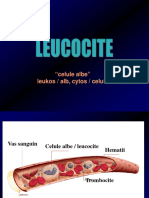
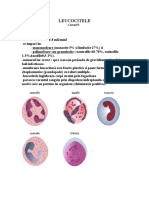
S-ar putea să vă placă și
- HEMATO Examen An 2Document81 paginiHEMATO Examen An 2Marina MalesÎncă nu există evaluări
- EritropoezaDocument7 paginiEritropoezasvlaic2006601Încă nu există evaluări
- Dieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivDe la EverandDieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivEvaluare: 5 din 5 stele5/5 (1)
- 56 Factori de Crestere HematopoieticiDocument6 pagini56 Factori de Crestere HematopoieticiRoxana ClimescuÎncă nu există evaluări
- Factori Crestere Biologie Cel PDFDocument13 paginiFactori Crestere Biologie Cel PDFRoxana ElenaÎncă nu există evaluări
- Hematopoieza 2010Document61 paginiHematopoieza 2010Oana AldeaÎncă nu există evaluări
- HematopoezaDocument4 paginiHematopoezaBogdan DianaÎncă nu există evaluări
- Curs 5 Hematopoieza Grupe SangeDocument81 paginiCurs 5 Hematopoieza Grupe SangeMischa Vlăsceanu100% (1)
- Hematologie Clinica Curs Dat La Farmacie An IIDocument129 paginiHematologie Clinica Curs Dat La Farmacie An IIManuela StefuÎncă nu există evaluări
- Cursul IvDocument15 paginiCursul IvMarianÎncă nu există evaluări
- Curs 2 Celulele Si Tesuturile Sist. ImunDocument7 paginiCurs 2 Celulele Si Tesuturile Sist. ImunMiruna-Elena NazâruÎncă nu există evaluări
- CURS 3 SÂNGE EritropoiezaDocument8 paginiCURS 3 SÂNGE EritropoiezaAlexandraFlorentynaÎncă nu există evaluări
- Curs 4 PDFDocument55 paginiCurs 4 PDFladyddyÎncă nu există evaluări
- Tesutul Sanguin - CursDocument7 paginiTesutul Sanguin - CursIoana campeanÎncă nu există evaluări
- Hematologie MG V Hematopoieza Si Anemii PT Test Martie 2013Document34 paginiHematologie MG V Hematopoieza Si Anemii PT Test Martie 2013Carmen JurcoaneÎncă nu există evaluări
- 5.fiziologia Seriei EritrocitareDocument4 pagini5.fiziologia Seriei EritrocitareOprea MariusÎncă nu există evaluări
- Hematologie FarmacologieDocument92 paginiHematologie Farmacologiemagdalena nita vladÎncă nu există evaluări
- Histologie Curs 9Document7 paginiHistologie Curs 9itismeroseÎncă nu există evaluări
- Hematologie ClinicaDocument56 paginiHematologie ClinicaCornelia SescuÎncă nu există evaluări
- C1 - HematopoiezaDocument50 paginiC1 - HematopoiezaMarta DumitracheÎncă nu există evaluări
- EritropoiezaDocument123 paginiEritropoiezavictorspanciocÎncă nu există evaluări
- HematopoezaDocument103 paginiHematopoezaDaniela CalinÎncă nu există evaluări
- Cursul IvDocument9 paginiCursul Ivantonio20002001Încă nu există evaluări
- Clase de Gene Implicate in Dezvoltarea CanceruluiDocument9 paginiClase de Gene Implicate in Dezvoltarea CanceruluiMihaela Țarnă100% (1)
- Cardiomicitele Si Celulele Stem Pentru RegenerareDocument43 paginiCardiomicitele Si Celulele Stem Pentru RegenerareViorel BudacÎncă nu există evaluări
- 2.b. Curs 2 Imunologie 2021 (Hematopoieza)Document67 pagini2.b. Curs 2 Imunologie 2021 (Hematopoieza)Cristian StanÎncă nu există evaluări
- Curs 6Document7 paginiCurs 6PopescuÎncă nu există evaluări
- 6 HematopoezaDocument8 pagini6 HematopoezaValentin MoisăÎncă nu există evaluări
- FizpatologieDocument50 paginiFizpatologieTaniușa SpataruÎncă nu există evaluări
- Fiziologia EritrocitelorDocument7 paginiFiziologia EritrocitelorSandra PopaÎncă nu există evaluări
- HematopoiezaDocument14 paginiHematopoiezaTeona OanaÎncă nu există evaluări
- HematopoiezaDocument6 paginiHematopoiezaIuliana BicecÎncă nu există evaluări
- HematopoiezaDocument19 paginiHematopoiezaElly ElizaÎncă nu există evaluări
- Leuco Cite LeDocument5 paginiLeuco Cite LeciprianÎncă nu există evaluări
- Hematologie Clinica StudentiDocument57 paginiHematologie Clinica StudentiNicky_Coravu100% (1)
- CITOKINEDocument11 paginiCITOKINENdbebe BebeÎncă nu există evaluări
- TROMBOCITELEDocument8 paginiTROMBOCITELEAndrada OargăÎncă nu există evaluări
- Medicatia SangeluiDocument2 paginiMedicatia SangeluiBogdan Adrian AgrigoroaieÎncă nu există evaluări
- C6 Eritropoieza Propr Morfofunc EritrDocument25 paginiC6 Eritropoieza Propr Morfofunc EritrDragos BaltaretuÎncă nu există evaluări
- Citokinele: Diferente Fata de Hormoni: Hormonii Clasici Circula in Concentratii Nanomolare, in Timp Ce Unele CitokineDocument5 paginiCitokinele: Diferente Fata de Hormoni: Hormonii Clasici Circula in Concentratii Nanomolare, in Timp Ce Unele CitokineIulia DragomirÎncă nu există evaluări
- Leucemia CronicaDocument66 paginiLeucemia CronicaioncatanÎncă nu există evaluări
- I. HematopoiezaDocument17 paginiI. HematopoiezaAna Maria ȘuteuÎncă nu există evaluări
- HematopoiezaDocument51 paginiHematopoiezaNico Ionaşcu100% (1)
- Hematologie Clinica ROMANA Pus Pe IntranetDocument168 paginiHematologie Clinica ROMANA Pus Pe IntranetIoana PietricicaÎncă nu există evaluări
- Ciroza HepaticaDocument18 paginiCiroza HepaticalarisaP100% (4)
- Eritropoieza Si CoagulareaDocument6 paginiEritropoieza Si Coagulareacristina_zaharia865440Încă nu există evaluări
- Leucemia CronicaDocument66 paginiLeucemia CronicaMiha Si ViliÎncă nu există evaluări
- EritrociteleDocument13 paginiEritrociteleAdrian98Încă nu există evaluări
- C3. Leucocitele Si TrombociteleDocument63 paginiC3. Leucocitele Si TrombociteleMirabelaRÎncă nu există evaluări
- Hematopoieza Normală Și PatologicăDocument19 paginiHematopoieza Normală Și Patologicăchwy48jwmyÎncă nu există evaluări
- 5.invazia Si Metastazarea 2018Document21 pagini5.invazia Si Metastazarea 2018Puscasu Alexandra TheodoraÎncă nu există evaluări
- C11 TrombocitulDocument19 paginiC11 TrombocitulMaria ChiricaÎncă nu există evaluări
- BazofilulDocument29 paginiBazofilulCristina Tomescu100% (1)
- Histologie 1 Curs 7) SangeleDocument14 paginiHistologie 1 Curs 7) SangeleMariaMaryaÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări