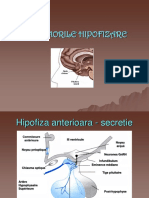
S-ar putea să vă placă și
- Referat HormoniDocument8 paginiReferat HormoniAlin Whiskey80% (5)
- Curs Nr. 9-Biochimie PDFDocument11 paginiCurs Nr. 9-Biochimie PDFAnaMariaMihaitaÎncă nu există evaluări
- HormoniiDocument174 paginiHormoniiMihai DumbravanÎncă nu există evaluări
- Socul NeurogenDocument5 paginiSocul NeurogenAndrei BulgariuÎncă nu există evaluări
- Curs1 Hormon CompletatiDocument57 paginiCurs1 Hormon CompletatiSilvia Nicoleta PredeleanuÎncă nu există evaluări
- Curs 1 EndocrinologieDocument78 paginiCurs 1 EndocrinologiePaula CristinaÎncă nu există evaluări
- Referat Hormoni Si ProteineDocument10 paginiReferat Hormoni Si ProteineEdiÎncă nu există evaluări
- Curs HormoniDocument25 paginiCurs HormoniNicolaÎncă nu există evaluări
- Proiect BiologieDocument12 paginiProiect BiologieAnastasia DarkÎncă nu există evaluări
- Curs 1 - Introducere in EndocrinologieDocument20 paginiCurs 1 - Introducere in EndocrinologieAndra PascuÎncă nu există evaluări
- Manual Endocrinologie - 2022 - Final 24,03,22Document295 paginiManual Endocrinologie - 2022 - Final 24,03,22Lorena ApostolÎncă nu există evaluări
- Referat Hormoni Si ProteineDocument24 paginiReferat Hormoni Si Proteinesilvana andreeaÎncă nu există evaluări
- Carte NutritieDocument108 paginiCarte NutritieOxana Tihon100% (1)
- Efectori Biochimici. HormoniDocument12 paginiEfectori Biochimici. HormoniLivia MariaÎncă nu există evaluări
- Endocrin CumulativDocument508 paginiEndocrin CumulativirrrsÎncă nu există evaluări
- HormoniDocument83 paginiHormoniionut1ilie-1Încă nu există evaluări
- Cursuri EndoDocument729 paginiCursuri EndoLorenaÎncă nu există evaluări
- Biochimie HormonalaDocument82 paginiBiochimie HormonalaMoldovan Nicolae-Andrei100% (3)
- Curs 3 Hipofiza-Generalitati, TumoriDocument33 paginiCurs 3 Hipofiza-Generalitati, TumoriDaniela ToporauÎncă nu există evaluări
- Endocrine Tot TesttDocument371 paginiEndocrine Tot TesttLorenaÎncă nu există evaluări
- Hipofiza-Generalitati, TumoriDocument34 paginiHipofiza-Generalitati, TumoriGeta ContescuÎncă nu există evaluări
- 10.fiziologia Glandelor Endocrine.2022Document102 pagini10.fiziologia Glandelor Endocrine.2022Elena BordeianuÎncă nu există evaluări
- Sistemul EndocrinDocument30 paginiSistemul EndocrinDARIUS IONUŢ MAFTEIÎncă nu există evaluări
- HipofizaDocument36 paginiHipofizaTiToÎncă nu există evaluări
- Curs 4 HormoniDocument29 paginiCurs 4 HormoniStefanCrew GamingÎncă nu există evaluări
- Sistemul EndocrinDocument30 paginiSistemul EndocrinDARIUS IONUŢ MAFTEIÎncă nu există evaluări
- 10.fiziologia Glandelor Endocrine.2018Document99 pagini10.fiziologia Glandelor Endocrine.2018Andrei Avîrvarei100% (1)
- Fiziopatologie Hiper Si HipocorticismDocument12 paginiFiziopatologie Hiper Si HipocorticismRailean CatalinaÎncă nu există evaluări
- HormoniiDocument8 paginiHormoniiSergiu AgostonÎncă nu există evaluări
- Sistemul EndocrinDocument15 paginiSistemul EndocrinMiron EvelinaÎncă nu există evaluări
- Curs Glande1Document50 paginiCurs Glande1Petre GÎncă nu există evaluări
- Curs 1 HormoniDocument20 paginiCurs 1 HormoniDaniela MariaÎncă nu există evaluări
- CURS 7 - Hipotalamus, Hipofiza, TiroidaDocument70 paginiCURS 7 - Hipotalamus, Hipofiza, TiroidaTeodor DiaconuÎncă nu există evaluări
- Clasificarea Hormonilor in Functie de Locul de ProducereDocument2 paginiClasificarea Hormonilor in Functie de Locul de ProducereVlaicu Dan MariusÎncă nu există evaluări
- 13 Ro Sistemul Endocrin 1Document94 pagini13 Ro Sistemul Endocrin 1Csabesz BenczoÎncă nu există evaluări
- Glandele Endocrine Si AP GenitalDocument79 paginiGlandele Endocrine Si AP GenitalAlinaÎncă nu există evaluări
- Hormonii AntehipofizariDocument5 paginiHormonii AntehipofizariAndrei BulgariuÎncă nu există evaluări
- Material AMGDocument55 paginiMaterial AMGMihaela Elena Purecel100% (1)
- Preparatele Hormonale Şi Antagoniştii LorDocument43 paginiPreparatele Hormonale Şi Antagoniştii LorOleg BalanÎncă nu există evaluări
- HIPOFIZADocument15 paginiHIPOFIZAAdela MoldovanÎncă nu există evaluări
- Sistemul EndocrinDocument112 paginiSistemul EndocrinIrina Fratila100% (1)
- Hormonii BiochimieDocument303 paginiHormonii BiochimieIngaŞaragovÎncă nu există evaluări
- Endo Rom 2016Document63 paginiEndo Rom 2016Sergiu MalinÎncă nu există evaluări
- Curs2r HormoniDocument55 paginiCurs2r Hormonichintesti100% (1)
- HormoniiDocument16 paginiHormoniielenaÎncă nu există evaluări
- Hormoni 2 2014Document71 paginiHormoni 2 2014Ade Adde100% (3)
- Hormonii Si Rolul Lor in Organismul Uman 1Document53 paginiHormonii Si Rolul Lor in Organismul Uman 1elena rusuÎncă nu există evaluări
- HORMONII BiochimieDocument4 paginiHORMONII BiochimieSorin Turcu100% (1)
- 12.TM Curs7Document46 pagini12.TM Curs7Ana BeneÎncă nu există evaluări
- Ginecologie VeterinaraDocument505 paginiGinecologie Veterinaraemutza7992% (12)
- Mielomul MultipluDocument9 paginiMielomul MultipluAndrei BulgariuÎncă nu există evaluări
- Biologia TumoralaDocument18 paginiBiologia TumoralaAndrei BulgariuÎncă nu există evaluări
- Infectii NeonataleDocument82 paginiInfectii NeonataleAndrei BulgariuÎncă nu există evaluări
- Alimentatia SugarilorDocument5 paginiAlimentatia SugarilorAndrei BulgariuÎncă nu există evaluări
- Icterele NeonataleDocument16 paginiIcterele NeonataleAndrei BulgariuÎncă nu există evaluări
- Tulburari de NutritieDocument23 paginiTulburari de NutritieAndrei BulgariuÎncă nu există evaluări
- Comportamentul AlimentarDocument23 paginiComportamentul AlimentarAndrei Bulgariu100% (1)
- Fiziologia NutritieiDocument72 paginiFiziologia NutritieiAndrei BulgariuÎncă nu există evaluări
- Genetica Bolilor MetaboliceDocument13 paginiGenetica Bolilor MetaboliceAndrei BulgariuÎncă nu există evaluări
- Bazele TeoreticeDocument242 paginiBazele TeoreticeAndrei BulgariuÎncă nu există evaluări
- Patologia Peretelui AbdominalDocument26 paginiPatologia Peretelui AbdominalAndrei BulgariuÎncă nu există evaluări
- Metode de InvestigatieDocument49 paginiMetode de InvestigatieAndrei BulgariuÎncă nu există evaluări
- Imagistica Cervico-ToracicaDocument44 paginiImagistica Cervico-ToracicaAndrei BulgariuÎncă nu există evaluări
- Socul HipovolemicDocument6 paginiSocul HipovolemicAndrei BulgariuÎncă nu există evaluări
- Socul CardiogenDocument13 paginiSocul CardiogenAndrei BulgariuÎncă nu există evaluări
- Socul AnafilacticDocument3 paginiSocul AnafilacticAndrei BulgariuÎncă nu există evaluări