S-ar putea să vă placă și
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Fenomene de TransportDocument116 paginiFenomene de TransportNoțingher RaduÎncă nu există evaluări
- CELULADocument10 paginiCELULAIoana-Camelia OlteanuÎncă nu există evaluări
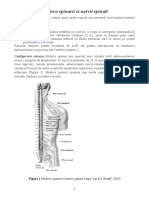
- Maduva Spinarii Si Nervii SpinaliDocument9 paginiMaduva Spinarii Si Nervii SpinalinetprintÎncă nu există evaluări
- Celula BacDocument19 paginiCelula Bacalina lascutÎncă nu există evaluări
- Biocel LP 9-Mitoza PDFDocument30 paginiBiocel LP 9-Mitoza PDFOana RaduÎncă nu există evaluări
- Organizarea Generala A CelulelorDocument7 paginiOrganizarea Generala A CelulelorDragos IdkÎncă nu există evaluări
- Anatomie - Curs1 - CelulaDocument27 paginiAnatomie - Curs1 - Celulaalfa5017Încă nu există evaluări
- MitozaDocument3 paginiMitozaAlexandra MiocÎncă nu există evaluări
- Teoria Cromozomială A EreditățiiDocument19 paginiTeoria Cromozomială A EreditățiiAlexia PăpădieÎncă nu există evaluări
- Diviziunea CelularaDocument3 paginiDiviziunea CelularaValentinoÎncă nu există evaluări
- CelulaDocument8 paginiCelulaAnca Elena SavuÎncă nu există evaluări
- Curs 5,6,7,8 EmbriologieDocument18 paginiCurs 5,6,7,8 EmbriologieAlexandru GiubegaÎncă nu există evaluări
- Celula ProcariotaDocument10 paginiCelula ProcariotaAndreea VirtejÎncă nu există evaluări
- Călătorie În Interiorul Unei Celule EucarioteDocument6 paginiCălătorie În Interiorul Unei Celule EucarioteAleee 06100% (1)
- Celula BacterianaDocument4 paginiCelula BacterianaSss Vvv100% (1)
- Celula EucariotaDocument15 paginiCelula Eucariotaiancu alexandru100% (2)
- Biologe LX - X.Document39 paginiBiologe LX - X.Alexandra IrimiaÎncă nu există evaluări
- Botanica Notiuni IntroductiveDocument3 paginiBotanica Notiuni IntroductiveMinciuna Alina100% (1)
- Celula. ComponenteDocument32 paginiCelula. ComponenteAnca Dănăilă0% (1)
- Membrana CelularaDocument9 paginiMembrana CelularaRadu MaziluÎncă nu există evaluări
- CitoscheletulDocument6 paginiCitoscheletulJessica Smith100% (1)
- ProtozoareDocument2 paginiProtozoarenicu159Încă nu există evaluări
- Laborator HistologieDocument3 paginiLaborator HistologieMatei BenoneÎncă nu există evaluări
- L 2 Celula Structura ProprietatiDocument10 paginiL 2 Celula Structura ProprietatirahatnikitaÎncă nu există evaluări
- Celula UmanaDocument14 paginiCelula Umanapaulv123100% (1)
- Capitolul I CELULA SI TESUTURIDocument14 paginiCapitolul I CELULA SI TESUTURIVioleta DiaconuÎncă nu există evaluări
- Cito PlasmaDocument19 paginiCito PlasmaAlexandra Vidz100% (1)
- Celula Eucariotă. Citoplasma Și Organitele CelulareDocument4 paginiCelula Eucariotă. Citoplasma Și Organitele CelulareLeul GamerÎncă nu există evaluări
- Evaluarea CorinaDocument4 paginiEvaluarea Corinaconstantin100% (2)
- BD 2Document89 paginiBD 2Ion MariusÎncă nu există evaluări
- Examen Biologie CelularaDocument8 paginiExamen Biologie CelularaElena IscuÎncă nu există evaluări
- CelulaDocument5 paginiCelulaProfu' de biologieÎncă nu există evaluări
- LP - OSMOZA Cu Calcul Pres OsmoticaDocument7 paginiLP - OSMOZA Cu Calcul Pres OsmoticaDarth Raz-WanÎncă nu există evaluări
- Diviziunea CelularaDocument4 paginiDiviziunea CelularaAle XaÎncă nu există evaluări
- CelulaDocument40 paginiCelulaSorin HoteaÎncă nu există evaluări
- Tesutul EpitelialDocument10 paginiTesutul EpitelialCorina Oprea-BorceaÎncă nu există evaluări
- Celula AdmitereDocument20 paginiCelula AdmitereBerta CopertaÎncă nu există evaluări
- Tema - ,,celula - Unitate Structural - Funcțională A Organismului UmanDocument12 paginiTema - ,,celula - Unitate Structural - Funcțională A Organismului UmanIon MosnoiÎncă nu există evaluări
- Transportul MembranarDocument8 paginiTransportul MembranarCojocariu ClaudiuÎncă nu există evaluări
- Biochimie Curs 3, 4, 5, 6Document16 paginiBiochimie Curs 3, 4, 5, 6Andreea MoraruÎncă nu există evaluări
- CelulaDocument12 paginiCelulaIustinian LesinschiÎncă nu există evaluări
- Matrice ExtracelularaDocument27 paginiMatrice ExtracelularaOana PatriciaÎncă nu există evaluări
- Studiul Comparativ A Celulei Procariote Si EucarioteDocument11 paginiStudiul Comparativ A Celulei Procariote Si EucarioteAndreea BarbuÎncă nu există evaluări
- Patologie Celulara - 2 - Celula - Organizare Si FunctiiDocument24 paginiPatologie Celulara - 2 - Celula - Organizare Si FunctiiSimina AnamariaÎncă nu există evaluări
- Membrana CelularaDocument6 paginiMembrana Celularaveronica.99Încă nu există evaluări
- Anatomie Comparata StudentiDocument42 paginiAnatomie Comparata StudentibiancaÎncă nu există evaluări
- Celula Diviziunea CelularaDocument26 paginiCelula Diviziunea CelularaGabØrÎncă nu există evaluări
- Imunitatea InnascutaDocument30 paginiImunitatea InnascutaKfa Dkh100% (1)
- Anatomie EmbriologieDocument50 paginiAnatomie EmbriologieSiera Gierone100% (1)
- CelulaDocument10 paginiCelulaValentina Gina MuaÎncă nu există evaluări
- CELULADocument29 paginiCELULAMadalina AndreeaÎncă nu există evaluări
- Referat Membrana CelularaDocument15 paginiReferat Membrana CelularaRazvan_GabrielÎncă nu există evaluări
- Proteinele Si Lipidele MembranareDocument13 paginiProteinele Si Lipidele MembranareFlorentina Claudia CalusaruÎncă nu există evaluări
- Structura Membranei CelulareDocument11 paginiStructura Membranei CelulareOana Palade100% (2)
- Celula EucariotaDocument5 paginiCelula EucariotaStefan GhergheÎncă nu există evaluări
- Membrana CelularăDocument3 paginiMembrana CelularăRobert201Încă nu există evaluări
- BiocelDocument24 paginiBiocelficioruÎncă nu există evaluări
- T5,6,7. CelulaDocument16 paginiT5,6,7. CelulaIuliana IrinaÎncă nu există evaluări
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
- Fisa de Activitate Drumeție Pe Malul JiuluiDocument1 paginăFisa de Activitate Drumeție Pe Malul JiuluiFlorentina Claudia CalusaruÎncă nu există evaluări
- Mari Compozitori Ai LumiiDocument1 paginăMari Compozitori Ai LumiiFlorentina Claudia CalusaruÎncă nu există evaluări
- Fisa de Activitate - Lumea Pe Care o DescoperiDocument1 paginăFisa de Activitate - Lumea Pe Care o DescoperiFlorentina Claudia Calusaru100% (1)
- Activ ExtracurriculareDocument2 paginiActiv ExtracurriculareFlorentina Claudia CalusaruÎncă nu există evaluări
- Grafic Școala Altfel 2023Document3 paginiGrafic Școala Altfel 2023Florentina Claudia CalusaruÎncă nu există evaluări
- Planif. 6 Bio 2022-2023Document6 paginiPlanif. 6 Bio 2022-2023Florentina Claudia CalusaruÎncă nu există evaluări
- Fisa - de - Activitate - Extrascolara+raport - Acoala Altfel 2023Document3 paginiFisa - de - Activitate - Extrascolara+raport - Acoala Altfel 2023Florentina Claudia Calusaru100% (1)
- Respiratia Aeroba Si AnaerobaDocument2 paginiRespiratia Aeroba Si AnaerobaFlorentina Claudia CalusaruÎncă nu există evaluări
- Programul Eco-Școala 2019-2020Document19 paginiProgramul Eco-Școala 2019-2020Florentina Claudia CalusaruÎncă nu există evaluări
- Consiliere Si Dezvoltare PersonalaDocument5 paginiConsiliere Si Dezvoltare PersonalaFlorentina Claudia Calusaru100% (1)
- Planificare Anuală V BioDocument1 paginăPlanificare Anuală V BioFlorentina Claudia CalusaruÎncă nu există evaluări
- Planificare - Anuala Cls. A 6-ADocument1 paginăPlanificare - Anuala Cls. A 6-AFlorentina Claudia CalusaruÎncă nu există evaluări
- Apa Aer SolDocument5 paginiApa Aer SolFlorentina Claudia CalusaruÎncă nu există evaluări
- Test Sumativ Biologie Clasa Av-ADocument2 paginiTest Sumativ Biologie Clasa Av-AFlorentina Claudia Calusaru100% (3)
- FIȘĂ DE RECAPITULARE Biologie VIIDocument2 paginiFIȘĂ DE RECAPITULARE Biologie VIIFlorentina Claudia CalusaruÎncă nu există evaluări
- pLANIFICARE BIOLOGIE VIIDocument7 paginipLANIFICARE BIOLOGIE VIIFlorentina Claudia Calusaru100% (1)
- 3.fise de Lucru Clasa 8 Suspendarea Cursurilor2Document8 pagini3.fise de Lucru Clasa 8 Suspendarea Cursurilor2Florentina Claudia CalusaruÎncă nu există evaluări
- Planificare Biologie VIII Semestrul - 1Document2 paginiPlanificare Biologie VIII Semestrul - 1Florentina Claudia CalusaruÎncă nu există evaluări
- Lectie Online Biotop. Biocenoza - Ecosistem.Călușaru FlorentinaDocument5 paginiLectie Online Biotop. Biocenoza - Ecosistem.Călușaru FlorentinaFlorentina Claudia Calusaru100% (1)
- Înmulțirea Prin SporiDocument2 paginiÎnmulțirea Prin SporiFlorentina Claudia Calusaru100% (2)
- Proiect de Lecție CreativDocument3 paginiProiect de Lecție CreativFlorentina Claudia CalusaruÎncă nu există evaluări
- Test Factorii EvoluțieiDocument1 paginăTest Factorii EvoluțieiFlorentina Claudia CalusaruÎncă nu există evaluări
- Verificare Mediul Intern Si Circulatia La MamifereDocument2 paginiVerificare Mediul Intern Si Circulatia La MamifereFlorentina Claudia CalusaruÎncă nu există evaluări
- ANIMALE VERTEBRATE Bio VDocument2 paginiANIMALE VERTEBRATE Bio VFlorentina Claudia CalusaruÎncă nu există evaluări
- Înmultirea VegetativăDocument4 paginiÎnmultirea VegetativăFlorentina Claudia CalusaruÎncă nu există evaluări