S-ar putea să vă placă și
- Biomembrane Leabu PDFDocument132 paginiBiomembrane Leabu PDFConstantin VasyÎncă nu există evaluări
- 2008 Med Octavian DoagaDocument24 pagini2008 Med Octavian DoagaVasile EuÎncă nu există evaluări
- !!!!!!233modelarea Celulara in SilicoDocument12 pagini!!!!!!233modelarea Celulara in SilicoPetru AcozmeiÎncă nu există evaluări
- Biocel Curs + LPDocument386 paginiBiocel Curs + LPAna-Maria MititeluÎncă nu există evaluări
- Biologie CelularaDocument18 paginiBiologie CelularaRares IonutÎncă nu există evaluări
- MRM Rezistenta MatDocument385 paginiMRM Rezistenta MatZZÎncă nu există evaluări
- Urmarirea Fibrelor Principii Si StrategiiDocument4 paginiUrmarirea Fibrelor Principii Si StrategiiMady BuzeaÎncă nu există evaluări
- C1 BiofizicaDocument26 paginiC1 BiofizicaAdoptedchildÎncă nu există evaluări
- BIOFIZICADocument90 paginiBIOFIZICAy0nutz321100% (7)
- BiomecanicaDocument13 paginiBiomecanicaAntoanellaÎncă nu există evaluări
- Organe BioartificialeDocument11 paginiOrgane BioartificialeDana LebadaÎncă nu există evaluări
- Celula - MembranaDocument11 paginiCelula - MembranaAlexandra ViolettÎncă nu există evaluări
- Biomimetica Si Regenerare TisularaDocument7 paginiBiomimetica Si Regenerare TisularaAisar FaiadÎncă nu există evaluări
- Modelul Mozaicului Fluid Al Membranei CelulareDocument21 paginiModelul Mozaicului Fluid Al Membranei CelulareZgherea MihaiÎncă nu există evaluări
- Curs CitologieDocument7 paginiCurs CitologieCojocariu Hanganu DenisiaÎncă nu există evaluări
- Lista SubiecteDocument116 paginiLista SubiecteYazan ShawishÎncă nu există evaluări
- Modele - Nano - PpsDocument67 paginiModele - Nano - PpsDenisa Neagu100% (1)
- Citologie VegetalaDocument70 paginiCitologie VegetalaGabriela TanasescuÎncă nu există evaluări
- Introducere in Cristale Lichide Cristina Cirtoaje 2010Document6 paginiIntroducere in Cristale Lichide Cristina Cirtoaje 2010cristina_cirtoajeÎncă nu există evaluări
- 2020-2021 Membrana CelularaDocument5 pagini2020-2021 Membrana CelularaRéd Ðrågøș100% (1)
- Notiuni Introductive IDocument2 paginiNotiuni Introductive IElena Corina MoiseÎncă nu există evaluări
- Cap. Iii. Modele Matematice Care Descriu Funcționarea Bioreactoarelor Proceselor În BioreactoareDocument25 paginiCap. Iii. Modele Matematice Care Descriu Funcționarea Bioreactoarelor Proceselor În BioreactoareCatalin ScriosteanuÎncă nu există evaluări
- Invelisul CelularDocument7 paginiInvelisul CelularDumitrita100% (1)
- Curs Biologie Celulara IFR PDFDocument76 paginiCurs Biologie Celulara IFR PDFIvan MihaelaÎncă nu există evaluări
- Lisii Corneliu Teza Licenta PDFDocument39 paginiLisii Corneliu Teza Licenta PDFPatricia HeraÎncă nu există evaluări
- Modele Celulare StructurateDocument38 paginiModele Celulare StructurateGabriel GabiÎncă nu există evaluări
- .Lecția 1Document23 pagini.Lecția 1Alina VoicoÎncă nu există evaluări
- Definiţia Şi Funcţiile Membranelor BiologiceDocument23 paginiDefiniţia Şi Funcţiile Membranelor BiologiceDanila ElenaÎncă nu există evaluări
- Curs Biofizica 2Document4 paginiCurs Biofizica 2Mariana DimaÎncă nu există evaluări
- Proprietăți Mecanice de Os Cortical Și Țesut Osos CancerosDocument10 paginiProprietăți Mecanice de Os Cortical Și Țesut Osos CancerosRomînu VladÎncă nu există evaluări
- Biotehnologie-Pro Sau ContraDocument5 paginiBiotehnologie-Pro Sau ContrasupravietuireÎncă nu există evaluări
- Citologie FRDocument50 paginiCitologie FRdenesgt500100% (1)
- Electro SpinningDocument4 paginiElectro SpinningMaria CristinaÎncă nu există evaluări
- Oana Niculescu Teza DoctoratDocument246 paginiOana Niculescu Teza DoctoratMarius RosuÎncă nu există evaluări
- Punctele CuanticeDocument16 paginiPunctele Cuanticeelish3baÎncă nu există evaluări
- 1 Tema AMG-47896-54608Document19 pagini1 Tema AMG-47896-54608Kuharskaia ElenaÎncă nu există evaluări
- BCM C1 2022Document6 paginiBCM C1 2022Matei NegoitaÎncă nu există evaluări
- Biomateriale Pentru Ingineria TisularaDocument19 paginiBiomateriale Pentru Ingineria TisularaAlexandra IoanaÎncă nu există evaluări
- Mecanobiologia CeluleiDocument30 paginiMecanobiologia CeluleiBianca Ionela DraganÎncă nu există evaluări
- Curs 1 Biologie Celulara Si Moleculara 2020Document11 paginiCurs 1 Biologie Celulara Si Moleculara 2020barney balalauÎncă nu există evaluări
- 9C - Celula, nr.1 Si 2Document2 pagini9C - Celula, nr.1 Si 2Oana DobreÎncă nu există evaluări
- BCM C5 2022Document4 paginiBCM C5 2022Matei NegoitaÎncă nu există evaluări
- ExcitabilitateaDocument27 paginiExcitabilitateastrengara100% (2)
- Excitabilitatea PDFDocument27 paginiExcitabilitatea PDFAndrei Nicolae BejanÎncă nu există evaluări
- Originea Și Evoluția Celulei Eucariote, Curs, Evoluționism, 6.04.2015Document15 paginiOriginea Și Evoluția Celulei Eucariote, Curs, Evoluționism, 6.04.2015Cristina Cris100% (1)
- Modelare MolecularaDocument4 paginiModelare MolecularaantinieaÎncă nu există evaluări
- Nanotehnologie in ElectronicaDocument10 paginiNanotehnologie in ElectronicaC0colinoÎncă nu există evaluări
- Biologie Celulara - Lucrari Practice PDFDocument85 paginiBiologie Celulara - Lucrari Practice PDFcherim ailin80% (5)
- Carte LP 2007 Biologie Celulara-1Document85 paginiCarte LP 2007 Biologie Celulara-1Alina Madalina50% (2)
- Ecuatia lui Dumnezeu: In cautarea unei teorii a tuturor lucrurilorDe la EverandEcuatia lui Dumnezeu: In cautarea unei teorii a tuturor lucrurilorÎncă nu există evaluări
- Epistemologia gravitației experimentale: Raționalitatea științificăDe la EverandEpistemologia gravitației experimentale: Raționalitatea științificăÎncă nu există evaluări
- Cunoașterea Științifică, Volumul 2, Numărul 3, Septembrie 2023De la EverandCunoașterea Științifică, Volumul 2, Numărul 3, Septembrie 2023Încă nu există evaluări
- Grumezescu Florina - Tehnici Computerizate de Proiectare A Biomaterialelor Complexe PDFDocument5 paginiGrumezescu Florina - Tehnici Computerizate de Proiectare A Biomaterialelor Complexe PDFFlorina GrumezescuÎncă nu există evaluări
- Aplicaţii Ale Spectroscopiei Ir Şi Raman În Studiul MaterialelorDocument42 paginiAplicaţii Ale Spectroscopiei Ir Şi Raman În Studiul MaterialelorMarius IonutÎncă nu există evaluări
- Mecanobiologia CeluleiDocument30 paginiMecanobiologia CeluleiBianca Ionela DraganÎncă nu există evaluări
- Utilizarea Celulelor Stem in Ingineria TisularaDocument13 paginiUtilizarea Celulelor Stem in Ingineria TisularaFlorina GrumezescuÎncă nu există evaluări
- Biochimie Generala CursDocument118 paginiBiochimie Generala CursSimona BozaÎncă nu există evaluări
- Utilizarea Celulelor Stem in Ingineria TisularaDocument13 paginiUtilizarea Celulelor Stem in Ingineria TisularaFlorina GrumezescuÎncă nu există evaluări
- Aplicaţii Ale Spectroscopiei Ir Şi Raman În Studiul MaterialelorDocument42 paginiAplicaţii Ale Spectroscopiei Ir Şi Raman În Studiul MaterialelorMarius IonutÎncă nu există evaluări
- Tehnici de Cautare Si Verificare Pe Internet Despre PatenteDocument13 paginiTehnici de Cautare Si Verificare Pe Internet Despre PatenteFlorina GrumezescuÎncă nu există evaluări
- SalciaDocument2 paginiSalciaFlorina GrumezescuÎncă nu există evaluări
- Mecanobiologia CeluleiDocument30 paginiMecanobiologia CeluleiBianca Ionela DraganÎncă nu există evaluări
- CC4 - Relatie celula-MEC PDFDocument24 paginiCC4 - Relatie celula-MEC PDFFlorina GrumezescuÎncă nu există evaluări
- CC3 - Organite Celulare PDFDocument25 paginiCC3 - Organite Celulare PDFFlorina GrumezescuÎncă nu există evaluări
- CC1 - Introducere Si Biologie CelularaDocument23 paginiCC1 - Introducere Si Biologie CelularaAndrei E. Haway100% (1)
- Biotehnologii MedicaleDocument111 paginiBiotehnologii MedicaleFlorina Grumezescu100% (4)
- Curs Biologie MolecularaDocument243 paginiCurs Biologie MolecularaMarin Rabei100% (1)
- Proteza de GleznaDocument25 paginiProteza de GleznaFlorina GrumezescuÎncă nu există evaluări
- Proteza de GleznaDocument25 paginiProteza de GleznaFlorina GrumezescuÎncă nu există evaluări
- Lab 1 - Reologie - 2017-2018Document5 paginiLab 1 - Reologie - 2017-2018Andreea CasadjicovÎncă nu există evaluări
- Tehnici de Cautare Si Verificare Pe Internet Despre PatenteDocument13 paginiTehnici de Cautare Si Verificare Pe Internet Despre PatenteFlorina GrumezescuÎncă nu există evaluări
- CC6 - Caracterizarea Celulelor in CulturaDocument18 paginiCC6 - Caracterizarea Celulelor in CulturaFlorina GrumezescuÎncă nu există evaluări
- Scolioza Lombara Dextroconvexa Cu Rotatie de AxDocument1 paginăScolioza Lombara Dextroconvexa Cu Rotatie de AxFlorina GrumezescuÎncă nu există evaluări
- Tehnici de Cautare Si Verificare Pe Internet Despre PatenteDocument13 paginiTehnici de Cautare Si Verificare Pe Internet Despre PatenteFlorina GrumezescuÎncă nu există evaluări
- SalciaDocument2 paginiSalciaFlorina GrumezescuÎncă nu există evaluări
- BIM 4 Curs Biotehnologii MedicaleDocument30 paginiBIM 4 Curs Biotehnologii MedicaleFlorina GrumezescuÎncă nu există evaluări
- Vascozitatea SangeluiDocument21 paginiVascozitatea SangeluiFlorina Grumezescu100% (2)
- Curs Biologie MolecularaDocument243 paginiCurs Biologie MolecularaMarin Rabei100% (1)
- Asigurarea Calitatii Bioproduselor Prin GMP Si GLPDocument31 paginiAsigurarea Calitatii Bioproduselor Prin GMP Si GLPFlorina Grumezescu100% (1)
- Specificatii MatlabDocument1 paginăSpecificatii MatlabFlorina GrumezescuÎncă nu există evaluări
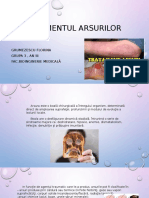
- Tratamentul ArsurilorDocument13 paginiTratamentul ArsurilorFlorina GrumezescuÎncă nu există evaluări