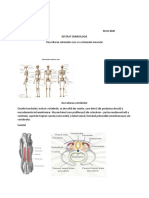
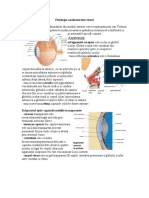
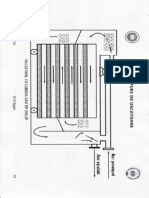
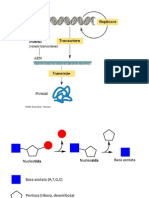
S-ar putea să vă placă și
- Curs Nr. 3 Transporturile TransmembranareDocument56 paginiCurs Nr. 3 Transporturile TransmembranareMarta OstopoviciÎncă nu există evaluări
- Curs Fiziologie An I Medicina - 2012Document71 paginiCurs Fiziologie An I Medicina - 2012Gabriel Ovidiu Avram100% (1)
- Singe FiziologieDocument73 paginiSinge FiziologieAnonymous ck0aGAqZl100% (1)
- Absorbția IntestinalăDocument21 paginiAbsorbția IntestinalăMarius-Sorin CionteaÎncă nu există evaluări
- Citoscheletul MembranarDocument1 paginăCitoscheletul MembranarCristina PopÎncă nu există evaluări
- Tesuturile AnatomieDocument13 paginiTesuturile AnatomieDan marinescuÎncă nu există evaluări
- SangeleDocument3 paginiSangeleely_ely3395Încă nu există evaluări
- Fiziologie - Curs 5 Neuronul + MiocardulDocument8 paginiFiziologie - Curs 5 Neuronul + Miocardulionut_lupul100% (1)
- Biomoleculele Si CeluleleDocument26 paginiBiomoleculele Si CeluleleAlexandra PoenaruÎncă nu există evaluări
- Anatomie Curs 1Document4 paginiAnatomie Curs 1puiutdragutmicut100% (1)
- Radulescu Lavinia EmbriologieDocument7 paginiRadulescu Lavinia EmbriologieGabriel RadulescuÎncă nu există evaluări
- Subiecte Examen BiochimieDocument3 paginiSubiecte Examen BiochimieAlexandra BalanÎncă nu există evaluări
- Curs CitoscheletDocument12 paginiCurs CitoscheletVladDanilaÎncă nu există evaluări
- Rezumat MicroDocument35 paginiRezumat Microremus_stoica23Încă nu există evaluări
- Anatomie Si Fiziologie - AmgDocument8 paginiAnatomie Si Fiziologie - AmgcostinalinaÎncă nu există evaluări
- Cap - 2 Structura Acizilor NucleiciDocument8 paginiCap - 2 Structura Acizilor Nucleicicreatza20Încă nu există evaluări
- Sistem Vascular - 2018Document10 paginiSistem Vascular - 2018rodicaÎncă nu există evaluări
- Jonctiuni Celulare NurseDocument5 paginiJonctiuni Celulare NurseasafteioanaÎncă nu există evaluări
- Curs 3Document30 paginiCurs 3Anamaria PetrutÎncă nu există evaluări
- 2015 - Fiziologia Tubului Digestiv Curs 1 PDFDocument36 pagini2015 - Fiziologia Tubului Digestiv Curs 1 PDFAndreea MxdÎncă nu există evaluări
- Curs 9 PDFDocument12 paginiCurs 9 PDFCosmyn BelciuÎncă nu există evaluări
- Biocel Rezumat PDFDocument71 paginiBiocel Rezumat PDFArina PrisacariuÎncă nu există evaluări
- Tema - ,,celula - Unitate Structural - Funcțională A Organismului UmanDocument12 paginiTema - ,,celula - Unitate Structural - Funcțională A Organismului UmanIon MosnoiÎncă nu există evaluări
- Histologie - Subiecte - Semestrul 2 DavilaDocument3 paginiHistologie - Subiecte - Semestrul 2 DavilaNoNameInfernal50% (2)
- LDH CPK Referat LpmedDocument10 paginiLDH CPK Referat LpmedLaurentiu Olteanu100% (1)
- Lipoproteine PlasmaticeDocument13 paginiLipoproteine PlasmaticeipaperrrÎncă nu există evaluări
- Echilibrul Hidro-Electrolitic CursDocument68 paginiEchilibrul Hidro-Electrolitic CursRazvan MocanuÎncă nu există evaluări
- ATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialDocument8 paginiATP Si ADP - Uzinele Energetice Celulare - Mitocondriile - Invelisul Mitocondrial, Matrixul MitocondrialPascu Alexandru0% (1)
- CELULADocument18 paginiCELULAValentina Budurca100% (1)
- VitamineleDocument16 paginiVitamineleRaduÎncă nu există evaluări
- Sub EmbriologieDocument16 paginiSub EmbriologieSandu Emanuela RamonaÎncă nu există evaluări
- Biofizica I Imagistica MedicalaDocument85 paginiBiofizica I Imagistica MedicalaLivia MiclausÎncă nu există evaluări
- Epiteliu Glandular - ProfesorDocument27 paginiEpiteliu Glandular - ProfesorAdÎncă nu există evaluări
- Curs PancreasDocument17 paginiCurs PancreasZÎncă nu există evaluări
- Histologia Generală Şi HistogenezaDocument457 paginiHistologia Generală Şi HistogenezaRaluca MihalacheÎncă nu există evaluări
- Vitamina PPDocument3 paginiVitamina PPIulian StorojovÎncă nu există evaluări
- Embriologie UmanaDocument20 paginiEmbriologie UmanaCozmescualinÎncă nu există evaluări
- NEUROPSIHOLOGIEDocument3 paginiNEUROPSIHOLOGIEMaria Mădălina100% (1)
- #Micro Sem 1 Subiecte RezolvateDocument34 pagini#Micro Sem 1 Subiecte RezolvateRadu DimulescuÎncă nu există evaluări
- Maduva Spinarii Si Nervii SpinaliDocument9 paginiMaduva Spinarii Si Nervii SpinalinetprintÎncă nu există evaluări
- Aparatul Genital MasculinDocument6 paginiAparatul Genital MasculinAriela SăvuţÎncă nu există evaluări
- Fiziologia Analizatorului VizualDocument2 paginiFiziologia Analizatorului VizualPopa CarmenÎncă nu există evaluări
- LP 10 - Histochimie +Document43 paginiLP 10 - Histochimie +Andreea Maria BanicaÎncă nu există evaluări
- Receptori BiochimieDocument96 paginiReceptori Biochimiedeynyk0% (1)
- AlergiaDocument155 paginiAlergiaElena CataragaÎncă nu există evaluări
- Hormoni LiposolubiliDocument45 paginiHormoni LiposolubiliIchbinSSÎncă nu există evaluări
- Biochimie - Subiecte MetabolismDocument84 paginiBiochimie - Subiecte Metabolismdcn0% (1)
- Digestia Si Absorbtia Principiilor Alimentare PDFDocument8 paginiDigestia Si Absorbtia Principiilor Alimentare PDFAndreea MărcușÎncă nu există evaluări
- Fiziologia NutritieiDocument72 paginiFiziologia NutritieiAndrei BulgariuÎncă nu există evaluări
- METABOLISMULDocument17 paginiMETABOLISMULAndreea StefaniaÎncă nu există evaluări
- Amg - Biochimie, C 3 Metabolismul OkDocument4 paginiAmg - Biochimie, C 3 Metabolismul OkGrupa AMG 1A-2018 SeminarÎncă nu există evaluări
- Digestia Si Absorbtia GlucidelorDocument7 paginiDigestia Si Absorbtia GlucidelorBogdan Bogdan75% (4)
- Fiziologia Sistemului DigestivDocument5 paginiFiziologia Sistemului DigestivlauraÎncă nu există evaluări
- Cursul 5Document16 paginiCursul 5Claudia MădălinaÎncă nu există evaluări
- Absorbtia GlucidelorDocument19 paginiAbsorbtia GlucidelordolaÎncă nu există evaluări
- Cu MetabolismulDocument13 paginiCu MetabolismulAdrian X AttraktiveÎncă nu există evaluări
- C8 Metabolism PROTEINE IPA 20161Document37 paginiC8 Metabolism PROTEINE IPA 20161Bogdan NaeÎncă nu există evaluări
- Biologie DigestiaDocument2 paginiBiologie DigestiaGabriel HerbeiÎncă nu există evaluări
- Stan Ariadna - AMG 1 - Metabolismul Fiintelor VIIDocument11 paginiStan Ariadna - AMG 1 - Metabolismul Fiintelor VIIarrriadna100% (1)
- Institutul de Biochimie Al Academiei Romane Istorie Prezent Si PerspectiveDocument10 paginiInstitutul de Biochimie Al Academiei Romane Istorie Prezent Si PerspectiveMădălina Elena AndrişoaeiÎncă nu există evaluări
- Tipuri de UscatoareDocument5 paginiTipuri de UscatoareMădălina Elena AndrişoaeiÎncă nu există evaluări
- METABOLISMUL LaboratorDocument10 paginiMETABOLISMUL LaboratorMădălina Elena AndrişoaeiÎncă nu există evaluări
- MitocondriaDocument13 paginiMitocondriaCo BbÎncă nu există evaluări
- Ciclul KrebsDocument7 paginiCiclul KrebsAndrei AndreyÎncă nu există evaluări
- Metabolismul Grad IIDocument15 paginiMetabolismul Grad IIMădălina Elena AndrişoaeiÎncă nu există evaluări
- TMDocument66 paginiTMMădălina Elena AndrişoaeiÎncă nu există evaluări
- Acizi Nucleici FDIDocument34 paginiAcizi Nucleici FDIMădălina Elena AndrişoaeiÎncă nu există evaluări
- Mintea Conduce LumeaDocument238 paginiMintea Conduce LumeaSebastian Şandor100% (2)
- MuscularDocument79 paginiMuscularMădălina Elena AndrişoaeiÎncă nu există evaluări
- Reactii RedoxDocument4 paginiReactii RedoxMădălina Elena AndrişoaeiÎncă nu există evaluări
- Reactii RedoxDocument4 paginiReactii RedoxMădălina Elena AndrişoaeiÎncă nu există evaluări