S-ar putea să vă placă și
- Celula UmanaDocument6 paginiCelula UmanaAndreea F92% (12)
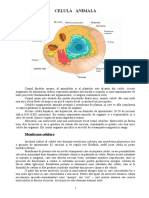
- Celula AnimalaDocument10 paginiCelula AnimalaRaluka Siliște80% (5)
- MITOCONDRIADocument5 paginiMITOCONDRIAClaudiu Bud100% (1)
- BIOCEL - MitocondriaDocument5 paginiBIOCEL - MitocondriaVoineta GheorghiuÎncă nu există evaluări
- Celula UmanaDocument13 paginiCelula Umana11224499Încă nu există evaluări
- WWW - Referat.ro-Tesuturi Si Celula30268e3fDocument8 paginiWWW - Referat.ro-Tesuturi Si Celula30268e3fAaron Wolfe100% (1)
- Celula Eucariotă. Citoplasma Și Organitele CelulareDocument4 paginiCelula Eucariotă. Citoplasma Și Organitele CelulareLeul GamerÎncă nu există evaluări
- 1 Si 2 Radiologie Curs I II Medicina Copie 2Document119 pagini1 Si 2 Radiologie Curs I II Medicina Copie 2Bianca DenisaÎncă nu există evaluări
- Curs 5Document13 paginiCurs 5Denisa NeaguÎncă nu există evaluări
- Celula UmanaDocument14 paginiCelula Umanapaulv123100% (1)
- REFERAT Celula UmanaDocument15 paginiREFERAT Celula UmanaIoana100% (2)
- Celula UmanaDocument12 paginiCelula UmanaUchiha AlexandraÎncă nu există evaluări
- Celula - BiochimieDocument6 paginiCelula - BiochimieAlice DouglasÎncă nu există evaluări
- 1Document10 pagini1Simona StanciuÎncă nu există evaluări
- Mitocondria Procese Și Structuri Implicate În Sinteza Energiei CelulareDocument19 paginiMitocondria Procese Și Structuri Implicate În Sinteza Energiei CelulareGeorgiana SimionÎncă nu există evaluări
- CelulaDocument10 paginiCelulaMonica AbbateÎncă nu există evaluări
- Organite Celulare ComuneDocument13 paginiOrganite Celulare ComuneDennisÎncă nu există evaluări
- Formatiuni Structurale CitoplasmaticeDocument7 paginiFormatiuni Structurale Citoplasmaticemyhaela_marculescuÎncă nu există evaluări
- Histologie LizozomiDocument3 paginiHistologie LizozomiMihai CodinÎncă nu există evaluări
- Celula (Structură, Proprietăţi)Document37 paginiCelula (Structură, Proprietăţi)Sorina DobreÎncă nu există evaluări
- Curs II CelulaDocument44 paginiCurs II CelulaMaria-Luminita ChiritaÎncă nu există evaluări
- Cap 3Document8 paginiCap 3Marfescu CristianÎncă nu există evaluări
- Celula Animala vs. VegetalaDocument14 paginiCelula Animala vs. VegetalaCristina Gabriela100% (1)
- Biologia Plantelor_RO_Manual_Celula vegetala 1_Particularitati_OrganiteDocument11 paginiBiologia Plantelor_RO_Manual_Celula vegetala 1_Particularitati_OrganiteFlorinaBurduceaÎncă nu există evaluări
- Curs II FizioDocument49 paginiCurs II FizioBraga Lavinia100% (1)
- Fiziologie GeneralaDocument6 paginiFiziologie GeneralaValentino PetreÎncă nu există evaluări
- Celula Si TesuturileDocument8 paginiCelula Si TesuturileCarleta Alexiana SîrbuÎncă nu există evaluări
- Celula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Document1 paginăCelula Și Fiziologia Celulară - Biologie Barron's - GinaMed 2Gabor IzabellaÎncă nu există evaluări
- BCM C3 2022Document7 paginiBCM C3 2022Matei NegoitaÎncă nu există evaluări
- Semiologie Referat Celula UmanaDocument9 paginiSemiologie Referat Celula UmanaCarmen Georgiana EnciuÎncă nu există evaluări
- Organizarea Structurala Si Functionala A CeluleiDocument19 paginiOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00Încă nu există evaluări
- Anatomie Curs 1Document16 paginiAnatomie Curs 1Danut Ancuța MihaelaÎncă nu există evaluări
- CURS 2 Organizarea StructuralaDocument11 paginiCURS 2 Organizarea StructuralaEugen IuracÎncă nu există evaluări
- L 2 Celula Structura ProprietatiDocument10 paginiL 2 Celula Structura ProprietatirahatnikitaÎncă nu există evaluări
- Histologie CitoplasmaDocument3 paginiHistologie CitoplasmaMihai CodinÎncă nu există evaluări
- Structura Şi Ultrastructura CeluleiDocument34 paginiStructura Şi Ultrastructura CeluleiNadeaÎncă nu există evaluări
- Fenomene de TransportDocument116 paginiFenomene de TransportNoțingher RaduÎncă nu există evaluări
- Celula Miracolul VTZ Evolutia Celulei EucarioteeeeeeDocument19 paginiCelula Miracolul VTZ Evolutia Celulei EucarioteeeeeeGeorgiana NițulescuÎncă nu există evaluări
- Celula EucariotaDocument5 paginiCelula EucariotaStefan GhergheÎncă nu există evaluări
- Scoala Postliceala Sanitara FundeniDocument4 paginiScoala Postliceala Sanitara FundeniAndra MihaelaÎncă nu există evaluări
- Celula Animala vs. Celula VegetalaDocument13 paginiCelula Animala vs. Celula VegetalaCristina GabrielaÎncă nu există evaluări
- Reticul EndoplasmaticDocument16 paginiReticul EndoplasmaticIrina SpoialoÎncă nu există evaluări
- Histologie CitoscheletDocument46 paginiHistologie CitoscheletAnamaria ConstantinÎncă nu există evaluări
- Celula Și Fiziologia CelularăDocument16 paginiCelula Și Fiziologia Celularăzilahivanessa799Încă nu există evaluări
- CURS 7 Histologie Si EmbriologieDocument57 paginiCURS 7 Histologie Si EmbriologieLarissa TomlinsonÎncă nu există evaluări
- Reticulul EndoplasmaticDocument3 paginiReticulul EndoplasmaticLaurentiu BaiuÎncă nu există evaluări
- Proiect Biologie: Citoplasma, RIBOSOMII, DICTIOZOMII, MITOCONTRII, LIZOZOMII, CENTROZOMII - Structura Si RolDocument5 paginiProiect Biologie: Citoplasma, RIBOSOMII, DICTIOZOMII, MITOCONTRII, LIZOZOMII, CENTROZOMII - Structura Si RolCosmin IordacheÎncă nu există evaluări
- Celula, Proprietatile CeluleiDocument12 paginiCelula, Proprietatile CeluleiAlexÎncă nu există evaluări
- Organizare Celulare IDocument14 paginiOrganizare Celulare ISmadu AdrianaÎncă nu există evaluări
- Anatomia Si Fiziologia Omului Compendiu - FragmentDocument13 paginiAnatomia Si Fiziologia Omului Compendiu - FragmentBaltaroiu MariusÎncă nu există evaluări
- Biologie Celulara Curs 2Document4 paginiBiologie Celulara Curs 2Voineta Gheorghiu100% (1)
- Dosar BioDocument17 paginiDosar BioTeodor MoroianuÎncă nu există evaluări
- Celula UmanaDocument8 paginiCelula UmanaCatalina CiocanÎncă nu există evaluări
- Anatomie - Curs1 - CelulaDocument27 paginiAnatomie - Curs1 - Celulaalfa5017Încă nu există evaluări
- Elemente Celulare Cu Rol Genetic 1Document13 paginiElemente Celulare Cu Rol Genetic 1Anda OajdeaÎncă nu există evaluări
- 5 Biofizica CelularăDocument70 pagini5 Biofizica CelularăBalint Ana MariaÎncă nu există evaluări
- OrganiteDocument5 paginiOrganiteAnda Stoica100% (1)
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Curs 3 Adezivitatea Si Recunoasterea IntercelularaDocument12 paginiCurs 3 Adezivitatea Si Recunoasterea IntercelularaBianca Denisa100% (1)
- Grile Poza 1Document2 paginiGrile Poza 1Bianca DenisaÎncă nu există evaluări
- Prezentare Spectroscopie 9 - ConvertDocument20 paginiPrezentare Spectroscopie 9 - ConvertIonela VioletaÎncă nu există evaluări
- Anato An2 S1 2Document8 paginiAnato An2 S1 2AnaRalucaDraganÎncă nu există evaluări
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- Citoplasma CelularaDocument16 paginiCitoplasma CelularaEnea DanielaÎncă nu există evaluări
- CURS 4 Transportul Prin MembranaDocument14 paginiCURS 4 Transportul Prin MembranaAlexandra GeorgianaÎncă nu există evaluări
- Curs 6 Organitele Sintezei Si Secretiei Celulare OnisorDocument15 paginiCurs 6 Organitele Sintezei Si Secretiei Celulare Onisormarkone222Încă nu există evaluări
- Curs 4Document25 paginiCurs 4Elena GheorghițăÎncă nu există evaluări
- Examen Cardio GutuDocument10 paginiExamen Cardio GutuBianca DenisaÎncă nu există evaluări
- Suprafata CeluleiDocument31 paginiSuprafata CeluleiEnea DanielaÎncă nu există evaluări
- Curs 1Document13 paginiCurs 1Mangalea IonutÎncă nu există evaluări
- Curs 3Document6 paginiCurs 3Bianca DenisaÎncă nu există evaluări
- Curs 10Document9 paginiCurs 10Bianca DenisaÎncă nu există evaluări
- Curs 8Document5 paginiCurs 8Bianca DenisaÎncă nu există evaluări
- Curs 9Document21 paginiCurs 9Bianca DenisaÎncă nu există evaluări
- Curs 7Document8 paginiCurs 7Bianca DenisaÎncă nu există evaluări
- 11 Programul de Asigurare Şi Îmbunătăţire A CalităţiiDocument10 pagini11 Programul de Asigurare Şi Îmbunătăţire A CalităţiiBianca DenisaÎncă nu există evaluări
- Curs 6Document20 paginiCurs 6Bianca DenisaÎncă nu există evaluări
- Radiologie Cursul 12 MGDocument52 paginiRadiologie Cursul 12 MGBianca DenisaÎncă nu există evaluări
- Curs 5Document10 paginiCurs 5Bianca DenisaÎncă nu există evaluări
- Contabilitatea Operatiunilor Cu CarduriDocument17 paginiContabilitatea Operatiunilor Cu CarduriBianca DenisaÎncă nu există evaluări
- 16 Dec Seminar Aplicatii Operatiuni DiverseDocument2 pagini16 Dec Seminar Aplicatii Operatiuni DiverseBianca DenisaÎncă nu există evaluări
- Aplicatie Titluri de TranzactieDocument5 paginiAplicatie Titluri de TranzactieBianca DenisaÎncă nu există evaluări
- PDFsam MergeDocument118 paginiPDFsam MergeBianca DenisaÎncă nu există evaluări
- Aplicatie Operaţii de FactoringDocument2 paginiAplicatie Operaţii de FactoringBianca DenisaÎncă nu există evaluări
- 7 Contabilitatea Operatiunilor de FactoringDocument10 pagini7 Contabilitatea Operatiunilor de FactoringBianca DenisaÎncă nu există evaluări
- 7 Desfăşurarea Misiunii de Audit InternDocument18 pagini7 Desfăşurarea Misiunii de Audit InternBianca DenisaÎncă nu există evaluări