S-ar putea să vă placă și
- Introducere în psihologia emoțiilor: De la Darwin la neuroștiințe, ce sunt și cum funcționează emoțiileDe la EverandIntroducere în psihologia emoțiilor: De la Darwin la neuroștiințe, ce sunt și cum funcționează emoțiileEvaluare: 4.5 din 5 stele4.5/5 (8)
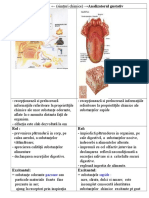
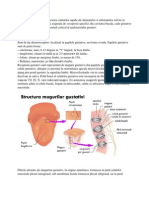
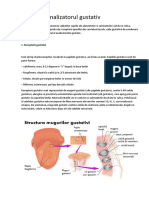
- Analizatorul GustativDocument12 paginiAnalizatorul GustativAlexandra Bădescu100% (1)
- Curs 5 Senzatiile. Tipuri de SenzatiiDocument33 paginiCurs 5 Senzatiile. Tipuri de SenzatiiIuliana Catalina100% (1)
- Analizatorul Olfactic Si GustativDocument14 paginiAnalizatorul Olfactic Si GustativPirvu RamonaÎncă nu există evaluări
- Analizatorul GustativDocument11 paginiAnalizatorul GustativEnascuta AlinaÎncă nu există evaluări
- Tipuri de SenzatiiDocument33 paginiTipuri de SenzatiiGeorgiana Tisianu100% (1)
- 8 Analizatorul Olfactiv Si GustativDocument3 pagini8 Analizatorul Olfactiv Si GustativRadu Andrei CătinasÎncă nu există evaluări
- Analizatorul GustativDocument41 paginiAnalizatorul GustativPetecariu Ioana MariaÎncă nu există evaluări
- Organe de Simţ PDFDocument118 paginiOrgane de Simţ PDFVerinceanu AlexandruÎncă nu există evaluări
- Analizatori SenzorialiDocument31 paginiAnalizatori SenzorialiBucur IoanaÎncă nu există evaluări
- Curs 5 Senzatiile. Tipuri de SenzatiiDocument30 paginiCurs 5 Senzatiile. Tipuri de SenzatiiOfelia Violeta RacovițăÎncă nu există evaluări
- Ref Proiectie CorticalaDocument8 paginiRef Proiectie Corticalaalutus20060% (1)
- Coles Cosmina Pipp An 1, Grupa 1 Fundamentele Psihologiei Universitatea ,,ovidius'' Din ConstantaDocument10 paginiColes Cosmina Pipp An 1, Grupa 1 Fundamentele Psihologiei Universitatea ,,ovidius'' Din ConstantaMina CosminaÎncă nu există evaluări
- Analizatorul OlfactivDocument3 paginiAnalizatorul OlfactivDenise Maria100% (1)
- Senzatii GustativeDocument3 paginiSenzatii GustativeRussu DumitruÎncă nu există evaluări
- Asemanari Si Deosebiri Dintre Receptorii Gustativi Si OlfactiviDocument6 paginiAsemanari Si Deosebiri Dintre Receptorii Gustativi Si OlfactiviRaluca PădurariuÎncă nu există evaluări
- AnalizatoriiDocument4 paginiAnalizatoriiCristiana MorariuÎncă nu există evaluări
- ANALIZATORIIDocument7 paginiANALIZATORIITaniaPopescuÎncă nu există evaluări
- Curs 13 AnalizatoriiDocument58 paginiCurs 13 AnalizatoriiRamona MihaelaÎncă nu există evaluări
- Fiziologia Analizatorului OlfactivDocument5 paginiFiziologia Analizatorului OlfactivIrina Georgiana100% (1)
- Analizatori Curs An 2 2014Document85 paginiAnalizatori Curs An 2 2014Marta BabiiÎncă nu există evaluări
- ReceptoriiDocument8 paginiReceptoriidoinitadmdÎncă nu există evaluări
- Tipuri de SenzatiiDocument32 paginiTipuri de SenzatiiGeo Georgiana91% (11)
- SimturileDocument6 paginiSimturileIacob DariaÎncă nu există evaluări
- Analizatorul Olfactiv, GustativDocument7 paginiAnalizatorul Olfactiv, Gustativdeeadiaconu0% (1)
- Analizatorul OflfactivDocument12 paginiAnalizatorul OflfactivTeodora CoroiÎncă nu există evaluări
- Aparatul OlfactivDocument3 paginiAparatul OlfactivTudorVasluianuÎncă nu există evaluări
- Fizio 1Document6 paginiFizio 1Constantin UntilaÎncă nu există evaluări
- Analizatorul GustativDocument6 paginiAnalizatorul GustativCristina LoveÎncă nu există evaluări
- Tema 3. SenzatiiDocument30 paginiTema 3. SenzatiiDina ManoliÎncă nu există evaluări
- Analizatorul GustativDocument6 paginiAnalizatorul GustativAndreea Deea100% (3)
- Analizatorul GustativDocument5 paginiAnalizatorul GustativGgy VictorÎncă nu există evaluări
- Analizatorul GustativDocument2 paginiAnalizatorul GustativBianca BrlcÎncă nu există evaluări
- Analizatorul GustativDocument4 paginiAnalizatorul GustativLory EelenaÎncă nu există evaluări
- Fundamentele Psihologiei - Senzatia OlfactivaDocument3 paginiFundamentele Psihologiei - Senzatia OlfactivapetresculorenaÎncă nu există evaluări
- Analizatorul Gustativ PDFDocument9 paginiAnalizatorul Gustativ PDFMirela OsmanÎncă nu există evaluări
- Curs 4 AsaDocument8 paginiCurs 4 AsabogdanÎncă nu există evaluări
- Analizatorul Gustativ: Simțul MirosuluiDocument4 paginiAnalizatorul Gustativ: Simțul MirosuluiProfu' de biologieÎncă nu există evaluări
- Analizatorul OlfactivDocument9 paginiAnalizatorul Olfactivpaun_alexandra89Încă nu există evaluări
- Gustul Si MirosulDocument4 paginiGustul Si Mirosulallyk0% (1)
- Analizatorii Olfactiv Si Gustativ (S)Document12 paginiAnalizatorii Olfactiv Si Gustativ (S)svetocika2100% (1)
- Analizatorul GustativDocument10 paginiAnalizatorul GustativBeatrice ScarlatÎncă nu există evaluări
- Importanța MirosuluiDocument8 paginiImportanța MirosuluiAlina AndriutaÎncă nu există evaluări
- Analizatorul GustativDocument5 paginiAnalizatorul GustativFaieryÎncă nu există evaluări
- Analizatorul Gustativ Si Vestibular - Curs 12Document5 paginiAnalizatorul Gustativ Si Vestibular - Curs 12Ioana Navalici100% (1)
- Analizatorul GustativDocument5 paginiAnalizatorul GustativVasilica BekoÎncă nu există evaluări
- Document Fără TitluDocument5 paginiDocument Fără TitluGiuliaÎncă nu există evaluări
- Analizatorul GustativDocument4 paginiAnalizatorul GustativAndreiÎncă nu există evaluări
- GustDocument5 paginiGustElena CalapodÎncă nu există evaluări
- Sensibilitatea OlfactivaDocument3 paginiSensibilitatea OlfactivaRussu DumitruÎncă nu există evaluări
- AparatulDocument9 paginiAparatulLarisa AlexandruÎncă nu există evaluări
- BBBBBBBBBBBBBBBBBDocument9 paginiBBBBBBBBBBBBBBBBBValeria SircuÎncă nu există evaluări
- Analizatorul Olfactiv Şi GustativDocument20 paginiAnalizatorul Olfactiv Şi GustativAndre1boloÎncă nu există evaluări
- Analizatorul Gustativ-VinurileDocument16 paginiAnalizatorul Gustativ-VinurileAndreea Patrascu100% (1)
- Fsof LPDocument23 paginiFsof LPMadalina DelicoteÎncă nu există evaluări
- Curs 10Document6 paginiCurs 10sham harbaÎncă nu există evaluări
- Curs 6Document13 paginiCurs 6sham harbaÎncă nu există evaluări
- Curs 5Document14 paginiCurs 5sham harbaÎncă nu există evaluări
- Curs 3Document21 paginiCurs 3sham harbaÎncă nu există evaluări