S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 4 din 5 stele4/5 (8)
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Ofta Curs TotDocument641 paginiOfta Curs TotDarie AntonescuÎncă nu există evaluări
- Cursuri Tot PDFDocument670 paginiCursuri Tot PDFMitrea Andreea CristinaÎncă nu există evaluări
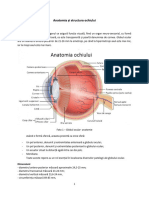
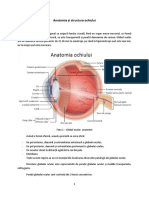
- Anatomia Analizatorului VizualDocument51 paginiAnatomia Analizatorului VizualMariaBoicu100% (5)
- Ochi UrecheDocument11 paginiOchi UrecheGabriela NeacsuÎncă nu există evaluări
- Curs 1 ANATOMIA ANALIZATORULUI VIZUALDocument34 paginiCurs 1 ANATOMIA ANALIZATORULUI VIZUALPop AlexÎncă nu există evaluări
- Curs Organele de Simt - Organul VazuluiDocument6 paginiCurs Organele de Simt - Organul VazuluiCasiana GuiÎncă nu există evaluări
- Neurofiziologia Analizatorilor SenzorialiDocument27 paginiNeurofiziologia Analizatorilor SenzorialiSiMonNykÎncă nu există evaluări
- Curs 1 Anatomia Analizatorului VizualDocument35 paginiCurs 1 Anatomia Analizatorului VizualLiviu LivaxÎncă nu există evaluări
- Analizatorul VizualDocument12 paginiAnalizatorul VizualMîndru Lara100% (1)
- Organe de SimtDocument10 paginiOrgane de SimtIuliana BicecÎncă nu există evaluări
- Analizatorul VizualDocument6 paginiAnalizatorul VizualIrina ASÎncă nu există evaluări
- Traumatisme OculareDocument8 paginiTraumatisme Ocularearyadna_9Încă nu există evaluări
- Anatomia Analizatorului VizualDocument28 paginiAnatomia Analizatorului VizualDaniela Maria GaneaÎncă nu există evaluări
- AnatomieDocument28 paginiAnatomieEduard TomițaÎncă nu există evaluări
- Organe de Simt Curs DefinitivDocument26 paginiOrgane de Simt Curs DefinitivIoana GrayÎncă nu există evaluări
- Analizato RiiDocument9 paginiAnalizato RiiLaura MariaÎncă nu există evaluări
- 8 Organe de SimtDocument5 pagini8 Organe de SimtzennmotoÎncă nu există evaluări
- Analizatorul Vizual Anus.Document40 paginiAnalizatorul Vizual Anus.anisoaraÎncă nu există evaluări
- Analizatorul Vizual Este Un Sistem Multifuncţional Complex Format Din 3 Subsisteme - OdtDocument36 paginiAnalizatorul Vizual Este Un Sistem Multifuncţional Complex Format Din 3 Subsisteme - OdtRoxana CioflîncÎncă nu există evaluări
- Analizatorul VizualDocument41 paginiAnalizatorul Vizualmariana77100% (2)
- ANALIZATORII. 2pptDocument47 paginiANALIZATORII. 2pptCasandra TudoracheÎncă nu există evaluări
- Tiutiuca OftalmologieDocument146 paginiTiutiuca Oftalmologiemirea claudiaÎncă nu există evaluări
- Analizatorul OpticDocument25 paginiAnalizatorul OpticOana OpraÎncă nu există evaluări
- Anatomia Analizorului OpticDocument64 paginiAnatomia Analizorului OpticLina CitulencoÎncă nu există evaluări
- GlaucomulDocument38 paginiGlaucomulD@m@ige100% (2)
- Ingrijirea Pacientului Cu GlaucomDocument27 paginiIngrijirea Pacientului Cu GlaucomKarla Dana100% (6)
- Oftalmologie Curs1-Anatomia AP. VizualDocument12 paginiOftalmologie Curs1-Anatomia AP. VizualsymohsnÎncă nu există evaluări
- CONJUNCTIVITADocument47 paginiCONJUNCTIVITACIPSITÎncă nu există evaluări
- Histologie AnalizatoriDocument6 paginiHistologie AnalizatoriGiuliaGoth00Încă nu există evaluări
- Lucru IndividualDocument11 paginiLucru IndividualIgor Igor DasÎncă nu există evaluări
- Analizator VizualDocument6 paginiAnalizator VizualCristiana MorariuÎncă nu există evaluări
- OchiulDocument31 paginiOchiulRadu Stefan MaricaÎncă nu există evaluări
- Organe de Simt GrigorasDocument16 paginiOrgane de Simt GrigorasScolobiuc Emma ElenaÎncă nu există evaluări
- Anatomia Analizatorului VizualDocument51 paginiAnatomia Analizatorului VizualFlorentina Mihaela IonescuÎncă nu există evaluări
- OftalmologieDocument58 paginiOftalmologieMadPiuÎncă nu există evaluări
- Anatomia, Fiziologia Si Patologia OchiuluiDocument11 paginiAnatomia, Fiziologia Si Patologia OchiuluiAnastasiaÎncă nu există evaluări
- Anatomia Globului Ocular-11839-31335Document31 paginiAnatomia Globului Ocular-11839-31335Lefter GheorgheÎncă nu există evaluări
- Anatomia Globului OcularDocument34 paginiAnatomia Globului OcularroxanaperianuÎncă nu există evaluări
- Organe de SimtDocument65 paginiOrgane de SimtClaudia ScpÎncă nu există evaluări
- CURS 1 MG Anatomia Și Fiziologia Gloooobului OcularDocument14 paginiCURS 1 MG Anatomia Și Fiziologia Gloooobului OcularcetinaÎncă nu există evaluări
- Anatomia Si Structura OchiuluiDocument10 paginiAnatomia Si Structura OchiuluiAmalia TircomnicuÎncă nu există evaluări
- Anatomia Si Structura OchiuluiDocument22 paginiAnatomia Si Structura OchiuluiCătălin VanÎncă nu există evaluări
- 01.05 - RetinaDocument6 pagini01.05 - RetinaciprianÎncă nu există evaluări
- Analizatorul VizualDocument7 paginiAnalizatorul VizualDaniela MariaÎncă nu există evaluări
- Analizatorul Vizual - Curs An II, Fac. Medicina (Umana)Document17 paginiAnalizatorul Vizual - Curs An II, Fac. Medicina (Umana)Anca Maria Istrate-OfițeruÎncă nu există evaluări
- Lucrare de Diploma - GlaucomulDocument39 paginiLucrare de Diploma - GlaucomulkittystefymÎncă nu există evaluări
- Anatomia Globului Ocular PrezDocument31 paginiAnatomia Globului Ocular PrezCorina AlexandrescuÎncă nu există evaluări
- Substante Agresive PT VedereDocument96 paginiSubstante Agresive PT VedereIonut HutuleacÎncă nu există evaluări
- Anatomia Globului OcularDocument72 paginiAnatomia Globului Ocularסולימאן פוקראÎncă nu există evaluări
- Curs 12Document18 paginiCurs 12Zamfirescu Mihaela-ElenaÎncă nu există evaluări
- Curs 2Document51 paginiCurs 2DragoșMarcuÎncă nu există evaluări
- Curs-13 (Sem. Ap. Ren)Document37 paginiCurs-13 (Sem. Ap. Ren)Ionelv IvpÎncă nu există evaluări
- Curs 11Document13 paginiCurs 11Zamfirescu Mihaela-ElenaÎncă nu există evaluări
- Cursul 14Document20 paginiCursul 14Ghitiu Nicolae AlexandruÎncă nu există evaluări
- curs 6-semioDocument65 paginicurs 6-semioAndreea CristinaÎncă nu există evaluări
- Curs 8 SemioDocument32 paginiCurs 8 SemioElenaCondratscribdÎncă nu există evaluări
- Cursul 15 Semiologie MedicalaDocument24 paginiCursul 15 Semiologie MedicalaElenaCondratscribdÎncă nu există evaluări
- Curs 6 MedicalaDocument12 paginiCurs 6 MedicalaZamfirescu Mihaela-ElenaÎncă nu există evaluări
- 5 Ci, Ap, Im, Endo, PeriDocument15 pagini5 Ci, Ap, Im, Endo, PerimarkopavloskiÎncă nu există evaluări
- Curs 03 - Inspectia Generala A OrganismuluiDocument40 paginiCurs 03 - Inspectia Generala A OrganismuluiKimskiusÎncă nu există evaluări
- Curs 5Document32 paginiCurs 5Madalina TrandafirÎncă nu există evaluări
- Curs 7Document22 paginiCurs 7Zamfirescu Mihaela-ElenaÎncă nu există evaluări
- Curs nr.1 SemioDocument21 paginiCurs nr.1 SemioIoana ToaderÎncă nu există evaluări
- Curs nr2 S1Document31 paginiCurs nr2 S1Christopher HatfieldÎncă nu există evaluări