S-ar putea să vă placă și
- Fiziologie VegDocument11 paginiFiziologie VegAda Adishor67% (3)
- Fiziologia Plantelor CursDocument112 paginiFiziologia Plantelor CursMIHAIL - CĂTĂLIN CHIREAÎncă nu există evaluări
- L6 IeaDocument6 paginiL6 IeaSerbanCorneliuÎncă nu există evaluări
- Culturi in VitroDocument2 paginiCulturi in VitroRoma Plamadeala100% (1)
- Combaterea Integrata A DaunatorilorDocument17 paginiCombaterea Integrata A DaunatorilorDorina LucaÎncă nu există evaluări
- Dokumen - Tips - Proiect Didactic Circulatia Sangelui Viidocx 1Document5 paginiDokumen - Tips - Proiect Didactic Circulatia Sangelui Viidocx 1Mihaela VizinÎncă nu există evaluări
- Ev cl.9 Sist .ReproducDocument2 paginiEv cl.9 Sist .ReproducLena Bondari-CebotariÎncă nu există evaluări
- Amidon 7Document1 paginăAmidon 7Марина МаринаÎncă nu există evaluări
- Algele RosiiDocument60 paginiAlgele RosiiDannaVasi8747100% (1)
- Tema 2 Adaptarea Plantelor La Diverse Conditii de MediuDocument68 paginiTema 2 Adaptarea Plantelor La Diverse Conditii de MediuLiviaÎncă nu există evaluări
- 1-Biotehnologii AgricoleDocument2 pagini1-Biotehnologii AgricoleDenisaPricope100% (1)
- Fructul. Diversitatea-Fructelor.Document3 paginiFructul. Diversitatea-Fructelor.Ion TascaÎncă nu există evaluări
- CLASIFICARE STIINTIFICA F SoareluiDocument4 paginiCLASIFICARE STIINTIFICA F SoareluiGabriela MihaelaÎncă nu există evaluări
- Proiect Didactic Clasa 6, Diversitatea Org. Pe TeraDocument6 paginiProiect Didactic Clasa 6, Diversitatea Org. Pe TeraBeschieruVadim0% (1)
- IERBARDocument27 paginiIERBARVasile ElenaÎncă nu există evaluări
- Curs NR.4Document10 paginiCurs NR.4anca_gheormaÎncă nu există evaluări
- Lucrare Practica NR 2 Structura Plantei Cl8Document2 paginiLucrare Practica NR 2 Structura Plantei Cl8Artur PopoviciÎncă nu există evaluări
- Tehnologie de Cultura Si Ameliorarea Legumelor, Burnichi FloareaDocument109 paginiTehnologie de Cultura Si Ameliorarea Legumelor, Burnichi FloareaToma MitelÎncă nu există evaluări
- Funcțiile Vitale La PlanteDocument22 paginiFuncțiile Vitale La PlanteMihai Boinceanu2004Încă nu există evaluări
- BotanicaDocument14 paginiBotanicaAlexandra StanÎncă nu există evaluări
- 4 Adaptarea Plantelor La Diverse Conditii de MediuDocument66 pagini4 Adaptarea Plantelor La Diverse Conditii de MediuIonela ZubcoÎncă nu există evaluări
- Cultura CapsunuluiDocument8 paginiCultura CapsunuluiMariusTudoreanÎncă nu există evaluări
- 7 - Rezistența Plantelor La SecetăDocument22 pagini7 - Rezistența Plantelor La SecetăMihaela Vizin100% (1)
- 7 - Rezistența Plantelor La SecetăDocument22 pagini7 - Rezistența Plantelor La SecetăMihaela Vizin100% (1)
- Ovazul in Republica MoldovaDocument8 paginiOvazul in Republica MoldovaLaurentiu MunteanuÎncă nu există evaluări
- LichenophytaDocument18 paginiLichenophytaCamelia Radulescu100% (1)
- Istoria Dezvoltării Științelor Botanice În Epoca RenașteriiDocument15 paginiIstoria Dezvoltării Științelor Botanice În Epoca RenașteriiMihaela Vizin100% (1)
- 1 CaracteristicileSoluluiDocument20 pagini1 CaracteristicileSoluluixhuljaÎncă nu există evaluări
- S 21 Bolile Plantelor CultivateDocument11 paginiS 21 Bolile Plantelor CultivateMădălina GrigorașÎncă nu există evaluări
- Comportament Alimentarla Animalele PradatoareDocument12 paginiComportament Alimentarla Animalele PradatoareSvetlana Raifura100% (1)
- Cicluri BiogeochimiceDocument83 paginiCicluri BiogeochimicesbotacondÎncă nu există evaluări
- Proiect IngrasaminteDocument4 paginiProiect IngrasaminteCiulei SebastianÎncă nu există evaluări
- Coslet Liliana - Rezistenta Plantelor La FrigDocument13 paginiCoslet Liliana - Rezistenta Plantelor La FrigLiliana Vizitiu Coșleț100% (1)
- Rezistenta La Ger Si Inghet, CalireaDocument13 paginiRezistenta La Ger Si Inghet, CalireaveronicaÎncă nu există evaluări
- Curs Fiziologia PlantelorDocument33 paginiCurs Fiziologia PlantelorSebastian Oancea100% (1)
- SămânțaDocument3 paginiSămânțaGeorgiana BusuÎncă nu există evaluări
- Fiziologia VegetalăDocument34 paginiFiziologia VegetalăNatalia Atamaniuc100% (1)
- 10 - Rezistența Plantelor La Poluarea Mediului de ViațăDocument12 pagini10 - Rezistența Plantelor La Poluarea Mediului de ViațăMihaela VizinÎncă nu există evaluări
- Fisa de Observatie Bolile Pomilor FructiferiDocument2 paginiFisa de Observatie Bolile Pomilor FructiferiVlad Alexandru MihaiÎncă nu există evaluări
- Stiinte BiologiceDocument17 paginiStiinte BiologiceDumitru SanduÎncă nu există evaluări
- Polaritatea Si Corelatia La PlanteDocument3 paginiPolaritatea Si Corelatia La PlanteNicu VladÎncă nu există evaluări
- Curs NR.2Document12 paginiCurs NR.2Robert Sindila-BurceaÎncă nu există evaluări
- Curs 1Document24 paginiCurs 1calinb25Încă nu există evaluări
- SamantaDocument21 paginiSamantaPetru GiraÎncă nu există evaluări
- TaxonomieDocument25 paginiTaxonomieAndrei CherecheșuÎncă nu există evaluări
- Vocabularul PlanteiDocument3 paginiVocabularul PlanteiViorica CebanÎncă nu există evaluări
- Plante ToxiceDocument8 paginiPlante ToxiceAlexaelisaÎncă nu există evaluări
- Curs 1Document5 paginiCurs 1Nicu VladÎncă nu există evaluări
- PROIECT DIDACTIC DE LUNG - DURAT - LA Biologie Cl. 9Document7 paginiPROIECT DIDACTIC DE LUNG - DURAT - LA Biologie Cl. 9DianaÎncă nu există evaluări
- Comport. Aliment. Anim. Pradatoare - Prezentare.Document33 paginiComport. Aliment. Anim. Pradatoare - Prezentare.olesea100% (4)
- AgrotehnicaDocument16 paginiAgrotehnicaCristina PaunÎncă nu există evaluări
- Cl. 9 C - Bioritmul SezonierDocument9 paginiCl. 9 C - Bioritmul SezonierdanielÎncă nu există evaluări
- Fisa de Lucru CelenterateDocument2 paginiFisa de Lucru CelenterateClaudinia Cazac100% (1)
- Hormonii Vegetali2222.Document7 paginiHormonii Vegetali2222.Georgiana GianniniÎncă nu există evaluări
- Importanta Plantelor in Viata Omului: 3 Pagina 01/10Document9 paginiImportanta Plantelor in Viata Omului: 3 Pagina 01/10DianaÎncă nu există evaluări
- Lucrare Practica 7Document3 paginiLucrare Practica 7Adela Popa100% (1)
- Dezvoltarea PlantelorDocument12 paginiDezvoltarea PlantelorSerbanCorneliuÎncă nu există evaluări
- Codiaeum VariegatumDocument21 paginiCodiaeum VariegatumMarcela MalachiÎncă nu există evaluări
- Rebus Trecerea La AngiospermeDocument2 paginiRebus Trecerea La AngiospermeАндрей Митрозор100% (1)
- Bilantul Hidric Al PlantelorDocument19 paginiBilantul Hidric Al PlantelorPopescu OctavianÎncă nu există evaluări
- Bioritmul PlantelorDocument4 paginiBioritmul PlantelorMonica Hristoforov0% (1)
- EcofiziologieDocument9 paginiEcofiziologieIrina UngureanuÎncă nu există evaluări
- 6 - Rezistența Plantelor La ArșițăDocument4 pagini6 - Rezistența Plantelor La ArșițăMihaela VizinÎncă nu există evaluări
- 1 AdaptariDocument6 pagini1 AdaptariAlina Stefania100% (1)
- "Gîndul Nescris Este o Comoară Pierdută". (D.I.Mendeleev)Document1 pagină"Gîndul Nescris Este o Comoară Pierdută". (D.I.Mendeleev)Mihaela VizinÎncă nu există evaluări
- 2 - Bazele Fiziologice Ale Adaptării Plantelor La Condițiile NefavorabileDocument4 pagini2 - Bazele Fiziologice Ale Adaptării Plantelor La Condițiile NefavorabileMihaela VizinÎncă nu există evaluări
- 12 - Autoreglare Creșterii Și Dezvoltării PlantelorDocument5 pagini12 - Autoreglare Creșterii Și Dezvoltării PlantelorMihaela VizinÎncă nu există evaluări
- Prezentare Sistemul Locomotor PPTDocument8 paginiPrezentare Sistemul Locomotor PPTMihaela VizinÎncă nu există evaluări
- 6 - Rezistența Plantelor La ArșițăDocument4 pagini6 - Rezistența Plantelor La ArșițăMihaela VizinÎncă nu există evaluări
- Prezentare Stil de Viata Sanat.Document12 paginiPrezentare Stil de Viata Sanat.Mihaela VizinÎncă nu există evaluări
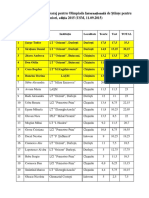
- Rezultate Barajolimpiada Internationala de Stiinte Pentru Juniori2015Document1 paginăRezultate Barajolimpiada Internationala de Stiinte Pentru Juniori2015Mihaela VizinÎncă nu există evaluări
- Program Congres MD PL RO Aprilie22Document18 paginiProgram Congres MD PL RO Aprilie22Mihaela VizinÎncă nu există evaluări
- Aerul Proiec de LectieDocument2 paginiAerul Proiec de LectieMihaela VizinÎncă nu există evaluări
- Ovcerenco - Texte de Lectii - PsihopedagogiaDocument128 paginiOvcerenco - Texte de Lectii - PsihopedagogiaMihaela VizinÎncă nu există evaluări
- 258 2012 Biologie EDocument9 pagini258 2012 Biologie EMihaela VizinÎncă nu există evaluări
- E CH Clasa8 SubstantaDocument2 paginiE CH Clasa8 SubstantaMihaela VizinÎncă nu există evaluări
- Tema 5 La GrigorceaDocument3 paginiTema 5 La GrigorceaMihaela VizinÎncă nu există evaluări
- Test Fenomene TermiceDocument1 paginăTest Fenomene TermiceMihaela Vizin0% (1)
- Evaluare CL 9 Biologie Sisteme Coordonare Si Integrare in MediuDocument3 paginiEvaluare CL 9 Biologie Sisteme Coordonare Si Integrare in MediuMihaela VizinÎncă nu există evaluări
- ПрезентацияDocument3 paginiПрезентацияMihaela VizinÎncă nu există evaluări
- Oxizii PSDDocument7 paginiOxizii PSDMihaela VizinÎncă nu există evaluări
- Tema Pentru AcasaDocument26 paginiTema Pentru AcasaMihaela VizinÎncă nu există evaluări