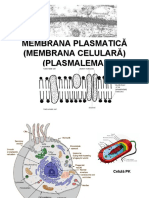
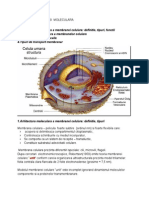
S-ar putea să vă placă și
- Ai grija de viata, de tine, de ceilalti si de naturaDe la EverandAi grija de viata, de tine, de ceilalti si de naturaÎncă nu există evaluări
- CelulaDocument10 paginiCelulaValentina Gina MuaÎncă nu există evaluări
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Definiţia Şi Funcţiile Membranelor BiologiceDocument23 paginiDefiniţia Şi Funcţiile Membranelor BiologiceDanila ElenaÎncă nu există evaluări
- Permeabilitate Şi TransportDocument14 paginiPermeabilitate Şi TransportLisa Briggs100% (1)
- Celula Eucariota Structura Si UltrastructuraDocument19 paginiCelula Eucariota Structura Si UltrastructuraFlorentina Claudia Calusaru100% (2)
- Membrana PlasmaticăDocument62 paginiMembrana PlasmaticăAndra-Amalia ArmulescuÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Membrana CelularăDocument3 paginiMembrana CelularăRobert201Încă nu există evaluări
- CELULADocument29 paginiCELULAMadalina AndreeaÎncă nu există evaluări
- Biologia Moleculara A Membranelor Celulare 2Document9 paginiBiologia Moleculara A Membranelor Celulare 2Diana BaltaÎncă nu există evaluări
- Modelul Mozaicului Fluid Al Membranei CelulareDocument21 paginiModelul Mozaicului Fluid Al Membranei CelulareZgherea MihaiÎncă nu există evaluări
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- CURS 2 Organizarea StructuralaDocument11 paginiCURS 2 Organizarea StructuralaEugen IuracÎncă nu există evaluări
- Proteinele Si Lipidele MembranareDocument13 paginiProteinele Si Lipidele MembranareFlorentina Claudia CalusaruÎncă nu există evaluări
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara Subiectedorofteim100% (1)
- Membrana Celulara PDFDocument40 paginiMembrana Celulara PDFOzana CopyShop100% (1)
- Structura Membranei CelulareDocument11 paginiStructura Membranei CelulareOana Palade100% (2)
- Lista 3 Histo Rezumat Fara 15,25,27Document15 paginiLista 3 Histo Rezumat Fara 15,25,27Manea AlexandruÎncă nu există evaluări
- Modelul Mozaicului Fluid Al Membranei CelulareDocument12 paginiModelul Mozaicului Fluid Al Membranei CelulareBuliga Claudia50% (2)
- Subiecte BiocelDocument26 paginiSubiecte BiocelDenisa AndreeaÎncă nu există evaluări
- Curs II CelulaDocument44 paginiCurs II CelulaMaria-Luminita ChiritaÎncă nu există evaluări
- Subiecte Biologie CelularaDocument11 paginiSubiecte Biologie CelularaBradeanu AndreiÎncă nu există evaluări
- Celula EucariotaDocument5 paginiCelula EucariotaStefan GhergheÎncă nu există evaluări
- Curs Biologie celulara si molecularăDocument43 paginiCurs Biologie celulara si molecularăionutbranza19Încă nu există evaluări
- Membrane 2019Document38 paginiMembrane 2019Urecheanu DorinÎncă nu există evaluări
- Referat Membrana CelularaDocument15 paginiReferat Membrana CelularaRazvan_GabrielÎncă nu există evaluări
- Curs 9Document17 paginiCurs 9aleahim_aciroivÎncă nu există evaluări
- Structura Şi Ultrastructura CeluleiDocument34 paginiStructura Şi Ultrastructura CeluleiNadeaÎncă nu există evaluări
- Membrana PlasmaticăDocument56 paginiMembrana PlasmaticăAndrada Raicu100% (1)
- Membrana PlasmaticäDocument53 paginiMembrana PlasmaticäFantasyoflove VallyÎncă nu există evaluări
- Noţiuni de CitologieDocument8 paginiNoţiuni de CitologieAnaMihaelaManguÎncă nu există evaluări
- Membrana CelularaDocument25 paginiMembrana CelularaFlavius DobreÎncă nu există evaluări
- Biologia Plantelor_RO_Manual_Celula vegetala 1_Particularitati_OrganiteDocument11 paginiBiologia Plantelor_RO_Manual_Celula vegetala 1_Particularitati_OrganiteFlorinaBurduceaÎncă nu există evaluări
- Elemente de Transport Prin Membrana CelularaDocument29 paginiElemente de Transport Prin Membrana CelularaIoana CojocaruÎncă nu există evaluări
- Celula, Proprietatile CeluleiDocument12 paginiCelula, Proprietatile CeluleiAlexÎncă nu există evaluări
- HistologieDocument51 paginiHistologieRoxana CiofalcaÎncă nu există evaluări
- Invelisul CelularDocument4 paginiInvelisul CelularNicola CreangaÎncă nu există evaluări
- Biocel Cursuri WordDocument38 paginiBiocel Cursuri Wordbubu_lina_ruza100% (2)
- BiocelDocument24 paginiBiocelficioruÎncă nu există evaluări
- Membrană Celulară2Document9 paginiMembrană Celulară2Cosmin MihoreanuÎncă nu există evaluări
- Biocel - Subiecte RezolvateDocument94 paginiBiocel - Subiecte RezolvateVictorVasile95% (20)
- Biologie Celulara SubiecteDocument11 paginiBiologie Celulara SubiecteIoana ManoleÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- Curs 1 - Istoric, Cel Procariota, Eucariota, Membrana CelularaDocument40 paginiCurs 1 - Istoric, Cel Procariota, Eucariota, Membrana CelularaAdrian UrsuÎncă nu există evaluări
- C2-ț cartilaginos și ososDocument42 paginiC2-ț cartilaginos și ososAlexandra SavinÎncă nu există evaluări
- Curs 5 - HistologieDocument63 paginiCurs 5 - HistologieAlexandra Florina BrujbanÎncă nu există evaluări
- Curs 7 Aparat Golgi & MitocondriaDocument18 paginiCurs 7 Aparat Golgi & MitocondriaBianca DenisaÎncă nu există evaluări
- Curs BiocelDocument75 paginiCurs BiocelLavinia BragaÎncă nu există evaluări
- BIOLOGIE CELULARA Curs2Document8 paginiBIOLOGIE CELULARA Curs2mewe13Încă nu există evaluări
- Dosar BioDocument17 paginiDosar BioTeodor MoroianuÎncă nu există evaluări
- Curs 3 - HistologieDocument63 paginiCurs 3 - HistologieAlexandra Florina BrujbanÎncă nu există evaluări
- CURS 1 - HistologieDocument43 paginiCURS 1 - HistologieAlexandra Florina BrujbanÎncă nu există evaluări
- Curs 4 - HistologieDocument81 paginiCurs 4 - HistologieAlexandra Florina BrujbanÎncă nu există evaluări
- Curs 5 - HistologieDocument63 paginiCurs 5 - HistologieAlexandra Florina BrujbanÎncă nu există evaluări
- Artrologie. Generalitati. Art Membrului Anterior - MEDICINA VETERINARADocument32 paginiArtrologie. Generalitati. Art Membrului Anterior - MEDICINA VETERINARAAlexandra Florina BrujbanÎncă nu există evaluări
- Scheletul Trunchilului - MEDICINA VETERINARADocument29 paginiScheletul Trunchilului - MEDICINA VETERINARAAlexandra Florina BrujbanÎncă nu există evaluări
- Craniul - MEDICINA VETERINARADocument42 paginiCraniul - MEDICINA VETERINARAAlexandra Florina BrujbanÎncă nu există evaluări
- Curs Anul I - Oase Membru PelvinDocument23 paginiCurs Anul I - Oase Membru PelvinLarissa TomlinsonÎncă nu există evaluări
- Generalitati Membrul AnteriorDocument24 paginiGeneralitati Membrul AnteriorSava RoxanaÎncă nu există evaluări