Proteinkinaza C acioneaz pe dou cai: pe de o pane inhib hidroliza fosfolipide-lor din membran, sursa IP
3
i
DAG, pe de alt parte nltur calciul din celule prin intervenia pompelor de calciu din membrana celular sau prin
pomparea calciului napoi n reticulul endoplasmic.
Stimulii vizuali sau olfactivi prin reflexe condiionate pot iniia secreia pan-creatic.
Faza gastric. Distensia stomacului determin prin mecanisme nervoase i umoraie secreia pancreatic.
Secionarea vagilor sau atropin reduc parial secreia produs prin distensia pereilor gastrici, plednd pentru existena
unui reflex ga-stro-pancreatic, vago-vagal. Gastrina eliberat de mucoasa antrului piloric, constituie un excitant
umoral al secreiei pancreatice care intervine in faza gastrica.
Faza intestinal. Primul hormor, intestinal descoperit, care intervine n reglarea secreiei pancreatice a fost
descris n 1902 de Bayliss i Starling. Ei au artat c introducerea unui acid, a unor aminoacizi, acizi grai sau peptone
ntr-o ans intestinal denervat, declaneaz o secreie pancreatic abundent i foarte alcalin. Au denumit substanta
stimulatoare , ce se elibereaza mai ales din mucoasa duodenala : secretina. Secretina purificata contine 27 de
aminoacizi, dintre care 14 sunt identici si in aceeasi pozitie ca in glucagon. (Are o greutate de 3200-3500 daltoni).
Structura secretinei se mai aseamana cu a polipeptidului vasoactiv intestinal (VIP) si a polipeptidului inhibitor gastric
(GIP). Hormonul actioneaza asupra celulelor epiteliale din duetele intercalare, mrind concentraia AMPc in-
tracelular, secreia de bicarbonat i trecerea apei n lumenul duetului. Secreia pancreatic provocat de secrelin este
voluminoas i alcalin. Sucul pancreatic foarte alcalin, elaborat prin intervenia se-cretinei, contribuie la neutralizarea
chimului gastric (Fig. 35).
15
Eliberarea hormonului are loc din celulele S localizate n profunzimea glandelor mucoase ale intestinului superior
oridecteori pH-ul intestinal coboar sub 4,5 sau la om sub 3. Secretina stimuleaz de-asemenea secreia biliar, a
glandelor Brunner i secreia de insulina .
In 1945 Harper si Raper descriu un alt hormon ce provine din celulele I ale mucoasei
primei poriuni a intestinului subire, ce este lansat n circulaie sub influena unor produi de
digestie: amino-acizi (mai ales fenilalanina, valina, metionina. triptofanul). peptide, acizi grai
cu lanuri lungi, ca acidul palmitic, stearic i oleic. Hormonul a fost numit pancreozimin.
Administrarea i.v. a pan-creoziminei iniiaz o secreie bogat n enzime. redus ca volum.
Pancreozimin are aceeai structur ca i un alt hormon numit colecistokinin i de aceea se
numete colecistokinm-pancreozimin, notat prescurtat CCK-PZ sau numai colecistokinin
(CCK). Hormonul conine n structura sa 58, 39, 33, 12 sau 8 amino-acizi. (Octapeptidul C
terminal pstreaz nc o intens activitate stimulatoare). Pentapeptidul C terminal este identic
cu cel din structura gastrinei. In duoden i jejun se secret mai ales CCK8 i CCK12, iar
CCK8 i n creier. Timpul de njumtire a CCK n circulaie este de 5 min. Receptorii
periferici pentru CCK sunt receptori CCK-A iar n creier se gsesc att receptori CCK-A ct
i receptori CCK-B. Stimularea ndeosebi a receptorilor CCK-B genereaz senzaia de sai-
etate.
Colecistokinin se consider a fi cel mai puternic activator al proliferrii celulelor pane re
ance. De aceia, CCK ar putea fi utilizai n tratamentul bolnavilor cu pancreatita cronic.
Aciunea purgativ a sulfatului de magneziu i a altor sruri, atribuit efectelor osmotice, se
explic cel puin n parte, prin eliberarea de CCK. care exercit efecte exdto-aotorii i asupra
musculaturii intestinale.
16
Un decapeptid extras din pielea broatei Hyla caerulea numit caerulein are o structur
asemntoare CCK i se folosete pentru stimularea secreiei pan-creatice.
Celulele F ale pancreasului endocrin stintetizeaz polipeptidul pancreatic (format din 36
aminoacizi nrudit cu peptidul YY i neuropeptidul Y), care se secret dup ingerarea de
proteine, n perioadele de nfometare, efort fizic i hipoglicemie acut. Secreia sa se afl sub
control coli-nergic. Se pare c se opune activitii CCK, n sensul c reduce producia de
tripsin i ncetinete absorbia intestinal. Concentraia polipeptidului pancreatic crete n
sngele circulant la bolnavii cu tumori ale pancreasului endocrin. Deoarece dozarea
polipeptidului se face uor, ajut la diagnosticul precoce al bolii.
Faza intestinal a secreiei pancreatice este exclusiv umoral. In timpul secreiei active, din
poriunea exocrin a pancreasului se elibereaz bradikinina, ce intensific circulaia sanguin.
Sinteza enzimelor din sucul pancreatic se adapteaz regimului alimentar. Alimentaia
bogat n glucide crete producia de amilaz dup 3-5 zile, iar aportul mrit de cazein, cea
de chi-motripsin.
Pancreatectomia sau ligatura canalelor excretoare la animale, determin pierderea din
greutate i dup 5-15 sptmni, o infiltraie gras a ficatului. Tulburarea este consecina
absorbiei defectuoase a metioninei i colinei, din cauza absenei proteazelor i lipazelor
pancreatice. Structura normal a ficatului poate fi restabilit prin ingerarea de metionina,
colin, suc pancreatic sau tripsin cristalin.
Explorarea secreiei pancreatice
Explorarea secreiei pancreatice n clinic se face prin: 1) Examinarea materiilor fecale. In
insuficiena pancretica materiile fecale conin fragmente de fibre musculare nedigerate,
granule de amidon i picturi de grsime. Materiile fecale pot include substanele amintite i
n sindromul de malabsorbie. Deoarece pancreasul reprezint principala surs de lipaz n
intestin, insuficiena pancretica exocrin se traduce printr-o lips de digestie a lipidelor i
steatoree (pierderea grsimilor prin materiile fecale). O per soan sntoas supus unui regim
ali mentar obinuit, cu cea 100 g grsimi zilnic, evacueaz prin fecale 6-7 g lipide n 24 ore.
Bolnavii cu insuficien exocrin a pancreasului ajung s elimine pe aceeai cale, pn la 50 g
lipide n 24 ore.
Tulburarea digestiei lipidelor devine manifest la distrugerea a peste 85% din glanda
exocrin. 2) Dozarea enzimelor pancreatice n snge. De obicei se determin amilaza i lipaza
sanguin. 3) Examinarea sucului pancreatic. Sucul pancreatic se poate recolta cu ajutorul unei
sonde duble. Unul din tuburi este astfel potrivit c rmne n stomac i aspir continuu
secreia gastric, celalalt ajunge pn n regiunea ampulei lui Vater. Stimularea secreiei
pancieatice se face cu colecistokinina (CCK) 2 U/kg corp/or,
urmata de reeokarea timp de 20-40 min a sucului. Se injecteaza apoi secretina tot 2
U/kg/ora si se continua recoltarea 80 min.
Volumul se apreciaz cantitativ i din suc se dozeaza enzimele. 4) Explorarea *in vivo* a
activitatii lipazei pancreatice. Proba se efectueaza dand persoanei sa ingere trioleina marcata
cu iod radioactiv. In caz de insuficienta pancreatica absorbia grsimii marcate nu se produce,
aceasta eliminndu-se n totalitate prin materiile fecale.
17
Niciunul dintre factorii excito-secretori ai pancreasului exocrin nu determin un rspund
funcional maxim, motiv pentru care testele de explorare pancreatice nu ofer indicii asupra
rezervelor funcionale ale organului.
FICATUL
Ficatul, gland anex a tubului digestiv, este cel mai mare organ intern, care cntrete la
adult 1500 g, adic aproximativ 2% din greutatea corporal. El ndeplinete funcii multiple,
imposibil de separat de cele ale altor organe i sisteme.
Structura
Unitatea morfofuncional a ficatului, lobului hepatic, are form cilindric sau hexagonal,
cu lungimea de civa mm i cu diametrul de 0,8-2 mm. Ficatul de om conine ntre 50000 i
100000 lobuli i cea 300 miliarde de celule (Fig. 36).
Lobului este construit n jurul venei centrolobulare, care se vars n venele hepatice, apoi n
cele suprahepatice i se dreneaz n cava inferioar. Spre vena centrolobular converg, ca
spiele unei roi, lamele sau plci de celule hepatice (hepatocite), care pe seciune apar sub
form de cordoane. n grosimea lamelelor intr unul sau dou rnduri de celule. Dou
hepatocite adiacente delimiteaz un canalicul biliar, care se vars n canalicu-
lul biliar perilobular.
Canaliculele biliare intraiobulare, cu un diametru de aproximativ l micromet ru, n-
au membran sau perete propriu, ci suprafaa lateral a membranei hepatocitului, n zona sa
mijlocie, servete i ca membran a canalicuiului. Jonciunile strnse dintre
18
JONCTIUNEA NEUROMUSCULARA
Contractia musculara se poate obtine dupa excitarea directa sau indirecta, prin
stimularea nervului motor. Axonul neuronului motor se ramnifica in apropierea muschiului,
fiecare ramnificare a axonului venind in contact cu o singura celula musculara. Contactul cu
fibra musculara se face in zona mijlocie a fibrei.
Fibrele nervoase impreuna cu fibrele musculare formeaza o unitate motorie
UNITATE NEUROMUSCULARA.
La muschii rapizi, cum sunt muschii externi ai globilor oculari, ai laringelui, unitatea
motorie este formata dintr-un motoneurion si 3-6 fibre musculare. Unitatea motoare poate
avea pana la 1000 fibre musculare muschii spatelui (mentinerea posturii).
Fiecare unitate motoare ar avea in medie cca. 150 fibre musculare (in organism).
Unitatile motorii lente sunt comandate de motoneuronii de dimensiuni mari. O terminatie
nervoasa de la o unitate motoare lenta, implantata intr-o unitate motorie rapida sau invers,
determina transformarea unitatii motorii (cea lenta devine rapida si viceversa). Aceasta
transformare se datoreaza frecventei descarcarii electrice care activeaza anumite enzime si
transforma fibra musculara din rapida pe lenta (sau invers).
Zona de jonctiune dintre FN (fibra nervoasa) si FM (fibra musculara) se numeste
jonctiune NEUROMUSCULARA si este de fapt o sinapsa. Terminatia nervoasa se aseaza intr-
un sant al sarcomerului. Partea terminala a neuronului constituie zona PRESINAPTICA iar
membrana musculara (sarcolema) alcatuieste membrana POSTSINAPTICA sau placa
terminala. PLACA TERMINALA se prezinta ca o membrana cu falduri care maresc suprafata
membranei postsinaptice. Intre cele 2 membrane exista un spatiu liber (fisura sinaptica)
avand o latime de 20-30 nm. FN sunt fibre colinergice iar ACO este ingramadita in veziculele
clare din apropierea fisurii sinaptice.
Monomerul transmite potentialul de actiune spre periferie , se insoteste de cresterea
concentratiei Ca
2+
care determina deplasarea veziculelor la membrana presinaptica,
separarea lor si eliberarea continutului in fisura. Actiunea ACO se exercita asupra R nicotinici
din membrana postsinaptica. Acestia sunt de fapt canale alcatuite din 5 subunitati care se
deschid cand ACO se fixeaza pe receptor.
Are loc deschiderea unui canal cu diametru de 0,65 nm prin care patrunde Na de la
exterior spre zona postsinaptica determinand depolarizarea membranei.
Cantitatea de ACO eliberata la un singur impuls e suficienta pentru a activa receptorii
postsinaptici si pentru a da nastere la potentialul propagat. De la valoarea de -90
19
mv(potential de repaus) se produce o modificare de 40 mv , potentialul crescand la -50 mv si
declanseaza potential propagat.
Aceste modificari constituie potentialul de placa terminala (de jonctiune). Actiunea
ACO este de scurta durata; in jonctiune exista acetilcolinesteraza ce descompune ACO in 1
ms . Din zona placii terminale, potentialul se raspandeste in intreaga fibra prin sarcolema.
Blocarea jonctiunilor neuromusculare se face cu diferite substante: TOXINA
BOTULINICA inhiba eliberarea mediatorului si produce paralizie (2-3 gr sunt fatale). Alte
substante sunt cele care blocheaza receptorii nicotinici: CURARA (se utilizeaza la anestezie)
sau NICOTINA
SUCCINILCOLINA este un fals mediator. Ea determina o contractie de scurta durata
insa nu poate fi descompusa de colinesteraza si astfel blocheaza jonctiunea.
Se utilizeaza numai atunci cand se asigura respiratie artificiala (in aceste cazuri se
administreaza o doza unica de anestezie si cu ajutorul curarei se obtine relaxarea)
Contractiile musculare apar dupa blocarea COLINESTERAZEI. Apar dupa administrarea
de EZERINA, NEOSTIGMINA, FIZOSTIGMINA. In practica medicala aceste substante se
utilizeaza in tratarea paraliziei musculare a musculatoruii netede intestinale (ilens paralitic)
Blocarea colinesterazei se poate produce cu INSECTICIDE. Ele determina o degradare
ireversibila a colinesterazei pe cand celelalte determina o degradare reversibila. In
degradarea ireversibila se acumuleaza multa acetilcolina => contracturi ce pot fi fatale.
Daca inregistram potential in zona postsinaptica a unui muschi in repaus -> mici
potentiale numite POTENTIALE MINIATURALE care se datoreaza deschiderii spontane a unor
canale de ioni. Modificarea de potential este mica (0,5 1,2 mv). Jonctiunea neuromusculara
are anumite particularitati:
- datorita potentialului de placa terminala, raspunsul cantractiei apare si fara
necesitatea sumarii potentialului de actiune.
- o alta diferenta este ca actiunea ACO se restrange la 1 ms si nu exista sinapse
inhibitorii neuromusculare ( nu exista mediatori de tip inhibitor)
- degradare rapida a ACO previne reexcitarea muschiului
Legea tot sau nimic
20
1871 BOWDICH a facut experiente pe inima si a observat ca: orice stimul peste
valoarea prag declanseaza concentratii maxime ale miocardului. Legea tot sau nimic afirma
astfel ca orice variatie a stimulului peste valoarea prag poate creste forta de contractie. S-a
observat ca fibra musculara izolata daca este stimulata peste valoarea prag se contracta in
totalitate. Miscarile gradate sunt totusi posibile prin antrenarea in activitate a unui numar
variabil de unitati motorii.
EFECTUL DE TREAPTA
O inima in repaus stimulata repetitiv se contracta din ce in ce mai puternic pana ajunge la un
nivel constant =efectul de treapta (+). Daca se raresc stimulii, amplitudinea constractiei
scade progresiv si efectul de treapta este (-).
Efectul de treapta + are o importanta deosebita in musculatura striata (scheletica).
Dupa 30-50 de secuse musculare performanta muschiului se imbunatateste aproape la dublu
(ex: sportivii fac miscari de incalzire inainte).
Efectul de treapta + se explica prin aceea ca indepartarea Ca din sarcoplasma se face
lent, incet. La ionii ramasi se adauga ionii introdusi cu noul stimul. Nivelul Ca in sarcoplasma
este alfel mai mare. Efectul de treapta nu se confunda cu TETANOSUL unde stimulii frecventi
determina semnarea contractiilor.
RASPUNSUL MECANIC AL MUSCHIULUI SCHELETIC STIMULAT
Aplicarea unui singur stimul cu intensitate peste valoarea prag determina o contractie
SECUSA MUSCULARA. Secusa musculara apare dupa o latenta de 2ms de la inceputul
potentialului de actiune si dureaza:
-in muschii rapizi: 7,5-10 msec
-in muschi lenti: 100-150msec
(potentialul de actiune dureaza 2,5-4 msec) dupa care se relaxeaza.
Stimularea cu frecventa mare a muschiului determina stimularea CONTRACTILOR din
motivul ca stimulul este aplicat in momentul relaxarii, relaxarea nu se produce si apare
SUMAREA. Raspunsul parta numele de TETANOS. In organism nu se intalnesc secuse
musculare, de obicei prin nervii motori se transmit intotdeauna salve de impulsuri si nu
impulsuri izolate.
Tetanosul -> incomplet atunci cand exista perioade de realxare
->complet cand nu exista perioade de relaxare a musculaturii intre
aplicarea stimulilor (este contractia in platou).
In timpul tetanosului complet , forta de contractie creste de 4 ori fata de secusa.(
acumularea Ca in citoplasma). In organismul uman nu se produc secuse pentru ca
impulsurile nervoase sunt repetitive si apare tetanosul. In functie de lungimea muschiului
contractat se descriu:
1.Contractii IZOTONICE : scurtarea muschiului
2.Contractii IZOMETRICE: lungimea muschiului ramane constanta dar creste
tensiunea intramusculara.
Cresterea tensiunii se explica prin intinderea elementelor elastice din muschi (din
tendoane, sarcomer). In musculatura care mentine postura lipsa de aparitie a oboselii se
21
explica prin asincronismul contractiei diferitelor unitati musculare( unele unitati se
contracta, altele se relaxeaza). Forta exercitata de muschi se aplica asupra sistemului osos
formeaza parghii si punctele de sprijin( este posibila miscarea si rezistenta la greutatile
aplicate asupra muschilor).
ELECTROMIOGRAMA
Muschiul in repaus nu produce curenti electrici. In timpul contractiei se produc
depolarizari care se sumeaza, se transmit la suprafata pielii. La suprafata pielii activitatea
electrica poate fi pusa in evidenta prin aplicarea unor electrozi dis..... cu suprafata de 1 cm
2
.
Inregistrarea obisnuita se numeste EMG. Se poate face o inregistrare cu un sistem
coaxial de electrozi. Sistemul coaxial este alcatuit dintr-un ac de seringa in care se introduce
o sarma (sarma este izolata de peretii acului iar peretii acului constituie cel de-al doilea
electrod)
Ruptura muschiului are loc atunci cand intinderea sa va depasi de 3 ori lungimea
muschiului relaxat, desprins de pe insertiile osoase.
Cand se folosesc electrozi de suprafata se obtine o EMG globala iar cu sistemul
coaxial -> EMG elementara.
In EMG globala se evidentiaza fasciculatia musculara iar fibrilatia se poate evidentia
numai pe EMG elementara. (ea recolteaza potentiale de la mai multe unitati motorii).
In timpul unei contractii coluntare de mica amplitudine pe EMG se observa oscilatii
cu amplitudine de 0,5-2 mv (dureaza 15 ms). Cu cat forta de contractie este mai mare
contractiile au amplitudine si frecventa mai mare ( 30-59/sec) suprapuse pe un ritm de 200
de potentiale de actiune/secunda. In unele forme de paralizie dispar potentialele frecvente.
EMG ajuta la diferentierea unei paralizii de origine musculara de una de origine
nervoasa.
MUSCHIUL NETED
Are forma piriforma, cu un singur nucleu dispus central.
-Diametru:10-100 m
-Lungime: 50-200 m
Nu are striatiuni. Elementele contractile sunt reprezentate de : actina, miozina,
tropomiozina. Lipseste troponina.
Elementele contractile sunt asezate oblic in celula. Filamentele de actina se insera pe
corpii densi. Acestia se afla inserati pe membrana sau pe esafodaj proteic din sarcoplasma.
La jumatatea distantei dintre filamentele de actina se afla filamentele de miozina. Miozina
este de 2,5 ori mai groasa decat actina si are la suprafata puntile transversale. Un filament
de miozina este inconjurat de 10-20 filamente de actina.
Datorita dispozitiei oblice a elementelor contractile in timpul contractiei fibra neteda
devine globuloasa( nu se scurteaza ca fibra striata). Filamentele de actina si miozina sunt mai
lungi in muschiul neted iar reticului sarcoplasmic este redus ca volum.
22
Tuburile in T sunt prezente sub forma rudimentara pe invaginari ale membranei
musculare numite CAVEOLE. Caveolele se afla in contact cu reticulul sarcoplasmic de aceea
depolarizarea membranei si a caveolelor determina contractia muschiului neted.
In 1948, Bozler deosebeste fibrele musculare nedete in :
-multiunitare
-viscerale (simple unit. sau unitare)
FIBRELE MULTIUNITARE: sunt fibre bine conturate acoperite cu un strat de glicoproteine
care le izoleaza una de alta. Se pot contracta si individual. Se gasesc in :
-iris, corpii ciliar
-vasele sangvine mari
-muschii piloerectori
-mucoasa uterina
In timpul travaliului, ele se transforma in fibre de tip visceral(stabilesc jonctiuni lacunare
intre ele, se contracta ca un SINCITIU).
Musculatura multiunitara are o inervatie:
-simpatica
-parasimpatica
Contactul dintre firba musculara si cea nervoasa se face in niste santuri ale membranei astfel
se aseamana cu placa neuromusculara.
Fibrele repetative prezinta o multime de dilatatii pe traseu. In zonele dilatate se
gasesc granule cu mediatori. Zona varicoasa (dilatata)nu are teaca Schwann. Cand transmite
impulsul din fiecare zona varicoasa se elibereaza mediatorul => activarea mai multo fibre.
Exista si fibre nervoase care se termina la distanta fata de fibra musculara.
Mediatorul nu trebuie sa strabata in aceste conditii o anumita distanta pana ajunge la
mushci ceea ce explica latenta de 50-150 ms(+).
FIBRELE VISCERALE : au contururi intense, se afla situate in viscerele cavitare ( interstin,
stomac, retina etc) si vasele mici.
Inregistrand variatiile de potential din interiorul fibrelor se observa ca atat variatiile
de potential cat si contractiile se produc sincron. Contractia concomitenta a mai multor fibre
viscerale se explica prina ceea ca ele sunt conectate prin jonctiuni lacunare si astfel impulsul
trece cu usurinta de la o jonctiune la alta. Ele reactioneaza ca un sincitiu (ca o singura
unitate).
Potentialul de repaus a fibrelor viscerale variaza intre -50mv si -60 mv (medie-55)
Valoarea mai mica a potentialului de repaus se datoreste permeabilitatii mai mari a
membranei pentru Na, K. Potentialul de repaus este insa inconstant. Cand oscilatiile ce apar
ajung la -35mv se produc potentiale de actiune ce pot ajunge pana la +10 mv ( potentialul de
actiunea dureaza 10-50 ms) iar contractia declansata de potential poate avea o durata de 3-
30 ms.
Tot la fibrele viscerale putem avea potentialul de actiune in platou care dureaza 1000
msec si in paralel cu el si contractia ( in ureter, uter). Undele lente, sinuoase care determina
instabilitatea potentialului de repaus se produc fie datorita unei activitati intermitente a
23
ATP-azei NA K dependenta, fie datorita modificarii ritmice periodice a permeabilitatii
membranei pentru ioni. Undele lente de pe traseul potentialului de repaus reprezinta
generatori de impulsuri (pace-makers). Ca urmare fibrele viscerale se pot contracta spontan.
Exista focare de automatism care se pot deplasa de la un loc la altul.
In constractia musculaturii netede au importanta ionii de Ca. Un fragment de
musculatura neteda introdus intr-o solutie fara Ca nu se contracta. Daca se blocheaza
canalele de Na rapide si in solutie exista Ca, concentratia de produce.Exista canale voltaj-
dependente de Ca. Calciul depolarizeaza membrana si initiaza mecanismul contractil.
Elementele contractile din muschiul neted se aseamana cu cele din muschiul striat.
-miozina are activitate ATP-azica de 100-1000 ori mai lenta de aceea contractiile
musculaturii netede sunt mult mai lente
- miozina isi poate exercita efectul ATP-azic numai daca fosforileaza lantul usor de pe
capatul globular al miozinei.
Declansarea fosforilarii este datorita Ca care penetreaza in interiorul fibrei musculare
datorita gradientului de concentratie. ( in afara celulei concentratia Ca : 10
-3
mol/.., in
interiorul celulei : 10
-7
mol/..)
In interiorul fibrei musculare Ca se prinde de un polipeptid format din 148 de
aminoacizi: CALMODULINA (greutate moleculara 167000 Daltoni), care se afla raspandita in
mediul animal si vegetal. Pe o molecula de calmodulina (arata ca un trifoi cu 4 foi) se fixeaza
4 molecule de Ca.
Calciul si calmodulina se fixeaza pe o enzima, kinaza lantului usor al miozinei si
formeaza un complex ( de pe capatul globular pe care-l activeaza). Complexul fosforileaza
lantul usor (detaseaza din ATP un fosfat pe care il ataseaza lantului usor) si numai apoi se
poate produce contractia.
In reglarea concentratiei un rol important il detine MIOZINA ( in musculatura
scheletica controlul il detine ACTINA) Rolul troponinei in contractia muschiului scheletic este
preluat de calmodulina in contractia musculaturii netede. Corpii densijoaca roul membranei
Z.
Relaxarea muschiului neted se face prin defosforilarea lantului usor si indepartarea
calmodulinei din sarcoplama. Defosforilarea se realizeaza prin interventia unei enzime L
FOSFATAZA MIOZINEI . Uneori dupa defosforilare muschiul neted ramane contractat.
Relaxarea s-ar produce in momentul detasarii Ca de calmodulina si expulzarea lui din
sarcoplasma.
M.viscerala se contracta in momentul intinderii. Intinderea determina si marirea
permeabilitatii canalelor ptr Ca si declansarea contractiei. Intinderea de lunga durata poate
sa produca o relaxare a organului = RELAXARE RECESIVA. Canalele de Ca se inchid treptat
astfel incat concentratia Ca este adusa la nivel de repaus.
Alungirea fibrelor poarta denumirea de RELAXARE RECEPTIVA si constituie o
proprietate numita PLASTICITATE (miofilamentele de actina si miozina sunt mai lungi).
Contractia fibrelor musculare netede se produce atat sub influenta mediatorilor cat si
sub influenta hormonilor.
24
In functie de localizarea musculaturii netede si de impulsurile repetative transmire,
rezultatul poate fi depolarizarea sau hiperpolarizarea. Astfel impulsurile simpatice transmire
caselor determina contractia iar cele transmire musculaturii intersinale determina relaxarea
musculaturii intestinale.
Efectul depinde si de receptorii de la suprafata membranei.
R. ptr mediatorii simpatici:
-alfa: asupra lor actioneaza noradrenalina si adrenalina in doze mari.
-Beta Beta 1 in miocard, actionati de NA
- Beta 2 (vase, bronhii) actionati de adrenalina cu efecte de obicei inhibitorii
asupra musculaturii.
(parasimpatic =excitator, simpatic = inhibitor)
Daca se stimuleaza simpaticul raspandit in intestin ( are loc o stimulare a R alfa si beta
=> inhibitia activitatii motorii. In cazul stimularii receptorilor beta are loc trecerea Ca din
sarcoplasma in reticulul sarcoplasmic iar daca se activeaza receptorii beta se produce
expulzia Ca din celula in lichidul interstitial. (determina tensiunea musculara).
Impusurile parasimpatice determina o accelerare a miscarilor intestinale prin
depolarizare. Asupra vaselor, parasimpatice are un efect relaxant indirect prin eliberarea de
NO (oxid nitric).
Hormonii sexuali feminini:
-estrogenii maresc excitabilitatea fibrelor musc.
-progesteronul hiperpolarizeaza fibra musc si reduce excitabilitatea la stimulii din
exterior.
DEOSEBIRI INTRE M. NETEDA SI CEA STRIATA
M. neteda poate fi stimulata atat de mediatori cat si de hormoni, pe cand cea
scheletica numai de mediatori (ACO). Mediatorii si hormonii pot nu numai excita fibrele, dar
o pot si inhiba.
Denevrarea m. neted nu este urmata de atrofierea lui, ca la musculatura scheletica.
Muschiul neted prezinta o hipersensibilitate la mediatori chimici (Na, A in excess) fiindca in
muschi scade concentratia de monoaminoxidaza(MAO) dupa denevrare.
Indepartarea catecolaminelor din lichidul interstitial se face prin recaptare. Muschiul
denevrat nu mai recapteaza.
OBOSEALA MUSCULARA
Oboseala = reducerea capacitatii functionale a unui sistem , reversibila prin repaus.
In contractia musculara voluntara ......mai multe elemente:
-comanda corticala
-calea de transmitere pana la muschi
-muschiul propriuzis in care se activeaza mecanismul contractil.
Oboseala musculara poate fi cauzata de afecatrea unuia dintre cele 3 nivele:
-Oboseala de tip central
-Oboseala de transmitere
-Oboseala a muschiului periferic
25
Perturbarea factorilor care suplimenteaza muschiul cu substante energetice
favorizeaza instalarea rapida a oboselii musculare. Aprovizionarea muschiului cu subst
energetice depinde de circulatie, respiratie.
1.Oboseala de tip central
In capacitatea SN de a elibera impulsuri suficiente pentru a mentine activitatea la un
anumit nivel sau pentru a atinge nivelul maxim de contractie.
Se caracterizeaza prin reducerea perceptiilor senzitive si senzoriale, diminuarea
atentiei, dificultatea si lentoarea luarii deciziilor.
Cauze: Efort fizic istovitor
Incordarea atentiei pe durata lunga
Activitate in mediu monoton
Lipsa de confort termic
Stres
Oboseala poate disparea instantaneu prin schimbarea tipului de activitate ( nu este
vorba de o oboseala musculara periferica intrucat daca sunt stimulati muschii se contracta).
In efortul istovitor oboseala este produsa de disparitia depozitelor de substante energetice
din muschi si de aparitia catabolitilor.
Factorii amintiti stimuleaza R-> impulsurile se transmit la scoarta unde se formeaza
deprimarea activitatii corticale. Prin actiune coluntara corticala se antreneaza in miscare
alaturi de muschiul obosit alte grupe musculare. Rezulta imprecizia miscarii.
2.Oboseala de transmitere
Se datoreaza epuizarii stocurilor de mediatori din jonctiunea neuromusculara si ai
capacitatii nervului de a sintetiza alti mediatori. Excitarea directa a muschiului sa nastere la
raspuns contractil (miastenia este o oboseala de transmitere). Poate fi reprodusa prin
mobilizarea unor substante ca fizostigmina, exerina, reostigmina care blocheaza
colinesteraza si ACO nu se desc.
3.Oboseala periferica
Se datoreste epuizarii stocurilor de glicogen cu acumularea de acid lactic, fosfati in
muschi si scaderea K intracelular. Epuizarea depozitelor de glicogen (glucoza) determina o
reducere a formarii de substante macroergice fosforilate (ATP, CP) fiind impiedicata totodata
si trecerea Ca din reticul in sarcoplasma. Catabolitii: din acidul lactic se disociaza H , fosfatul
se porduce prin eliberarea lui din ATP.
H are efecte negative asupra mobilitatii puntilor transversale si inhiba activitatea
normala a enzimelor de glicoliza ( nu se poate reface ATP, CP). Scaderea K se produce cu
fiecare potential de actiune in muschii scheletic. Ionii de K sunt eliminati din celula in
momentul repolarizarii fibrei.
Cresterea concentratiei de K in intestin modifica potentialul de repaus in celula. In
plus K scazut intracelular are efecte asupra mobilizarii Ca din sarcoplasma.
In trecut se credea ca oboseala se datoreaza scaderii ATP. Cu toate acestea s-a
observat ca oboseala se poate instala si atunci cand in muschi exista o cantitate suficienta de
ATP. Scaderea numai a ATP-ului determina contractura musculara.
Nu esista o metoda obiectica de masurare a ATP.
26
In repaus, depozitele energetice se refac.
Un alt organ efector =>Glandele
Secretia glandulara: glandele exocrine produc atat enzime cat si lichid incarcat cu electroliti.
Productia de enzime are la baza acelasi mecanism ca si productia de proteine. Enzimele sunt
de fapt Proteine. Ele iau nastere din aminoacizi. Asamblarea a.a in polipeptide are loc in
ribozom. Proteinele trec apoi in reticulul endoplasmatic si in 30 in aparatul Golgi. Aici sunt
ambalate in membrana. Ele se depoziteaza sub forma de granule in citoplasama. Din granule
enzimele sunt puse in libertate sub influenta impulsurilor nervoase care ajunse la glande
maresc permeabilitatea membranei celulare ptr Ca.
Enzimele se elibereaza prin EXOCITOZA Ca dependenta.
Glandele exocrine produc si un lichid bogat in electroliti.
Sub influenta impulsurilor parasimpatice are loc o crestere a permeabilitatii
membranei celulei glandulare pentru Cl care patrunde in interior si potentialul coboara de la
-30 mv la -40 mv. Datorita incarcaturii (-) din interior, Na
+
este atras si el in celula. Se
formeaza NaCl. Creste presiunea osmotica si atrage H2O . Apa si electrolitii trec in lumenul
glandelor antrenand si substante organice, inclusiv enzime.
27
NEURONUL
Este unitatea morfofunctionala a SN.
SNC are 10
11
neuroni iar celulele gliale sunt in numar de 10-50 de ori mai numeroase.
Neuronul este alcatui din :
-corp celular sau PERICARION
-2 tipuri de prelungiri: -subtiri, arborizate: DENDRITE
- o prelungire groasa, unica, care nu se ramnifica decat in
portiunea terminala: AXON
Forma pericarionului : stelata, sferica, rotunda, fusiforma, piriforma, piramidala.
In functie de numarul de prelungiri, neuronii pot fi : unipolari, bipolari, multipolari,
pseudopolari( au o singura prelungire care ulterior se ramnifica intr-o ramura centrala si una
periferica).
Neuronul este acoperit de membrana celulara.
1. Corpul celular: - este acoperit la exterior de o membrana celulara (NEURILEMA). In
interiorul celulei exista organite celulare comune : reticul endoplasmatic, ap.Golgi,
mitocondrii si organite celulare specifice.( corpii Nissl, neurofibrile)
Corpii Nissl( sau coprii tigroizi) sunt raspanditi in intreaga citoplasma si in portiunea
lor de emergenta a dendritelor, se pot vizualiza in celulele de dimensiuni mari prin
colorare cu coloranti bazici cand apar asemeni unor pete de culoare (blana de tigru).
C.Nissl contin ARN . In realitate CN sunt ribozomi care participa la sinteza de
proteine. Gh Marinescu a deschris in anul 1901 procesul de TIGROLIZA:
-disparitia din citoplasma a CN. Procesul survine odata cu imbatranirea celulelor,
dupa suprasolicitarea organismului sau dupa imbolnavirea axonului. Tigroliza
instalata dupa sectionarea axonului denota ca prelungirile periferice exercita o
influenta asuprea corpului celular.
Neurofibrilele: sunt prezente in corpul celular dar si in prelungiri. Ar detine un rol in
transmiterea impulsurilor nervoase iar recent s-a descris si un rol mecanic de
sustinere.
Nucleul : contine nucleoli, lipsesc centriolii. Neuronii nu se pot multiplica. In
momentul in care sunt distrusi se inlocuiesc cu celule gliale.
2. Prelungirile neuronale
Axonul este invelit intr-o teaca de mielina compusa din lipide (sfingozida) si proteine.
Mielina: este considerata cel mai bun izolator electric cunoscut. In nervii periferici teaca de
mielina o produc celulele Schwann. In timpul dezvoltarii embrionare , celulele Schwann
inconjoara CIUNDRAXUL(AXONUL) de mai multe ori. Mielina contine 100 straturi suprapuse
(poate ajunge pana la 300).
Mielinizarea N.Senzitiv incepe intrauterin (luna 4-5) iar a N.Motori (corticospinali) se
produce in perioada extrauterina(dupa luna 5). N.Motori ajung complet mielinizati cand
copilul incepe sa umble(varsta de 2 ani).
28
Mielina de pe fibrele nerv. periferice se extinde pe pertiuni de 1mm, din loc in loc
existand intreruperi de 1m: STRANGULATII RANVIER.
Celulele Schwann produce mielina care este dispusa intre 2 strangulatii Ranvier. In
mod obisnuit teaca de mielina este acoperita de celule Schwann. Celulele Schwann detin un
rol important in regenerarea neuronului. In SNC mielina este produsa de OLIGODENDROGLII
(produc mielina ce acopera mai multe fibre nervoase invecinate).
Deasupra tecii Schwann exista o membrana foarte subtire, cu alcatuire din tesut
conjunctiv numita teaca lui HENLE (Kay-Retzius). Teaca Schwann se mai numeste si
NEURILEMA.
Proteinele sunt sintetizate numai la nivelul corpului celular. De asemenea in
prelungirile neuronale mitocondriile sunt absente. Astfel desprinderea prelungirilor de
corpul celular provoaca degenerarea neuronului.
TRANSPORTUL NEURONAL
In axon au fost descrise mai multe tipuri de transporturi de organite si substante:
1. Transportul ANTEROGRAD (axo-plasmatic)
Se disting 2 astfel de tipuri de transport: unul cu viteza rapida si unul cu viteza lenta. Daca se
aplica o ligatura pe axon, substanta se acumuleaza deasupra ligaturii.
a. Transport RAPID: ajunge la o viteza de 400 mm/zi. Prin transport rapid sunt deplasate
la periferie organitele celulare( mitocondrii) pe care le gasim si in butonii sinaptici.
Deplasarea rapida are loc in prezenta microtubulilor( tubi lungi, dispusi de-a lungul
axonului, cu diametru de 15 m). Sunt formati din 2 proteine:
-TUBULINA alfa
-TUBULINA beta corelata cu functia ATP-azica.
Transportul rapid necesita ENERGIE. Este mediat de o proteina: KINEZINA. Kinezina
are o extremitate proximala cu 2 capete. 1 capat se prinde de microtubuli iar celalalt
de organitul celular. Dupa atasare, corpul se inclina permitand atasarea de
microtubuli a celui de-a doilea capat. Deplasarea organitului se face astfel in ritm
29
continuu. Daca se distrug microtubulii ( cu COLCHICINA) transportul rapid nu se mai
face.
b. Transport LENT: conduce proteinele spre extremitatea axonului (proteine necesare
pentru refacerea uzurilor) transportul lent depinde mai putin de prezenta
microtubulilor canad loc si dupa distrugerea acestora. Transportul lent se face de-a
lungul microfiilamentelor.
2. Transportul RETROGRAD ( se face cu viteza de 200 mm/zi)
Prin transportul retrograd ajung in interiorul pericarionului:
-organite celulare distruse
-colinesteraza
-factorii de crestere neuronala
-peroxidaza din hrean
Transportul retrograd necesita prezenta microtubulilor si este indeplinit de DINEINA (
aceasta actioneaza similar cu KINEZINA). Prin transportul retrograd ajung in pericarion si
anumiti agenti patogeni: virusul rabic, v. poliomielitei, herpetic. Prin transportul retrograd
prelungirea periferica a neuronului influenteaza metabolismul din interiorul pericarionului.
Neuronii sunt specializati in :
-receptionarea semnalelor
-transmiterea lor spre centrii nervosi
-prelucrarea informatiei
-transmiterea impulsului spre organul efector.
CELULELE GLIALE
1. Astrocitele sunt situate intracerebral. Se disting 2 tipuri de astrocite:
a. Fibroase- contin numeroase filamente intracitoplasmatice si se gasesc
situate mai ales in substanta alba.
b. Protoplasmatice care anu citoplasma ce contine numeroase granule.
Sunt situate in substanta cenusie.
Ambele tipuri de astrocite se dispun in jurul vaselor de sange participand la formarea
jocntiunilor stranse intre celulele endoteliale ale vaselor capilare si contribuind astfel la
formarea barierei HEMATOENCEFALICE. Se dispun de asemenea in jurul sinapselor si celulei
nervoase.
O alta functie indeplinita de astrocite este sinteza unor factori trofici neuronali.
Astrocitele participa la mentinerea unei concentratii ionice corespunzatoare si la
mentinerea integritatii procesului de neurotransmitere.
30
Astfel astrocitele preiau K si glutamatul pe care il convertesc la glutamina ->(neuroni
glutaminergici)
2. Oligodendrogliile sunt implicate in producerea de mielina in SNC
3. Microgliile- au rol fagocitar.
FUNCTIILE NEURONULUI
-excitabilitatea
-conductibilitatea
-degenerarea si regenerarea (in conditii patologice)
EXCITABILITATEA
DEF: intrarea in activitate a neuronilor sub actiunea unui stimul.
Se modifica permeabilitatea membranei si potentialul de repaus. Reactia oricarui tesut la un
stimul de numeste EXCITATIE.
Pentru ca un stimul sa produca excitatie el trebuie sa indeplineasca anumite conditii:
a. Sa fie suficient de intens ca sa poata determina depolarizarea
membranei(valoarea prag val a intensit stimulului de la care incepe
excitatia)
i. Stimuli supraliminari - sunt cei care declanseaza excitatia
ii. Subliminari- care sunt lipsiti de raspuns
b. Stimulul trebuie sa aiba o anumita bruschete. Cel mai utilizat stimul in
practica este curentul electric. Stimulii mai pot fi fizici, mecanici, chimici. Daca
stimulul electric creste progresiv prin intercalarea intre sursa de curent si
tesut a unui potentiometru, tesutul excitat nu mai raspunde la stimul. Se
produce asa numitul proces de ACOMODARE. Se produce un fel de crestere a
pragului de excitabilitate
c. Stimulul trebui sa realizele o anumita densitate pe unitatea de suprafata.
Daca aplicam 2 stimuli: unul punctiform si unul cu o suprafata mai mare,
excitatia porneste de la stimulul punctiform (densitatea lor pe unitatea de
suprafata este mai mare)
d. Stimulul trebuie sa actioneze o perioada de timp.
POTENTIALUL DE REPAUS
Toate celulele din organism sunt incarcate electric:
- Pozitiv la exterior
- Negativ la interior
Diferenta de potential intre interior si exterior in repaus: Potential de repaus, si poate fi
masurat cu ajutorul unui microelectrod introdus prin membrana. Valoarea PDR este de :
-90mv(muschi)
31
-70mv(neuron)
-4 la -12 mv (hematii)
La aparitia potentialului de repaus contribuie:
-pompa de Na/K dependenta de ATP
-difuziunea
-echilibrul de membrana DONNAN
Pompa Na/K dependenta este reprezentata de ATP-aza de Na-K dependenta. Ea
expulzeaza 3 Na pentr 2 K. Din cauza inegalitatii sarcinilor expulzate si primite, pompa se
mai numeste POMPA ELECTROGENERATOARE.
Cand concentratia K extracelular este mai mica, K paraseste celula, celula se incarca negativ
mai puternic si se hiperpolarizeaza.
Echilibrul de membrana DONNAN
Se produce datorita macromoleculelor proteice incarcate negativ care nu mai pot parasi
celula si determina incarcarea electrica interioare.
POTENTIALUL DE ACTIUNE
Se produce sub actiunea unui stimul. Dupa aplicarea unui stimul supraliminar are loc
o DEPOLARIZARE a membranei. Depolarizarea membranei este rezultatul cresterii
permeabilitatii acesteia pentru Na. Permeabilitatea pentru Na se mareste de cca 500-5000
de ori si Na patrunde in celula in cantitate crescuta. Astfel polaritatea membranei se
inverseaza: interiorul se pozitiveaza iar exteriorul se negativeaza.
Inregistrand suita de evenimente electrice din neuron se observa ca modificarea de
potential se instaleaza dupa o PERIOADA DE LATENT. Valoarea potentialului se deplaseaza
pana la 10-55 mv urmata de o depolarizare brusca cand se atinge valoarea de +35 - +45 mv,
urmata de revenirea potentialului spre valoarea de repaus (-70mv).
Revenirea potentialului (repolarizarea) se face brusc pana ce repolarizarea are loc in
proportie de 70% dupa care sinteza de repolarizare incetineste. Depozarizarea si
repolarizarea dureaza o perioada scurta 0,5ms.
O perioada de cca 4ms dupa repolarizar, potentialul ramane deasupra nivelului de
repaus constiutuind POST DEPOLARIZAREA sau POST POTENTIALUL NEGATIV. Dupa ce
32
potentialul a atins valoarea de repaus se constata ca el se subdeniveleaza (cu 1-2 mv) in
interval de 40-50ms sau mai mult ceea ce reprezinta POST POLARIZAREA sau POST
POTENTIAL POZITIV.
Cresterea si scadearea rapida a potentialului se cunoaste sub denumirea de
POTENTIAL de VARF sau VARIATIE NEGATIVA (0,5-1ms).
Potentialul de actiune este inclus in deschiderea canalelor de Na. Canalele sunt
captusite cu sarcini negative in interior care atrag Na. In timpul potentialului de repaus
canalele sunt inchise (la-70mv). Se presupune ca strajuirea canalului este datorata prezentei
a 2 bariere (porti) : o bariera de activare situata extracelular si o bariera de inactivare situata
intracelular, avand forma unei bile legata de canal.
Cand potentialul de membrana coboara la -55 mv se deschide bariera de activare si ionii de
Na navalesc in celula. In momentul in care potentialul se apropie de +35 mv se activeaza
bariera interna care inchide canalul si Na nu mai poate intra in celula.
In momentul revenirii la valoarea de repaus se inchide bariera de activare si se deschide din
nou bariera interna de inactivare incepand repolarizarea celulei.
33
Diametrul canalului de Na cca0,5nm. Canalele au forma ovalara iar portile sunt prelungiri
proteice care prin schimbari conformationale permeabilizeaza sau blocheaza canalul.
Pozitia barierelor este controlata cel putin pe 2 cai principale:
1. Variatiile de potential ale membranelor celulare aduc modificari conformationale ale
barierei si determina fie inchiderea, fie deschiderea ei: bariere executabile electric
sau VOLTAJ DEPENDENTE.
2. Modificarile conformationale ale proteinelor unor tipuri de bariere sustin dupa
cuplarea lor cu anumite substante. Substanta care se fixeaza pe bariera se numeste
LIGAND.
Intre liganzi se incadreaza mediatorii SNC: (acetilcolina), hormoni, factori endogeni
din celula: AMPc, Ca, proteina G. Cand ACO se leaga de canal se produce deschiderea
barierei si a unui canal cu diametrul de 0,65 nm permeabil pentru ionii pozitivi cu un
diametru mai mic. Datorita deschiderii canalelor pentru Na, potentialul de echilibru
se indreapta spre potentialul de echilibru sodic (+61mv). Nu atinge insa aceasta
valoare intrucat canalul pt Na se blocheaza (prin bariera de inactivare), interiorul
celulei se incarca pozitiv si Na nu mai este atras electrostatic in celula.
Canalele de Na pot fi blocate prin:
-anestezice locale: procaina, cocaina ,lidocaina.
-aplicarea de tetrodoxina, saxitoxina pe suprafata externa a membranei
celulare.
Incetinirea inactivarii canalelor de Na o produce veninul de scorpion si actinii ->
descarcari prelungite de potentiale in nervi si muschi cu efecte dureroase sau letale.
La producerea potentialului de actiune participa si ionii de K. Canalele de K
sunt rotunde, cu diametrul de 0,3 pe 0,5 nm, fara incarcatura electrica negativa. Are
numai o bariera tot sub forma globulara. La potentialul de repaus bariera interna este
inchisa.
Membrana celulara contine un numar de cca 9 ori mai mare de canale pentru
K deschise fata de cele pentru Na => o conductanta de 9 ori mai mare pt K decat
pentru Na.
Depolarizarea celulei determina o modificare a conformationala LENTA a
barierei cu deschiderea ei si difuziarea K spre exterior. Din cauza incetinirii cu care se
deschide canalul de K deblocarea lui care loc in acelasi timp cu activarea canalului de
Na ceea ce accelereaza procesul de REPOLARIZARE. La sfarsitul fazei de repolarizare
numarul canalelor pentru K deschise este de 15 ori mai mare decat a canalelor de Na
deschise.
Prin patrunderea Na in interiorul celulei diferenta de potential dintre
exteriorul si interiorul celulei dispare si al un moment dat se inverseaza dand nastere
la POTENTIALUL DE VARF. Trecerea Na prin membrana celulara in timpul POT DE
VARF se face pasiv fiind dependenta exclusiv de gradientul de concentratie.
Restabiliarea potentialului de repaus are loc prin restrangerea fluxului de Na
odata cu inchiderea canalelor de Na si cresterea permeabilitatii pentru K. Numarul de
34
ioni de K pierduti dintr-o fibra mielinica groasa in timpul restabilirii pot de repaus
este de 1:100 000 pana la 1: 500 000 (REPOLARIZARE). Inseamna ca pentru
DEPOLARIZAREA celulei in timpul potentialului de varf sunt necesari un numar egal
de ioni de Na.
Iesirea K din celula nu reuseste sa readuca imediat potentialul la valoarea de
repaus => POSTDEPOLARIZARE sau POST POTENTIAL (-).
Posthiperpolarizarea sau postpotentialul pozitiv se caracterizeaza prin
acumularea de Na si K la exterior si cresterea numarului de sarcini negative la
interior. Se porduce o data cu intrarea in functiune a pompei de Na.
Daca se blocheaza Na K ATP-aza posthiperpolarizarea dispare (dovada ca este
produsa de intrarea in functiune a pompei de Na).
Pe suprafata de 1 m
2
exista un canal de Na si K si 100 molecule de Na K ATP-aza
Modificarea excitabilitatii celulelor
1. Aplicarea pe nerv a unui stimul a carui intensitate creste progresiv si foarte lent
induce fenomenul de ACOMODARE.
ACOMODAREA canalele de Na se deschid progresiv astfel incat Na nu ajunge sa
declanseze pot de actiune. In schimb canalele de K intra in functiune lasand K sa iasa din
celula. Se produce astfel o echilibrare a fortelor de depolarizare si a celor de repolarizare.
2. Excitabilitatea se modifica paralel cu potentialul de actiune. In perioada potentialului
de varf fibra nervoasa este inexcitabila deoarece membrana este depolarizata.
Timpul in care celula nervoasa ramane inexcitabila reprezinta PERIOADA
REFRACTARA ABSOLUTA ( dureaza de la pot de varf pana in momentul in care
repolarizarea de face in proportie de 1/3)
PERIOADA REFRACTARA RELATIVA stimularea celulei e posibila dar numai de catre
stimuli de intensitate mare insa amplitudinea pot de actiune este mica (creste pragul
de excitabilitate). Sfarsitul perioadei refractare relative corespunde cu restabilirea
amplitudinii membranare a potentialului de actiune.
In perioada postpotentialului negativ (-) excitabilitatea este crescuta iar in
timpul PP(+) excitabilitatea este diminuata.
Frecventa potentialelor de actiune generate de un tesut depinde de durata
perioadelor refractare absolut:
35
-neuroni: excitati cu maxim 1000 stimuli/s
-muschi: excitati de max 500stimuli /s (PRA:2ms)
3. In cursul stimularii nervului cu curent electric continuu, la inchidera circuitului
excitatia porneste de la catod care aduce sarcini negative in plus si favorizeaza depolarizarea
membranei.
La deschiderea circuitului, excitatia porneste de la anod unde se creaza un
dezechilibru mai mare de ioni.
La curent subliminal in jurul catodului sunt atrase sarcini negative si excitabilitatea
tesutului se modifica, fenomen numit ELECTROTONUS.
Sub actiunea curentului electric continuu excitabilitatea nervului in jurul catodului se
mareste, fenomen cunoscut sub numele de CATELECTROTONUS. IN apropierea polului
pozitvi, excitabilitatea diminua (necesitand un stimul excitant de intensitate mai mare) :
ANELECTRORONUS.
4. Excitabilitatea variaza si in functie de frecventa stimulilor. Numarul maxim de stimuli
ce pot fi generati sau propagati de un tesut in unitatea de timp poarta numele de
mobilitate functionala sau LIABILITATE FUNCTIONALA.
Un stimul care depaseste mobilitatea functionala nu produce excitatie ci o stare
numita PARBIOZA. Curenti de inalta tensiune pot fi utilizati in terapie fara a produce
excitatii (deoarece depasesc mobilitatea functionala a tesuturilor).
Masurarea excitabilitatii tesuturilor
Se utilizeaza curentul electric
CONDUCTANTA: trecerea ionilor printr-o membrana sub influenta campului electric
continuu.
REOBAZA: intensitatea MINIMA a curentului capabila sa produca excitatia intr-un timp
nedefinit
TIMP UTIL : timpul in care un curent cu intensitatea de reobaza, produce excitatia
CRONAXIA: timpul in care un curent de 2 reobaze, produce excitatia.
Determinarea cronaxiei este metoda de electie pentru stabilirea excitabilitatii tesuturilor
CONDUCTIBILITATEA
DEF: transmiterea impulsurilor de-a lungul prelungirilor neuronului.
Fibrele nervoase nu se comporta ca un fir metalic care conduce curentul electric, in
sensul ca transmiterea are nevoie de energie.
Viteza de transmitere a impulsurilor nervoase este mai mica decat la curentul
electric, transmiterea se face sub forma de potential de actiune care isi pastreaza
amplitudinea si viteza de-a lungul intregului parcurs.
Conducerea impulsurilor se face diferit in functie de tipul de fibra nervoasa.
FIBRELE AMIELINICE: propagarea se face din aproape in aproape prin curenti
HERMAN care se raspandesc atat in suprafata cat si in interiorul fibrei nervoase. Pentru ca
potentialul sa se propage este necesara o depolarizare a membranei de 20mv. O scadere a
potentialului de repaus cu 20 mv determina propagarea excitatiei in ambele sensuri . In
biologie sarcinile electrice sunt purtate de ionii Na, K, Cl, HCO3. Prin conventie directia
deplasarii ionilor + este considerata directia curgerii curentului ( curentul trece de la + la -).
36
Viteza de transmitere a impulsurilor prin fibrele amielinice variaza direct proportional
cu diametrul fibrei. Deplasarea sarcinilor + la suprafata reduce potentialul electric favorizand
deplasarea impulsurilor.
FIBRELE MIELINICE : deplasare saltatorie de la o strangulatie la alta. Stratul de mielina
este izolator electric. Depolarizarea prelungirii nervoase are loc doar in zona stranguatiei
unde nu exista teaca de mielina.Depolarizarea din zona nodului Ranvier se datoreaza
patrunderii Na prin membrana inzestrata cu canale de Na, de cca 100 de ori mai multe decat
membrana fibrelor amielinice. Potentialul de actiune denerat se transmite de la un nod la
altul, fara intarziere la fel ca un curent electric, atat prin lichidul extracelular cat si prin
axoplasma.
Depolarizarea care se produce la nivelul unei strangulari Ranvier excitat are valoare
de cel putin 100mv. (de la -70mv la +35mv). In timpul transmiterii potentialului de 100 mv de
la o strangulatie la alta, el se reduce la jumatate, adica 50mv fiin insa suficient de intens
pentru a depolariza strangulatia. Cand ce-a de-a doua strangulatie este scoasa din functiune,
potentialul de 50mv ce se propaga la a III-a strangulatie se injumatateste din nou atingand
25mv. Variatia de potential produsa ramane suficient de puternica pentru depolarizarea
celei de-a treia strangulatii. Transmiterea saltatorie are un grad de siguranta: chiar daca
impulsul sare peste o gatuire Tanvier potentialul isi pastreaza capacitatea de a produce
depolarizarea in continuarea fibrei.
Conducerea prin fibrele mielinice este de 50 de ori mai rapida, cu consum
redus de energie. Viteza de propagare depinde de grosimea prelungirii nervoase. Cu cat fibra
este mai groasa cu atat conductibilitatea este mai mare.
37
Legile conductibilitatii
1. Legea integritatii neuronale; un neuron conduce atat timp cat este integru.
2. Legea conducerii izolate: impulsurile se conduc separat prin diferite prelungiri si nu
influenteaza conducerea fibrelor invecinate (exista si exceptii)
3. Legea conducerii indiferente: Impulsurile se conduc prin neuron si prelungirile sale in
abele directii (demonstrata de BABUHIN pentru prima data, pe pestele electric)
4. Legea conducerii nedecrementiale: potentialele de actiune sunt conduse cu aceeasi
amplitudine si cu aceeasi viteza pe tot parcursul nervului.
Clasificarea fibrelor nervoase in functie de viteza de conducere
Inregistrand potentialul de actiune intr-un NERV MIXT la distanta de locul stimularii se obtine
o curba cu mai multe deflexiuni. Deflexiunile se datoresc conducerii impulsurilor cu viteza
inegala prin fibrele ce alcatuiesc nervul. In functie de structura, firbele nervoase se clasifica
in :
1. Fibre mielinice (A , B)
2. Fibre amielinice (C)
Fibrele A
In raport de grosime se clasifica in :
-fibre mielinice A
-
-
-
Diametru lor variaza de la 1 al 20 microni iar viteza de conducere intre 5 m/s si 120 m/2.
A : aferente de la fusurile neuromusculare nervi somatici motori (neuroni motori si
senzitivi)
A : eferente de la receptorii tactili cutanati (semnale de atingere si presiune)
A : fibre motorii de la fusurile neuromusculare
A : transmiterea impulsuri termice si algice din teritoriul cutanat (fibre senzitive)
FIBRELE DE TIP B
Au diametru de 1-3m(micrometri) si viteza de conducere de 3m/s 14 m/s . Sunt
fibre vegetative preganglionare.
FIBRELE DE TIP C
Amielinice au diametrul < 1m, viteza de conducere cca2m/s. Ele formeaza fibre
vegetative postganglionare si nervii senzitivi ce conduc durerea.
38
DEGENERAREA SI REGENERAREA FIBRELOR NERVOASE
Degenerescenta neuronala a fost descrisa de Waller in 1850 si de aceea se mai numeste
degenerescenta Waloriana.
Degenerarea axonului incepe dupa sectionare prin trunchiere
Segmentarea tecii de mielina
Segmentarea axoplasmei
Dupa cca 8-32 zile teaca Schwann este golita de axoplasma si mielina datorita fagocitarii
resturilor rezultate in urma segmentarii si astfel ramane numai teaca Schwann goala.
In afara degenerescentei axonului (ANTEROGRADA) se descrie si o degenerscenta
RETROGRADA a pericarionului.
Trunchiere
Fragmentarea RE in 20-40 zile
Recuperearea neuronului incepe in zona axonului legat de pericarion cu aparitia a cca 50 de
muguri de crestere. Din acestia numai unul patrunde in teaca Schwann golita si se dezvolta
in continuare.
Viteza de crestere: 1,5 4 mm/24h
Ptr regenerare este obligatorie prezenta tecii Schwan. In SNC nu exista teaca si
regenerarea nu are loc.
Distanta dintre capetele axonale sectionate nu trebuie sa depaseasca 3mm, astfel
capatul neuronal nu ami ajunge in teaca si se incolaceste => Neuron (formarea lui explica
durerile fantoma aparute la bolnavii amputati). Formarea NEURONULUI poate fi prevenita
prin legarea fibrei nervoase.
S-a descoperit un factor natural de crestere a fibrei nervoase izolat din glandele salivare la
soricei(masculi), veninul unor serpi.
Daca se blocheaza la anumalul nou-nascut productia de factor cu anticorpi (ser imun)
la animal nu se mai dezvolta SN simpatic: simpatectomie imunologica.
Factorul de crestere favorizeaza dezvoltarea unor fibre senzitive si a unor centri
nervosi colinergici situati la baza creierului.
Recent s-a descoperit ca celulele Schwann si astrocitele produc factorul neurotrofic ciliar
(CNTF) care ar putea fi utilizat in boli degenerative motorii (ca mijloc terapeutic)
39
ORGANUL EFECTOR
Efectorul caii reflexe este reprezentat de muschi care are proprietatea de a se scurta sau de
a realiza tensiune.
In organismul uman mobilitatea nu este o proprietate exclusiva muschilor. Exista celule care
se misca:
-leucocitele: migreaza spre zona infectat
-organite celulare inzestrate cu mobilitate : cromozomii se deplaseara in timpul
diviziunilor celulare.
-cilii cu care sunt prevazute celulele epiteliului respirator se afla in permanenta
miscare.
-spermatozoizii se deplaseaza datorita flagelului cu care sunt inzestrati.
MUSCHII
Din punct de vedere morfologic si fiziologic se descriu:
1. Muschi striati: se contracta rapid sub influenta impulsului nervos si sunt fixati pe
schelet contribuind la locomotie. Datorita locomotiei, organismele animale si umane
castiga o independenta fata de mediu. Dezvoltarea musculaturii scheletice a avut
drept rezultat si o perfectionare a CN (ex: dezvoltarea vorbirii)
2. Muschi netezi: se afla situati in peretii vaselor si viscerelor cavitare(intestin, stomac,
etc)
Se contracta lent sau chiar spontan in afara organismului
3. Muschiul miocardic: este un muschi striat, se contracta mai lent decat musculatura
scheletica. Inima scoasa din organism poate sa-si continue activitatea =AUTOMATISM
CARDIAC
Muschiul striat
Aproximativ 40-45% din greutatea corpului uman o reprezinta muschii
scheletici si 10% muschii netezi.
Celulele musculare poarta denumirea de FIBRE MUSCULARE. Fibra musculara este
acoperita la suprafata de o membrana SARCOLEMA (se aseamana cu membranele
celulare sau citoplasmatice).
Ea are rolul de a raspandi excitatia din zona placii neuromusculare (SINAPSA) in toata
fibra musculara. La suprafata sarcolemei exista un strat de mucopolizaharide (MPZ)
reprezentate de proteoglicani care contine si fibre foarte fine de colagen. Fibrele de
colagen se undesc alcatuind TENDONUL care are rol functional: dirijeaza forta
contractila a tuturor fibrelor musculare asupra punctului de insertie. Tendoanele sunt
cele care favorizeaza miscarile foarte fine ale extremitatilor.
In interiorul fibrei musculare, marginita la exterior de sarcolema se afla
sarcoplasma (citolpasma) care contine, apa, substante umorale, K, MG, fosfati,
proteine.
40
In sarcoplasma se afla nucleii si organitele celulare. Musculatura scheletica dispune
de foarte multi nuclei: pe o distanta de 1 cm exista 30-40 de nuclei, fapt explicabil
prin aceea ca fibrele scheletice s-au format prin unirea MIOBLASTILOR si astfel au
aparut aceste celule multinucleate.
In sarcoplasma exista organite celulare specifice reprezentate de MIOFIBRILE (sute
sau chiar mii)
Miofibrilele sunt cele care creeaza impresia striatiilor longitudinale in fibra
musculara vizualizata la microscop.
La microscopul optic, miofibrilele prezinta alternanta de zone intunecate si zone
clare. Zonele intunecate (discuri, benzi) au o mare afinitate pentru coloranti. Discurile
clare se coloreaza in mai mica masura.
La lumina polarizata discurile intunecate sunt birefringente, anizotrope, iar
discurile clare sunt monorefringente, izotrope.
In zona centrala a discurilor intunecate apar fante mai putin intunecate
denumite zona H. In mijlocul zonei H exista o linie mai intunecata numita linia M. In
zona centrala a discurilor clare se gaseste o linie intunecata denumita linai Z sau
membrana Z. Ea are rolul de a fixa discul clar de membrana si de a-l mentine la
acelasi nivel in miofibrile invecinate.
Intre doua linii Z se delimiteaza un SARCOMER: format dintr-o banda
intunecata (central) si 2 jumatati de discuri clare.
Miofibrilele sunt formate din miofilamente. Miofilamentele pot fi (evid. la
microscop electronic):
- Groase MIOZINA
- Subtiri ACTINA, TROPOMIOZINA, TROPONINA
Filamentele groase au diametru de 10nm si lungime de 1,6 m. La suprafata lor
se gasesc niste excrescente: punti transversale.
Miozina se intinde in zona discurilor intunecate. Pe membrana Z se afla
miofilamente subtiri de actina, troponina si tropomiozina. Filamentele subtiri au
diametru de 0,5 nm si lungime de 1m si se afla asezate intre filamentele de
miozina. La extremitatea lor dinspre centrul sarcomerului, nu se ating aflandu-se
la distanta una de alta.
Zona H este lipsita de filamente de actina. In mijlocul zonei intunecate apare
linia M datorita prezentei unei retele proteice care fixeaza partea centrala a
miozinei si totodata datorita ingrosarii miozinei in partea centrala. Linia M ar avea
41
rol similar linie Z. In procesul de aliniere a filamentelor mai participa 3 molecule
proteice.
TITINA este una dintre cele mai mari molecule proteice cunoscute; contine
250 000 de a.a. Este o proteina elastica ce se insera pe linia Z si linia M. Ea
confera elasticitate sarcomerului. Cand sarcomerul este intins, titina asigura
revenirea la pozitia initiala. Titina reprezinta 10% din proteinele musculare.
NEBULINA insoteste filamentele de actina si le mentine in pozitie. Este tot o
proteina cu molecula mare.
DISTROFINA se fixeaza pe glicoproteinele din membrana fibrei musculare si de
lamina retelei interstitiale, iar in interiorul fibrei musculare, de actina.Distrofina
este sintetizata de o gena situata pe cromozomul X. Ea formeaza un fel de
citoschelet in fibra musculara.
Daca este deficitara gena care codifica sinteza distrofinei, aceasta nu se mai
sintetizeaza si rezultatul este aparitia DISTROFIEI MUSCULARE DUCHENE, boala cu
transmitere ereditara genozomal recesiva ( o fac numai barbatii iar femeile o
transmit)
Aceasta boala se caracterizeaza prin inlocuirea fibrelor musculare cu fibre
conjunctive si individul moare prin insuficienta respiratorie, de obicei in jurul
varstei de 30 de ani cand sunt fibrozate fibrele musculare din aparatul respirator.
In sarcoplasma se mai gasesc organite nespecifice:
- Mitocondrii: sunt raspandite in masa musculara, jucand un rol important in
metabolismul oxidativ celular
- Reticulul endoplasmatic- denumit in acest caz reticul sarcoplasmatic, este alcatuit
din tuburi longitudinale care se termina cu niste saci sau cisterne.
Tuburile longitudinale invelesc (ca o perdea) miofilamentele dintr-un sarcomer. In zona
cisternelor se afla Ca. Intre 2 cisterne se intercaleaza un alt canal care strabate muschiul:
tubul in T. Tubul In T se desprinde din membrana si trece intre 2 saci. El comunica liber cu
exteriorul si formeaza impreuna cu cisternele o TRIADA. Triada in musculatura scheletica
este striata in dreptul zonei de delimitare dintre discul intunecat si discul clar.
42
Fiecare sarcomer contine 2 triade. Tubul in T, ce reprezinta o infundare a membranei,
transmite depolarizarea in interiorul fibrei.
In membrana tubului in T se gasesc canale de calciu dihidropiridin sensibile
(dihidropiridina este blocant al canalelor de Ca). Canalele dunt asezate cate 4 pe mai multe
siruri paralele. In reticulul sarcoplasmic, corespunzator tubului in T se gasesc canale rianodin
sensibile (fata in fata cu canalele de Ca) care se deschid sub influenta rianodinei (alcaliod
vegetal). Sub influenta alcanoidului, canalele raman deschise, Ca trece in sarcoplasma si
provoaca contractura musculara ( contractie de lunga durata).
In sarcoplasma exista numeroase enzime legate de mitocondrii sau libere care participa la
procesul oxidativ. Unele fibre muscular contin MIOGLOBINA proteina similara HB care
poate retine O2. Fibrele musculare contin mioglobina sunt rosii iar cele care nu contin sunt
albe.
Fibrele albe : se contracta rapid chiar si in conditii de ANAEROBIOZA.
Fibrele rosii: se contracta mai lent in conditii de AEROBIOZA.
Compozitia muschiului:
Apa:74-80%
Proteine: 16-20%
Lipide: 0,5-1 %
Glucide: 0,2-0,8%
Minerale: 1%
Apa: uscarea muschiului impiedica contractia. Inlocuind apa cu apa grea (D2O D - e
izotopul deuteriu al hidrogenului) muschiul nu se mai relaxeaza.
Proteinele : - contractile :60%
-necontractile: proteine din citoschelet 20% si enzime 20%
Lipidele si glucidele reprezinta sursa de energie. Mineralele sunt implicare in polarozarea si
depolarizarea membranei si in declansarea contractiei.
Proteinele contractile sunt : Miozina , actina, troponina si tropomiozina.
Miozina - molecula mare cu G
M
=480 000 Da. Sub influenta unor detergenti se descompune
in 2 lanturi polipeptidice cu greutate mare si 4 lanturi polipeptidice cu greutate mica. Lantul
polipeptidic este impletit sub forma de dublu helix.
43
La extremitatea lui se afla 2 formatiuni globuloase : capetele MIOZINEI.
Portiunile subtiri ale miozinei intra in structura filamentului gros de miozina iar portiunile
globuloase sunt proeminente in afara. Ele se extind in afara si datorita unei prelungiri a
dublului helox. Partea globuloasa si partea externa formeaza PUNTI TRANSVERSALE.
Portiunile subtiri din miozina au terminatii care se privesc fata in fata. Pe portiunile
globuloase se afla celelalte 4 polipeptide cu G
M
mai mica. Se pare ca sunt implicate in
controlul contractiei musculare. Intr-un miofilament exista 1500 fibrile de miozina si cca 300
filamente de actina, troponina, tropomiozina. Ele sunt asezate intr-o simetrie hexagonala,
fiecare filament de miozina fiind inconjurat de 6 filamente de ACTINA. Puntile transversale
sunt asezate de o parte si de alta a filamentelor de MIOZINA.
La o distanta de 14,3nm ele sufera o rotatie de 120
0
, se fixeaza in alta zona iar dupa 360
o
se
afla in pozitia initiala (este vorba despre partile tranversale)
Actina are o greutate moleculara 420 000 Da. Se afla sub forma globulara: actina G. In
filamentele de actina ele se prind cap la cap si formeaza ACTINA F (filamentoasa) care
contine cca 300 molecule de actina G. Actina F se afla impletita sub forma de dubu helix. In
santul delimitat de dublu helix se afla TROPOMIOZINA care acopera cam 6 molecule de
actina G.
La suprafata tropomiozinei se fixeaza TROPONINA, alcatuita din 3 fragmente:
44
Troponina I: legata de actina
Troponina T: legata de tropomiozina
Troponina C: capabila sa fixeze calciul
Troponina mentine tropomiozina intr-o pozitie care nu permite legarea puntilor
transversale de moleculele de ACTINA. Moleculele de actina au ... zona acceptoare pentru
puntile transversale (in repaus zonele acceptoare sunt mascate). Cand Ca se fixeaza pe
fragmentul C al troponinei, tropomiozina se scufunda in santul dintre filamentele de ACTINA
si sunt dezvelite 7 molecule de actina globulara -> 7 locuri de interactiune cu puntile
transversale ale miozinei. Fragmentul C al troponinei seamana cu calmodulina din
citoplasma si este capabil sa fixeze 4 ioni de Ca.
MECANISMUL CONTRACTIEI MUSCULARE
Este un mecaniscm molecular
1. Teoria GLISARII formulata de Huxley 1969. Se bazeaza pe constatarea facuta la
microscopul optic si anume ca in timpul contractiei musculare liniile Z se apropie de
discul intunecat si zonele clare pot sa dispara (dispare zona H)
O astfel de modificare nu se poate explica decat prin aceea ca miofilamentele de actina
aluneca printre filamentele (miofilamentele) de miozina pana in momentul in care menbrana
Z ajunge la extremitatea miofilamentelor de miozina. Dispare zona H. Filamentele de actina
pot chiar sa se intalneasca in contractiile puternice. La relaxare, zonele clare revin la valoarea
initiala, adica ele cresc in dimensiune si creste zona H.
Miscarea filamentelor de actina se datoreaza aderarii puntii transverse de filamentele
de actina, inclinarii lor spre centrul sarcomerului cu tractionarea actinei. Pe urma are loc
desprinderea de actina si inceperea unui nou ciclu.
45
Declansarea contractiei se face sub influenta depolarizarii membranei care transmite
prin tubul T la reticulul sarcoplasmic. In reticulul sarcoplasmic Ca este mai concentrat de cca
2000 de ori decat in sarcoplasma (10
-7
10
-8
moli/l) . Odata cu deschiderea canalelor
rianodinice Ca paraseste reticulul sarcoplasmic si concentratia de Ca urca la 10
-5
moli/l. Ca se
prinde de troponina si adanceste tropomiozina in santul dintre actina fibrilara. Se descopera
7 molecule de actina cu zonele acceptoare pentru MIOZINA (pentru puntea transversala )
Tropomiozina se adanceste in santul format de filamentele de actina.
Fiind inlaturata mascarea zonelor receptoare, puntea transversala a miozinei asezata
perpendicular pe filamentele de actina se prinde de zona acceptoare. Dupa prindere sufera
o inclinatie de la -> 30
o
la 45
o
si atrage dupa sine si lantul de actina .
Capetele globulare contin o zona actinica (de interactiune) cu actina si o fisura in care
se fixeaza ATP.
Miozina initiata in fisura descompune ATP-ul in ADP si fosfat. Desprinderea miozinei de
actina se face numai in prezenta ATP.
Din descompunerea ATF rezulta energie care readuce puntea transversala intr-o pozitie
perpendiculara pe actina. Din descompunere => ADP si fosfat anorganic care raman atasate
in fisura. Descompunerea ATP spune ca incarca puntile transversale (le aduce in pozitie
perpendiculara)
Incarcarea inseamna si depozitarea de energie in fisura. Puntea transversala se poate
prinde de o noua zona acceptoare a filamentelor de actina. Cu fiecare atasare si detasare a
puntilor transversale, muschiul se scurteaza su 1% din lungimea lui. Miscarea puntilor
transversale imita miscarea vaslelor.
Fara ATP puntea transv nu se incarca ( nu revine la pozitia perpendiculara)
RELAXAREA : are loc prin recaptarea Ca din sarcoplasma in reticulul sarcoplasmic,
Recaptarea se face prin mecanism de transport activ adica consumator de energie.
In membrana reticulului sarcoplasmic axista o ATP-aza Ca si Mg dependenta si ea
favorizeaza trecerea Ca in sarcoplasma. In sarcoplasma se descrie existenta unei proteine:
46
CALSECHESTRINA care poate fixa ionii de Ca . Nu numai recaptarea dar si eliberarea de Ca
din reticulul sarcopl. se face in afara transportului pasiv si prin transp activ: printr-o pompa
care necesita ATP.
ATP: este utilizat in contractie , la expulzarea si recaptarea Ca in reticulul
sarcoplasmic.
Pana cand Ca in sarcoplasma nu coboara la 10
-7
relaxarea nu are loc.
Daca nu exista ATP apare contractura musculara. Se observa dupa deces- rigiditatea
cadaverica instalata dupa 1-7 ore. Instalarea ei se face cu atat mai rapid cu cat ATP este mai
scazut in muschi. La 1 zi dupa moarte rigiditatea dispare prin interventia enzimelor
proteolitice care distrug proteinele musculare.
Rigiditatea: poate fi provocata de substante care mentin deschise canalele
rianodinice sau cofeina -> impiedica recaptarea Ca.
Contractura: poate sa apara si din cauza cresterii K in lichidul extracelular. K in exces
provoaca o depolarizare a fibrelor musculare. Dupa eforturi intense K iese din celula si det.
CONTRACTURA.
PROCESELE ENERGETICE CARE AU LOC LA NIVELUL FIBREI MUSCULARE (in timpul contractiei)
In timpul contractiei musculare energia se elibereaza din ATP. Cantitatea de ATP in
muschi este de 4 mol/kg si asigura 8 secuse musculare. ATP se reface rapid pe seama
creatinfosfatului (PC) = 20 mmol/kg
PC+ADPATP +P ( sub actiunea CPK)
P+energia rezultata din glicoza aeroba -> 39(sau whatever) molecule PC
P + energia rezultata din glicoliza anaeroba -> 4 molecule PC
Daca se blocheaza creatinfosfokinaza cu dinitrofenol muschiul se poate contracta de
50 de ori maxim 100, iar daca activitatea musculaturii produce conditii de anaerobioza : de
600 de ori.
Glicoliza aeroba confera muschiului posibilitatea de a se contracta de 10-20 000 de
or. Glicoliza anaeroba are multe neajunsuri in sensul ca favorizeaza acumularea acidului
lactic in muschi:
Glucoza + 2 ATP (sau glicogen +1 ATP) 2 acid lactic +4 ATP (prin anaerobioza)
Acidul lactic provoaca oboseala musculara se poate bloca enzimele din muschi.
Inceputul activitatii musculare se produce in conditii de anaerobioza (20-45 sec). Dupa 20-45
sec irigatia sangvina se adapteaza efortului fizic astfel incat fluxul sanvin se mareste de 20 de
ori.
In primele 1-2 minute muschiul contracteaza o DATORIE de O
2
astfel incat dupa
incetarea efortului consumul de O
2
nu coboara la 0 si ramane ridicat inca o perioada de timp.
Plata datoriei de O
2
:
Plata datoriei de oxigen este mai indelungata si mai mare decat datoria de la
inceputul efortului. Acest lucru se explica prin faptul ca dupa efort oxigenul este folosit
pentru :
47
-refacerea PC, ATP
-incarcarea cu oxigen a mioglobinei
-intretinerea unui ritm cardiac si respirator accelerat
-mentinerea energiei necesare in termoreglare (indepartarea caldurii dupa efort)
Eforturile de scurta durata (pana la 100 m/10s) folosesc energie eliberata in proportie
de 98% ??? din glicoliza anaeroba.
Eforturile de lunga durata (ex 5000 metri) folosesc energie care se elibereaza 5% din
aderarea glucidelor. Acizii grasi reprezinta un alt combustibil utilizat de muschi. Din acizii
grasi liberi se elibereaza 75-95% din energie in cursul efortului fiziologic.
PRODUCTIA DE CALDURA: -tesutul muscular transforma energia chimica in energie
mecanica si o parte din energie se degaja sub forma de caldura. Muschii convertesc energia
in lucru mecanic in proportie de 20-25%, maxim 50% - RANDAMENT MUSCULAR
Chiar si in repaus musculatura elibereaza 0,0002 kcal/g/muschi. In efort
productia de energie calorica creste. Degajarea caldurii se poate face :
-imediat
-tardiv
Caldura imediata se transforma in CALDURA ACTIVARE aparuta in momentul
stimularii si CALDURA DE DEGAJARE, INTRETINERE la contractia izometrica urmata de
caldura de relaxare.
CALDURA IMEDIATA : - de activare
-de intretinere
-de relaxare
In contractiile IZOTONICE in afara caldurii de intretinere si activare se mai adauga caldura de
scurtare
48
Caldura tardiva se explica prin reactiile biochimice care conduc la refacerea ATP, PC si rezulta
in special din mecanismele aerobe. In muschiul tinut in conditii de anaerobioza, muschiul nu
mai degaja caldura tardiva.
Reflexul
Reflex reactia constienta aparuta dupa aplicarea unui stimul pe un receptor senzitiv
(sau senzorial), reactie dirijata de S.N.
Cel mai simplu reflex: atingerea unei suprafete fierbinti determina retractia mainii
inainte de aparitia senzatiei dureroase.
Exista o multime de reflexe care se produc fara participare constienta: reflexe respiratorii,
digestive, vasculare, de motricitate, proprioceptive (intervin in mentinerea posturii).
Baza anatomica a reflexului este ARCUL REFLEX.
ARCUL REFLEX se compune din :
1. Receptor terminatie nervoasa libera(sau receptor corpuscular)
2. Calea aferenta fibre nervoase care au pericarionul situat in ganglionul spinal sau
in ganglionii atasati .....
3. Centru nervos situat in maduva spinarii sau in structurile nervoase suprajacente
(tr. cerebral).
4. Calea eferenta reprezentata de fibre somatice sau fibre vegetative. Face
legatura cu organul efector.
Organul efector poate fi:
49
- Muschi: transforma energia chimica in miscare
- Glande secretorii (exocrine)
Receptorii: clasificati de Sherrington in 1906 (dupa pozitia ocupata de receptor)
- Exteroceptori
- Interoceptori
Exteroreceptorii pot fi
-Telereceptori: sunt receptori la distanta (ex: vizuali, auditivi)
-De contact: termici, durerosi, tactili
Interoceptorii pot fi :
-Proprioceptori: prezenti in muschi, tendoane, articulatii
-Visceroceptori: organe interne
In functie de stimulul care actioneaza supra lor pot fi :
1. Mecanoreceptori (presoreceptori) : receptori tactili, baroreceptori (presiune) din
cadrul aparatului CV
2. Chemoreceptori: sunt stimulati de substante chimice ( ex: olfactivi, gustativi,
chemoreceptorii din aparatul CV stimulati de variatii ale concentratiei de oxigen si
concentratii mari de dioxid de carbon)
3. Termoreceptori: pentru cald, rece
4. Algoreceptori: sunt receptionati stimulii durerosi
5. Electromagnetici: sensibili la radiatiile electromagnetice (ex: receptorii vizuali din
retina)
Sinapsa
Este zona de legatura dintre 2 neuroni. A fost descrisa pentru prima dara de catre
Sherrington in 1897. Sin=impreuna, nopsis = a lega.
Sinapsele se pot prezenta sub forma de:
1. Terminatii libere
2. Sub forma ingrosata la extremitate (buton sunaptic)
3. Cupa
In functie de zona neuronala cu care terminatia nervoasa vine in contact :
1. Sinapse axo-dendritice
2. Sinapse axo-somatice
3. Sinapse axo-axonice
Structura electronomiscroscopica a sinapselor:
1. Buton sinaptic: contine o parte presinaptica si o parte postsinaptica. In zona
presinaptica exista organite celular. Microtubulii ajung pana in zona butonului
sinaptic (au rol de transport)
50
Inainte de butonul sinaptic, axonul isi pierde teaca de mielina. In interiorul butonului
sinaptic exista mitocondrii ( importante pentru metabolismul oxidativ din sinapsa).
Sinapsa contine o multime de vezicule:
- Clare : contin acetilcolina, glicina, GABA, glutamina
- Dense: contin catecolamine (noradrenalina, adrenalina, dopamina)
Veziculele se aduna in zona presinaptica. Veziculele clare au dimensiuni mici.
Sinapsele pot contine si granulatii de dimensiuni mari (sunt intunecate) ce
contin ca mediator sinaptic polipeptide: VIP.
Granulatiile mari, intunecate sunt mai indepartate de membrana presinaptica.
2. Fanta sinaptica (spatiu sinaptic sau fisura sinaptica)
Este situata intre terminatia nervoasa si dendrita cu care vine in contact (pentru
sinapsele axo-dendritice) sau intre terminatia nervoasa si axonul celuilalt neuron (axo-
axonice) sau organul efector, (sau corpul celular).
Fanta sinaptica are dimensiuni de 20-40nm. Pe un motoneuron din coarnele
anterioare sunt aprox. 10 000 sinapse (8000 conectate cu dendrite, 2000 cu corpul celular).
In scoarta cerebrala, peste 90% din sinapse se fac cu dendritele si 3-5% cu corpul
celular.
Membrana care vine in contact cu butonul membrana postsinaptica (poate fi o spira
dendritica). In zona postsinaptica nu exista vezicule si organite celulare.
Cele mai multe vezicule clare, mici contin ACO. ACO se formeaza in butonul sinaptic din
colina si radicalul ACETIL sub actiunea unei enzime : COLIN ACETILT RANSFERAZA.
Fiecare vezicula presinaptica contine 3000-10 000 molecule de ACO. Butonul este
inzestrat cu mediatori ce pot transmire circa 10 000 de impulsuri. Transmiterea se face prin
eliberarea mediatorului in fanta sinaptica. Mediatorul actioneaza asupra unui receptor
membranar pentru ACO. Receptorii membranei, activati de mediatori sau chiar de hormoni
au 2 portiuni:
- o portiune externa pe care se fixeaza substanta de actioneaza pe receptor (liganzi)
- o portiune care se prezinta ca un canal de ioni : portiunea IONOFORA a receptorului.
Dupa ce ACO se fixeaza pe receptor, se deschide un canal cu diametrul de 0,65 nm.
Canalul incarcat cu sarcini negative nu permite trecerea ionilor negativi ( de ex. Cl) , dar pot
trece Na, Ca, K. Gradientul cel mai mare de concentratie il are Na (diferenta intre
concentratia extra si intracelulara) astfel incat Na navaleste in celula.
51
Efectele ACO asupra membranei postsinaptice sunt de scurta durata.
Intre zona pre si postsinaptica exista o retea de glicoproteine pe care este fixata o
enzima: COLINESTERAZA care descompune ACO. Timpul de actiune al ACO este de aprox
8ms. Unda de depolarizare care ajunge in zona sinaptica, deschide canalele de Ca
2+
din
membrana presinaptica si Ca
2+
navaleste in terminatii. Veziculele cu mediator sunt fixate in
citoscheletul terminatiei nervoase.
Ca
2+
se leaga de o proteina CALMODULINA care activeaza PROTEINKINAZA care
activeaza la randul sau SINAPSINA I care favorizeaza desprinderea veziculelor de pe
citoschelet si ancorarea lor la membrana presinaptica.
Eliberarea mediatorului se face prin exocitoza, proces la care participa 8 proteine.
In peretele veziculei se afla SINAPTOBREVINA
Prin interventia celor 8 proteine descrise se produce un por in membrana
presinaptica ce lasa sa treaca mediatorul in fisura sinaptica --> SINAPTOFORINA. Veziculele
ajung in zona butonului cu transportul activ axoplasmatic (de-a lungul microtubilor). Dupa ce
isi varsa continutul, veziculele sunt reciclate (veziculele sunt acoperite de CLATRINA.).
Sub actiunea DINAMINEI (o GTP-aza) se formeaza un inel in jurul gatului veziculei
golite care favorizeaza desprinderea veziculei ce se ataseaza endosomilor. Se desprinde din
endosomi, dupa care se incarca in terminatia nervoasa cu mediatori (endocitoza).
52
O data cu penetrarea unui ion ce Ca in zona presinaptica se produce mobilizarea a
300 de vezicule. Dupa actiunea mediatorului pe membrana presinaptica, in zona
postsinaptica se produce un proces de depolarizare sau hiperpolarizare.
Raspunsul postsinaptic nu poate fi declansat de activarea unei singure sinapse (
hiperpolarizare =inhibitie, depolarizare = excitatie ). Pentru a aparea acest raspuns trebuie sa
se descarce cel putin 10% din sinapse. Fenomenul se numeste SUMARE SPATIALA.
Exista insa posibilitatea ca una din sinapse sa descarce impulsuri repetitiv si sa seclanseze
raspunsul. In asemenea situatii vorbim de SUMARE TEMPORALA.
In zona postsinaptica poate sa apara o depolarizare: potential postsinaptic excitator
(PPSE). Acesta apare dupa o latenta de 0,5 ms. Atinge apogeul dupa 1-1,5 ms. Ajunge la
valoarea normala dupa 15 ms. PPSE se aseamana cu potentialul generator.
Cu cat frecventa impulsurilor este mai mare cu atat valoarea PPSE este mai mare. PPSE
poate declansa un potential de actiune propagat cand atinge valoarea de -59 mv.
Depolarizarea de 11mv ( -70+11=-59) declanseaza petential propagat (executatia) numai
cand sinapsa se afla localizata in partea initiala a axonului, care pe o distanta de 100m este
amielinica.
Pentru ca PPSE sa determine excitatia in celelalte zone (dendtite, corp celular),
trebuie ca diferenta de potential sa fie de 35-30 mv. Excitarea porneste tot de la portiunea
initiala a axonului unde exista densitatea cea mai mare de canale de Na
+
.
Anumiti mediatori ca acidul Gama animobutiric (GABA) sau glicina, actionand pe
zona postsiinaptica produc PPSI (post pot. sinaptic inhibitor) = imaginea in oglinda a PPSE.
PPSI are o latenta de 0,5ms, membrana se hiperpolarizeaza, potentialul de repaus
negativandu-se cu -3, -5 sau chiar -10 mv. (ajunge pana la -80 mv)
53
Aparitia PPSI se datoreste deschiderii unor canale de Cl
-
=> Cl
-
penetreaza in celula, si
a unor canale de K
+
avand drept consecinta iesirea K
+
din celula.
Inhibitia provocata de PPSI poarta numele de inhibitie directa sau postsinaptica.Se
cunoaste si o inhibitie presinaptica sau indirecta in care neuronii inhibitori activati nu
modifica potentialul transmembranar ci reduc amplitudinea PPSE si cantitatea de mediator
sinaptic eliberata de butonul sinaptic. Inhibitia presinaptica poate dura pana la 10-200-300
ms fata de cea postinaptica care persista 15-30 ms.
Ex:
INHIBITIA DIRECTA reflexele motorii medulare activeaza un grup de muschi agonisti si
inhiba un grup de muschi antagonisti. Inhibarea musculaturii antagoniste se datoreaza
prezentei in maduva a unor celule apoase cu o terminatie scurta (celule Golgi) care se
termina pe motoneuronii muschilor antagonisti pe care-i hiperpolarizeaza prin aliberare de
glicina.
PPSE: in neuronii care comanda muschii agonisti
PPSI: ia nastere in neuronii care comanda m. antagonisti.
INHIBITIA INDIRECTA: este realizata prin eliberare de GABA. GABA produce inhibitia
actionand pe receptori:
1. Receptori GABA A=> deschiderea canalelor de Cl
-
cu intrarea Cl
-
in celula
2. Receptori GABA B => deschiderea canalelor de K
+
cu iesirea K
+
din celula
Semnalele ajung in zona coarnelor posterioare si substanta gelationoasa unde fac sinapsa cu
cel de-al 2-lea neuron si se continua cu tractul spino-talamic. Fibrele senzitive somatice care
pornesc de la proprioceptori sunt colaterale in zona presinaptica (fasciculele GOLL si
BURDACH). Astfel executarea electrica a fibrelor radiculare care intra in constitutia
fasciculelor GOLL si BURDACH poate deseori suprima durerea.
Particularitatile transmiterii sinaptice
1. Conducerea impulsurilor prin sinapsa se face intr-o singura directie: din zona
presinaptica in zona postsinaptica (deoarece veziculele se afla numai in zona
presinaptica iar receptorii numai in zona postsinaptica).
2. Transmiterea sinaptica sufera o intarziere de cca 0,5 ms la fiecare sinapsa. Daca un
reflex are o latenta cu 0,5 ms mai mult decat altul inseamna ca in arcul sau reflex
exista o sinapsa in plus.
3. Oboseala sinaptica: cand o sinapsa este stimulata repetitiv un timp mai indelungat,
activitatea eu se reduce progresiv pana la rarirea si disparitia potentialelor de actiune
propagate. Oboseala se explica prin epuizarea mediatorilor din zona presinaptica sau
54
blocarea receptorilor din zona postsinaptica. Oboseala are uneori efecte benefice: in
criza epileptica unde sunt contractii musculare puternice.
4. Sensibilitatea sinapsei la hipoxie. Pentru sinteza de ATP (energie) necesar in
transmiterea impulsurilor sinaptice este necesara degradarea oxidatica a glucozei. In
hipoxie, ATP nu se reface. Astfel intreruperea circulatiei cerebrale pt 10 secunde
determina disparitia starii de veghe.
5. Sinapsele pot fi sensibile la anestezice, narcotice. Fiecare sinapsa se afla in
comunicare libera cu lichidul interstitial. Anestezicele actioneaza fie blocand
eliberarea de Ca
2+
, fie inhiba producerea de mediatori.
6. Facilitarea posttetanica. Stimularea repetitiva a unui muschi => tetanos. Acesta apare
dupa o stimulare repetitiva a nervului urmata de o intrerupere ptr o scurta perioada
de timp. Daca se revine cu un stimul, reactia de raspuns la stimul este mult mai mare.
Explicatia : impulsurile repetitive au determinat in zona presinaptica un nr mare de
moelcule de Ca
+2
sa se concentreze in terminatiile presinaptice din cauza ca pompa
de Ca
+2
evacueaza ionii prea incet.
Datorita faptului ca facilitarea posttetanica dureaza de la cateva secunde la cateva
ore, in functie de neuron, ar putea sta la baza memoriei de scurta durata.
--------------------------------------- ------------------------------------------------------------------
Corpusculului Vater- Pacini, la compresiunea directa a terminatiei nervoase are loc o
adaptare latenta a receptorului, adica receptorul fazic devine tonic.
b) prin procesul de acomodare a fibrei nervoase senzitive la potentialul generator.
Ex: de R. Fazici : r tactili, r olfactivi
R. Tonici: algoreceptorii, receptorii pt frig, baroreceptorii, fusurile
neuromusculare.
Acesti receptori au rol in protejarea organismului: senzatiile de durere si de reac sunt
declansate de stimuli cu potential nociv. Daca algoreceptorii si receptorii pentru frig
s-ar adapta rapid, si-ar pierde roul fiziologic de a dezvalui pericolul. Baroreceptorii
sinocarotidieni si cardioaortici intervin in permanenta in reglarea presiunii arteriale
iar adaptarea lor ar limita precizia cu care opereaza sistemul de reglare.
S-ar putea să vă placă și
- Fiziologie LP NR 6 Anul IDocument17 paginiFiziologie LP NR 6 Anul IAnna MariaÎncă nu există evaluări
- Toate Glandele Secretorii Ale Tractului Gastrointestinal Deservesc 2 Functii PrimareDocument5 paginiToate Glandele Secretorii Ale Tractului Gastrointestinal Deservesc 2 Functii PrimareRoxana EnachescuÎncă nu există evaluări
- Secretia GastricaDocument6 paginiSecretia GastricaElena MirelaÎncă nu există evaluări
- Ingrijrea Pacientului Cu Ulcer Gastro Duodenal PerforatDocument80 paginiIngrijrea Pacientului Cu Ulcer Gastro Duodenal PerforatPatrut Florentina100% (1)
- Receptorii Celulei Parietale Popescu Dragos PaulDocument6 paginiReceptorii Celulei Parietale Popescu Dragos PaulPopescu DragosÎncă nu există evaluări
- Secretia de Acid GastricDocument9 paginiSecretia de Acid GastricIrina ItuÎncă nu există evaluări
- Fiziologia Animalelor Si Omului - Digestiv Curs 6Document10 paginiFiziologia Animalelor Si Omului - Digestiv Curs 6ionutÎncă nu există evaluări
- Sucul GastricDocument10 paginiSucul GastricVoropanov Ion AlexandruÎncă nu există evaluări
- Tema 4.4 Fiziologie STOM ROMDocument4 paginiTema 4.4 Fiziologie STOM ROMstudentÎncă nu există evaluări
- Functia Motorie A StomaculuiDocument8 paginiFunctia Motorie A Stomaculuisvlaic2006601Încă nu există evaluări
- Tema 2 DigestivDocument10 paginiTema 2 DigestivMihaela Botnari-GuţuÎncă nu există evaluări
- Secretia GastricaDocument4 paginiSecretia GastricaOana PauliucÎncă nu există evaluări
- Sfincterul Piloric Si Evacuarea GastricaDocument21 paginiSfincterul Piloric Si Evacuarea Gastricasweet_d3ea100% (1)
- Reglarea Secretiei GastriceDocument11 paginiReglarea Secretiei GastriceAndrei TîrziuÎncă nu există evaluări
- Fazele Secretiei GastriceDocument16 paginiFazele Secretiei GastriceIoan-Radu StănculescuÎncă nu există evaluări
- Anul 2 Capitolul 5 Tema 5 Secrețiația AbsorbțiaDocument19 paginiAnul 2 Capitolul 5 Tema 5 Secrețiația AbsorbțiaDaniela DucantoniÎncă nu există evaluări
- Ulcerul 2020Document90 paginiUlcerul 2020CoblişPaulÎncă nu există evaluări
- PANCREASDocument8 paginiPANCREASgaftea_alexÎncă nu există evaluări
- Ingrijrea Pacientului Cu Ulcer Gastro-Duodenal PerforatDocument80 paginiIngrijrea Pacientului Cu Ulcer Gastro-Duodenal PerforatSchuldesz Paul100% (2)
- Reglarea Secretiei GastriceDocument29 paginiReglarea Secretiei GastriceConstantin George AdrianÎncă nu există evaluări
- Diagnosticarea Sindromului Zollinger-EllisonDocument10 paginiDiagnosticarea Sindromului Zollinger-EllisonAdrian BlidarÎncă nu există evaluări
- 05 Patologia Chirurgicala A Stomacului Si Duodenului PDFDocument168 pagini05 Patologia Chirurgicala A Stomacului Si Duodenului PDFAndreea ElenaÎncă nu există evaluări
- Medicamente Utilizate in Terapia Afectiunilor Aparatului DigestivDocument34 paginiMedicamente Utilizate in Terapia Afectiunilor Aparatului DigestivciobineluÎncă nu există evaluări
- Bariera Mucoasei GastriceDocument18 paginiBariera Mucoasei Gastriceemonil26100% (1)
- Fiziologia Sistemului DigestivDocument5 paginiFiziologia Sistemului DigestivlauraÎncă nu există evaluări
- 8ingrijrea Pacientului Cu Ulcer Gastro-Duodenal PerforatDocument80 pagini8ingrijrea Pacientului Cu Ulcer Gastro-Duodenal PerforatSchuldesz Paul100% (2)
- Digestia IntestinalaDocument5 paginiDigestia IntestinalaCosmin Calancia0% (1)
- Tulburările Motilităţii Şi Secreţiei Tractului DigestivDocument5 paginiTulburările Motilităţii Şi Secreţiei Tractului Digestivnicole_22Încă nu există evaluări
- Pancreasul ExocrinDocument15 paginiPancreasul Exocrinjust_me18100% (4)
- Pancreatita AcutaDocument40 paginiPancreatita AcutaAna-Maria Chiribau86% (7)
- 05 Patologia Chirurgicala A Stomacului Si Duodenului CU NOTITEDocument169 pagini05 Patologia Chirurgicala A Stomacului Si Duodenului CU NOTITEMicky MihalacheÎncă nu există evaluări
- Reglarea Secretiei GastriceDocument24 paginiReglarea Secretiei GastriceoanacamelliaÎncă nu există evaluări
- Medicamente Utilizate in Terapia Afectiunilor Aparatului DigestivDocument34 paginiMedicamente Utilizate in Terapia Afectiunilor Aparatului DigestivAlina PetreÎncă nu există evaluări
- Absorbţia În Tubul DigestivDocument18 paginiAbsorbţia În Tubul DigestivNicolaiChistolÎncă nu există evaluări
- Celulele ParietaleDocument26 paginiCelulele ParietaleAlexandra BÎncă nu există evaluări
- Reglarea Secretiei PancreasuluiDocument11 paginiReglarea Secretiei PancreasuluiAdelina MkyÎncă nu există evaluări
- Pansamente GastriceDocument40 paginiPansamente GastriceMari Clim100% (1)
- WWW - Referate.ro-Medicatia Antiulceroasa 689fcDocument77 paginiWWW - Referate.ro-Medicatia Antiulceroasa 689fcIonela Hagiu100% (1)
- Cap3 DigestivDocument73 paginiCap3 DigestivTripimeebÎncă nu există evaluări
- Splanhnic, Pancreas, EsofagDocument6 paginiSplanhnic, Pancreas, EsofagEllena CristianaÎncă nu există evaluări
- Fiziologie Curs 2Document13 paginiFiziologie Curs 2Radu TeodoruÎncă nu există evaluări
- Prezentare Digestia 1Document153 paginiPrezentare Digestia 1Tatiana ManoilaÎncă nu există evaluări
- Esofagita - Lucrare de DiplomaDocument34 paginiEsofagita - Lucrare de DiplomaLiliana50% (4)
- Histamina (1) .Agonisti Si Antagonisti Ai Receptorilor HistaminiciDocument3 paginiHistamina (1) .Agonisti Si Antagonisti Ai Receptorilor HistaminiciValerica Zarnescu CreangaÎncă nu există evaluări
- Secretia Pancreatica ExocrinaDocument21 paginiSecretia Pancreatica ExocrinaNarcisel Patratel100% (2)
- Digestiv Curs 1Document43 paginiDigestiv Curs 1Alina Alexandra ToeaÎncă nu există evaluări
- Afecţiunile Pancreasului Şi StomaculuiDocument9 paginiAfecţiunile Pancreasului Şi StomaculuiDidiÎncă nu există evaluări
- BRGE Esofag Barrett.Document38 paginiBRGE Esofag Barrett.Veronica ZubcoÎncă nu există evaluări
- 4-5determinismul Travaliului - DistociiDocument10 pagini4-5determinismul Travaliului - DistociiClaudia PenaruÎncă nu există evaluări
- PancreasulDocument13 paginiPancreasulandreyadÎncă nu există evaluări
- Fiziologie Curs 5Document6 paginiFiziologie Curs 5buburuzeleÎncă nu există evaluări
- 30 Fiziologia Sistemului DigestivDocument6 pagini30 Fiziologia Sistemului Digestivcosmin pesteÎncă nu există evaluări
- DigestiaDocument9 paginiDigestiaAlexandru Rusen100% (1)
- AcetilcisteinaDocument13 paginiAcetilcisteinaelish3baÎncă nu există evaluări
- Superintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateDe la EverandSuperintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateÎncă nu există evaluări
- Dieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivDe la EverandDieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivEvaluare: 5 din 5 stele5/5 (1)
- KetoFast: combină puterea postului intermitent cu dieta ketogenicăDe la EverandKetoFast: combină puterea postului intermitent cu dieta ketogenicăEvaluare: 4 din 5 stele4/5 (1)
- Rețete vegetariene din toată lumea. Mâncăruri sănătoase și gustoaseDe la EverandRețete vegetariene din toată lumea. Mâncăruri sănătoase și gustoaseEvaluare: 4 din 5 stele4/5 (8)
- Semio LogieDocument154 paginiSemio LogiebucureanionutÎncă nu există evaluări
- Far Maco LogieDocument24 paginiFar Maco LogiebucureanionutÎncă nu există evaluări
- Cursuri Biocel 2Document95 paginiCursuri Biocel 2bucureanionut100% (1)
- Curs Biochimie Sem IIDocument124 paginiCurs Biochimie Sem IIbucureanionut100% (3)
- BiocelDocument50 paginiBiocelMariana Sorică100% (1)