S-ar putea să vă placă și
- Fit Generation: Primul Manual De Fitness Din RomaniaDe la EverandFit Generation: Primul Manual De Fitness Din RomaniaEvaluare: 3.5 din 5 stele3.5/5 (9)
- Superintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateDe la EverandSuperintestinul: Un plan de patru săptămâni pentru reprogramarea microbiomului, refacerea sănătății și pierderea în greutateEvaluare: 5 din 5 stele5/5 (1)
- Dieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivDe la EverandDieta pentru zile senine II: De la durere la echilibru spiritual, psihologic, nutritivEvaluare: 5 din 5 stele5/5 (1)
- Sunetul fizicii: Acustica fenomenologicăDe la EverandSunetul fizicii: Acustica fenomenologicăEvaluare: 4.5 din 5 stele4.5/5 (2)
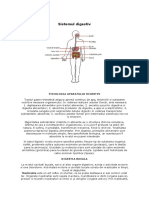
- Fiziologia Aparatului DigestivDocument57 paginiFiziologia Aparatului Digestivrussianangel100% (10)
- Microcirculația sanguină: Următoarea generație de asistență socialăDe la EverandMicrocirculația sanguină: Următoarea generație de asistență socialăÎncă nu există evaluări
- Pancreatita AcutaDocument36 paginiPancreatita Acutamihaella_litovcenco100% (2)
- Anatomia Si Fiziologia PancreasuluiDocument8 paginiAnatomia Si Fiziologia PancreasuluiCristian Varzan67% (3)
- Anatomia Si FiziologiaDocument28 paginiAnatomia Si FiziologiaRaul Hali100% (1)
- Sistemul Nervos EntericDocument22 paginiSistemul Nervos EntericAndrei FrincuÎncă nu există evaluări
- Sistemul Digestiv-RecapitulareDocument17 paginiSistemul Digestiv-RecapitulareElla DeaconuÎncă nu există evaluări
- Calatoria Prin Sistemul DigestivDocument25 paginiCalatoria Prin Sistemul Digestivalina_buÎncă nu există evaluări
- Anatomie PancreasDocument4 paginiAnatomie PancreasAndreeaMihaelaÎncă nu există evaluări
- Digestia in Intestinul Subtire - Curs5Document6 paginiDigestia in Intestinul Subtire - Curs5svlaic2006601Încă nu există evaluări
- Îngrijirea Pacientului Cu Cancer GastricDocument43 paginiÎngrijirea Pacientului Cu Cancer GastricIonela-Georgiana Urse100% (1)
- Functia Motorie A StomaculuiDocument8 paginiFunctia Motorie A Stomaculuisvlaic2006601Încă nu există evaluări
- Sistemul RenalDocument13 paginiSistemul RenalAndries AlexandruÎncă nu există evaluări
- Medicația Aparatului DigestivDocument19 paginiMedicația Aparatului DigestiviulianaÎncă nu există evaluări
- Curs Digestie 1Document38 paginiCurs Digestie 1Ioana Oprea100% (1)
- DigestiaDocument9 paginiDigestiaAlexandru Rusen100% (1)
- Tema 1 DigestivDocument8 paginiTema 1 DigestivMihaela Botnari-GuţuÎncă nu există evaluări
- Ocluzia IntestinalăDocument87 paginiOcluzia IntestinalăgeorgeÎncă nu există evaluări
- Anatomia Si Fiziologia Etajului Inferior Al Tubului DigestivDocument10 paginiAnatomia Si Fiziologia Etajului Inferior Al Tubului DigestivSergiu HelciugÎncă nu există evaluări
- FiziologieDocument27 paginiFiziologieDenisa MariaÎncă nu există evaluări
- Fiziologie Curs 2Document13 paginiFiziologie Curs 2Radu TeodoruÎncă nu există evaluări
- Digestiv Curs 1Document43 paginiDigestiv Curs 1Alina Alexandra ToeaÎncă nu există evaluări
- Referat Intestinul SubtireDocument6 paginiReferat Intestinul SubtireAlex IonescuÎncă nu există evaluări
- Fiziologia IntestinalaDocument76 paginiFiziologia IntestinalaMirela Maria BarÎncă nu există evaluări
- CURSUL I Partea 1Document7 paginiCURSUL I Partea 1Beatrice Sandra Chelaru100% (1)
- Motilitatea EsofagianaDocument6 paginiMotilitatea EsofagianaBianca VaidaÎncă nu există evaluări
- Sistemul DigestivDocument4 paginiSistemul DigestivAlex G15Încă nu există evaluări
- CURS3FIZIODocument18 paginiCURS3FIZIOSorina IoanaÎncă nu există evaluări
- Curs 3 DigestivDocument72 paginiCurs 3 DigestivVladone VladonelolÎncă nu există evaluări
- Fluidele Corpului Si RinichiiDocument53 paginiFluidele Corpului Si RinichiiAnişoara Frunze100% (1)
- 1 Digestiv Generalitati MotilitateDocument10 pagini1 Digestiv Generalitati MotilitateAlexandru TriponÎncă nu există evaluări
- Sistemul DigestivDocument3 paginiSistemul DigestivIon PopÎncă nu există evaluări
- Motilitatea GastrointestinalaDocument42 paginiMotilitatea Gastrointestinalatopea jessicaÎncă nu există evaluări
- Absorbţia În Tubul DigestivDocument18 paginiAbsorbţia În Tubul DigestivNicolaiChistolÎncă nu există evaluări
- Fiziologia Sistemului Digestiv I MDDocument164 paginiFiziologia Sistemului Digestiv I MDClaudiu LefterÎncă nu există evaluări
- Splanhnic, Pancreas, EsofagDocument6 paginiSplanhnic, Pancreas, EsofagEllena CristianaÎncă nu există evaluări
- C9 - 10 Intestin Si Absorbtie 2014Document43 paginiC9 - 10 Intestin Si Absorbtie 2014ZÎncă nu există evaluări
- Sistemul DigestivDocument7 paginiSistemul Digestivsimco09100% (1)
- DIGESTIADocument25 paginiDIGESTIAAlin FreeManÎncă nu există evaluări
- Curs 2 DigestivDocument35 paginiCurs 2 Digestivirenevoicu920Încă nu există evaluări
- Tulburările Motilităţii Şi Secreţiei Tractului DigestivDocument5 paginiTulburările Motilităţii Şi Secreţiei Tractului Digestivnicole_22Încă nu există evaluări
- Procesul Digestiei Şi AbsorbţieiDocument23 paginiProcesul Digestiei Şi AbsorbţieiMaricica RausÎncă nu există evaluări
- CURS 05 - Patologia PancreasuluiDocument22 paginiCURS 05 - Patologia PancreasuluiPătru DoruÎncă nu există evaluări
- Licenţă Crăciun Roxana IoanaDocument15 paginiLicenţă Crăciun Roxana IoanaRoxana IoanaÎncă nu există evaluări
- Rolurile Multiple Ale Rinichilor in HomeostazieDocument2 paginiRolurile Multiple Ale Rinichilor in HomeostazieMarinela MureaÎncă nu există evaluări
- Digestiv Curs 1Document40 paginiDigestiv Curs 1Teodor DiaconuÎncă nu există evaluări
- B UlceroasăDocument29 paginiB UlceroasăLăcrămioara ScripnicÎncă nu există evaluări
- 13.1. Stomac Si DuodenDocument2 pagini13.1. Stomac Si DuodenGruia AlexandraÎncă nu există evaluări
- PANCREASUL Anatomie Si FiziologieDocument3 paginiPANCREASUL Anatomie Si FiziologieDiana CotofreiÎncă nu există evaluări
- Digestia IntestinalăDocument28 paginiDigestia IntestinalăitsmeirinaÎncă nu există evaluări
- Tema 4.4 Fiziologie STOM ROMDocument4 paginiTema 4.4 Fiziologie STOM ROMstudentÎncă nu există evaluări
- Sistemul DigestivDocument8 paginiSistemul Digestivvalentin120Încă nu există evaluări
- Preparatele Cu Acţiune Asupra Inervaţiei Periferice Preparatele Colinomimetice Şi AnticolinesteraziceDocument3 paginiPreparatele Cu Acţiune Asupra Inervaţiei Periferice Preparatele Colinomimetice Şi AnticolinesteraziceFulgdeneaÎncă nu există evaluări
- ANALGEZICE OPIOIDE NEOPIOIDE PT Acad.Document88 paginiANALGEZICE OPIOIDE NEOPIOIDE PT Acad.FulgdeneaÎncă nu există evaluări
- Radiologie Examen Imagini1Document86 paginiRadiologie Examen Imagini1FulgdeneaÎncă nu există evaluări
- Farma Lesin AmusDocument2 paginiFarma Lesin AmusFulgdeneaÎncă nu există evaluări
- F I Z I o P A T o L o G I A Generală StudiazăDocument1 paginăF I Z I o P A T o L o G I A Generală StudiazăFulgdeneaÎncă nu există evaluări
- Patologia Vasculară. Patologia Cordului. Reumatism. - 0Document6 paginiPatologia Vasculară. Patologia Cordului. Reumatism. - 0FulgdeneaÎncă nu există evaluări
- Tot 2 FizpatDocument17 paginiTot 2 FizpatFulgdeneaÎncă nu există evaluări
- AUSCULTATIA PLAMANILOR - STDDocument122 paginiAUSCULTATIA PLAMANILOR - STDFulgdeneaÎncă nu există evaluări
- Ilovepdf MergedDocument44 paginiIlovepdf MergedFulgdeneaÎncă nu există evaluări
- Ar Și Ghini Un 10Document108 paginiAr Și Ghini Un 10FulgdeneaÎncă nu există evaluări
- MG III Rom SD Clinice În Gastrită Boală Ulceroasă Pancreatită-87855Document148 paginiMG III Rom SD Clinice În Gastrită Boală Ulceroasă Pancreatită-87855FulgdeneaÎncă nu există evaluări
- FarmaDocument14 paginiFarmaFulgdeneaÎncă nu există evaluări
- ChirurgieDocument14 paginiChirurgieFulgdeneaÎncă nu există evaluări
- Farma TabeleDocument2 paginiFarma TabeleFulgdenea100% (1)
- Orarul Sem 5 2023-2024Document1 paginăOrarul Sem 5 2023-2024FulgdeneaÎncă nu există evaluări
- Test Examen SIMU FP Speciala 2023Document169 paginiTest Examen SIMU FP Speciala 2023FulgdeneaÎncă nu există evaluări
- Lucrare - de - Control - 2 DUCANTONI DANIELADocument6 paginiLucrare - de - Control - 2 DUCANTONI DANIELAFulgdeneaÎncă nu există evaluări
- Indicatii Metodice Farmacologie Sem V 2022 2023Document70 paginiIndicatii Metodice Farmacologie Sem V 2022 2023FulgdeneaÎncă nu există evaluări
- Caracteristica Medico-Tactică A Focarelor Chimice (Cu Substanțe Toxice) .Document8 paginiCaracteristica Medico-Tactică A Focarelor Chimice (Cu Substanțe Toxice) .FulgdeneaÎncă nu există evaluări
- Răspunsuri La Testarea nr.3 Opțional Genetica 2023 PDFDocument3 paginiRăspunsuri La Testarea nr.3 Opțional Genetica 2023 PDFFulgdeneaÎncă nu există evaluări
- Răspunsuri La Testarea nr.4 Genetica Opțional 2023 PDFDocument4 paginiRăspunsuri La Testarea nr.4 Genetica Opțional 2023 PDFFulgdeneaÎncă nu există evaluări
- Biochimie 1.2Document24 paginiBiochimie 1.2FulgdeneaÎncă nu există evaluări
- Einstein RulitDocument39 paginiEinstein RulitFulgdeneaÎncă nu există evaluări