S-ar putea să vă placă și
- Superintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateDe la EverandSuperintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateÎncă nu există evaluări
- TOT CE TREBUIE SĂ ȘTII CA SĂ FII AS LA BIOLOGIE ÎNTR-UN DITAMAI CAIETULDe la EverandTOT CE TREBUIE SĂ ȘTII CA SĂ FII AS LA BIOLOGIE ÎNTR-UN DITAMAI CAIETULEvaluare: 5 din 5 stele5/5 (3)
- Microbiota NormalaDocument12 paginiMicrobiota NormalaAnca Elena100% (1)
- Microbiota UmanaDocument20 paginiMicrobiota UmanaApostu AncaÎncă nu există evaluări
- Microbiologie MedicalaDocument13 paginiMicrobiologie MedicalaIoana100% (1)
- Capitolele 8 Si 9Document20 paginiCapitolele 8 Si 9Gavrilescu AlexandraÎncă nu există evaluări
- Proiect ParodontologieDocument8 paginiProiect ParodontologieMaria PopÎncă nu există evaluări
- CURS 13 Caracteristicile BiofilmuluiDocument16 paginiCURS 13 Caracteristicile BiofilmuluiElena GiuscaÎncă nu există evaluări
- Elaborarea Nr.4-Natalia DoniciDocument22 paginiElaborarea Nr.4-Natalia DoniciButnaru TatianaÎncă nu există evaluări
- PlacaDocument5 paginiPlacaIoana Maria AvramÎncă nu există evaluări
- Microbiota Si Principalele BacteriiDocument78 paginiMicrobiota Si Principalele BacteriiOlga BurduniucÎncă nu există evaluări
- Bacteriologie Curs 10Document127 paginiBacteriologie Curs 10AnamariaMntÎncă nu există evaluări
- C4. Flora Normala, Cavitatea Bucala, Flora Normala A Cavitatii BucaleDocument42 paginiC4. Flora Normala, Cavitatea Bucala, Flora Normala A Cavitatii BucaleP?TR�NJEI BOGDAN- ALEXANDRUÎncă nu există evaluări
- 3microbiota PDFDocument10 pagini3microbiota PDFVictoria GlavanÎncă nu există evaluări
- C 2 Ecosistemul BucalDocument32 paginiC 2 Ecosistemul BucalWass KutseyÎncă nu există evaluări
- Apararea La Nivelul Cavitatii Orale - ScurtDocument7 paginiApararea La Nivelul Cavitatii Orale - Scurtram1089Încă nu există evaluări
- Lucrarea Practica N 6Document11 paginiLucrarea Practica N 6Mihaela CaruntuÎncă nu există evaluări
- Colonizarea Microbian A Omului. Microbiota Indigen .Document78 paginiColonizarea Microbian A Omului. Microbiota Indigen .Adriana AdrianaÎncă nu există evaluări
- Curs 1 - Omul Si MicroorganismeleDocument12 paginiCurs 1 - Omul Si MicroorganismeleIoana CoseruÎncă nu există evaluări
- Microflora Normala A Organismului UmanDocument15 paginiMicroflora Normala A Organismului UmanGhenadii Betivu100% (3)
- Bacterii ProbioticeDocument50 paginiBacterii ProbioticeAndreea CameliaÎncă nu există evaluări
- CURS 12 Flora Normală A OrganismuluiDocument12 paginiCURS 12 Flora Normală A OrganismuluiElena GiuscaÎncă nu există evaluări
- Etiologia Cariei DentareDocument9 paginiEtiologia Cariei DentareOctavian CalaraşÎncă nu există evaluări
- Ecosistemul Cavității OraleDocument6 paginiEcosistemul Cavității OraleValentin MihneaÎncă nu există evaluări
- Curs 1 ModDocument67 paginiCurs 1 ModVlad HangaÎncă nu există evaluări
- LucrareaDocument100 paginiLucrareaKaterina BogatayaÎncă nu există evaluări
- Proprietatile Microorganismelor PatogeneDocument34 paginiProprietatile Microorganismelor PatogenesupravietuireÎncă nu există evaluări
- Microbiota Si Principalele BacteriiDocument71 paginiMicrobiota Si Principalele BacteriiShugani KaterinaÎncă nu există evaluări
- CURS-4-Relatii Micro-MacroorganismDocument19 paginiCURS-4-Relatii Micro-MacroorganismCristina DobrinÎncă nu există evaluări
- Microbiologia OualorDocument6 paginiMicrobiologia OualorToader Ana-MariaÎncă nu există evaluări
- Placa Dentara Modif Fara PozeDocument18 paginiPlaca Dentara Modif Fara PozeOprea RoxanaÎncă nu există evaluări
- Genul StaphylococcusDocument57 paginiGenul StaphylococcusDora PopÎncă nu există evaluări
- RJP 2018 2 RO Art-01Document5 paginiRJP 2018 2 RO Art-01Alexandru Codrin-IonutÎncă nu există evaluări
- Curs 5Document16 paginiCurs 5Daniel DiduÎncă nu există evaluări
- Etiopatogenia Cariei DentareDocument13 paginiEtiopatogenia Cariei DentareElena BejanÎncă nu există evaluări
- Power Point Microb Spec BiotDocument136 paginiPower Point Microb Spec BiotDima CebotariÎncă nu există evaluări
- Interactiuni Ale Bacteriilor Patogene Cu Organismul UmanDocument15 paginiInteractiuni Ale Bacteriilor Patogene Cu Organismul UmanCosmin CosticaÎncă nu există evaluări
- Diareea La CopilDocument27 paginiDiareea La CopillilianaÎncă nu există evaluări
- BIOFILMUL - Curs MDDocument38 paginiBIOFILMUL - Curs MDRuxandra VîrlanÎncă nu există evaluări
- Curs 8 Relatii Microorganism-GazdaDocument40 paginiCurs 8 Relatii Microorganism-GazdaFlorin DumitracheÎncă nu există evaluări
- Curs 2 PrevDocument13 paginiCurs 2 PrevktylauraÎncă nu există evaluări
- Curs MicrobiDocument46 paginiCurs MicrobiAd AdrianaÎncă nu există evaluări
- Regnul MoneraDocument23 paginiRegnul MoneraAbrudan PatriciaÎncă nu există evaluări
- UntitledDocument15 paginiUntitledElenaÎncă nu există evaluări
- MicroDocument45 paginiMicrokolo1234777Încă nu există evaluări
- Microbiologie DFCCT Part1Document48 paginiMicrobiologie DFCCT Part1Gianina Madalina SachelaruÎncă nu există evaluări
- Placa Bacteriana PPT BencovschiDocument10 paginiPlaca Bacteriana PPT BencovschiMl BeÎncă nu există evaluări
- Mucoasa de Tapetare Face Legatura Cu Structurile Subiacente Prin IntermediulDocument36 paginiMucoasa de Tapetare Face Legatura Cu Structurile Subiacente Prin IntermediulAlexandra MagaleasÎncă nu există evaluări
- t2 FizioDocument39 paginit2 FizioMorojnoeÎncă nu există evaluări
- BIOFILMUL - Curs MDDocument38 paginiBIOFILMUL - Curs MDTeodora OanceaÎncă nu există evaluări
- 1.imunitatatea Naturala-DobanditaDocument17 pagini1.imunitatatea Naturala-Dobanditalilip6Încă nu există evaluări
- Microorganisme Bacterii VirusiDocument8 paginiMicroorganisme Bacterii VirusiEcaterina BădărăuÎncă nu există evaluări
- Microorganisme 3 Bacterii - Note de CursDocument13 paginiMicroorganisme 3 Bacterii - Note de CursLiluÎncă nu există evaluări
- Curs PMAIA 1-6+LABDocument112 paginiCurs PMAIA 1-6+LABGeoo 99Încă nu există evaluări
- Microbiologie+table Bacterii Flora OralaDocument42 paginiMicrobiologie+table Bacterii Flora OralaAgache AnamariaÎncă nu există evaluări
- Rețete vegetariene din toată lumea. Mâncăruri sănătoase și gustoaseDe la EverandRețete vegetariene din toată lumea. Mâncăruri sănătoase și gustoaseEvaluare: 4 din 5 stele4/5 (8)
- Dieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivDe la EverandDieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivEvaluare: 5 din 5 stele5/5 (1)
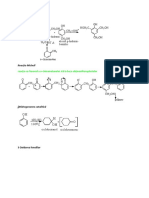
- Fenoli 3Document5 paginiFenoli 3Simo NaaÎncă nu există evaluări
- Dragi MămiciDocument2 paginiDragi MămiciSimo NaaÎncă nu există evaluări
- Curs 19. Familia Enterobacteriaceae. Genul EscherichiaDocument12 paginiCurs 19. Familia Enterobacteriaceae. Genul EscherichiaNicoleta Andreea TudoseÎncă nu există evaluări
- Cand Eram in Floarea MeaDocument1 paginăCand Eram in Floarea MeaSimo NaaÎncă nu există evaluări
- Fenoli 2Document7 paginiFenoli 2Simo NaaÎncă nu există evaluări
- Fenoli 4Document3 paginiFenoli 4Simo NaaÎncă nu există evaluări
- Anto 2Document23 paginiAnto 2Simo NaaÎncă nu există evaluări
- Genul Salmonella PDFDocument3 paginiGenul Salmonella PDFCristina CarstenoiuÎncă nu există evaluări
- Subiectul 5: A. Clasificare GeneralăDocument5 paginiSubiectul 5: A. Clasificare GeneralăSimo NaaÎncă nu există evaluări
- Enterobacterii Lp1 LumiDocument42 paginiEnterobacterii Lp1 LumiElly ElizaÎncă nu există evaluări
- Subiectul 5: A. Clasificare GeneralăDocument5 paginiSubiectul 5: A. Clasificare GeneralăSimo NaaÎncă nu există evaluări
- Piano George BacoviaDocument1 paginăPiano George BacoviaSimo NaaÎncă nu există evaluări
- AntoniaDocument11 paginiAntoniaSimo NaaÎncă nu există evaluări
- La Moartea Principelui Ştirbey de Mihai EminescuDocument1 paginăLa Moartea Principelui Ştirbey de Mihai EminescuNo NameÎncă nu există evaluări
- La Moartea Principelui ŞtirbeyDocument1 paginăLa Moartea Principelui ŞtirbeyBert HardÎncă nu există evaluări
- VirusuriDocument4 paginiVirusurimariaÎncă nu există evaluări
- LunguDocument18 paginiLunguSimo NaaÎncă nu există evaluări
- Gene CancerDocument16 paginiGene CancerSimo NaaÎncă nu există evaluări
- Gene CancerDocument16 paginiGene CancerSimo NaaÎncă nu există evaluări
- LunguDocument18 paginiLunguSimo NaaÎncă nu există evaluări