S-ar putea să vă placă și
- Curs Fiziologie An I Medicina - Secretia SalivaraDocument9 paginiCurs Fiziologie An I Medicina - Secretia SalivaraAndreea VladanÎncă nu există evaluări
- Biochimie CTD Suport CursDocument118 paginiBiochimie CTD Suport CursCB100% (1)
- Curs 1Document28 paginiCurs 1Marina PirnauÎncă nu există evaluări
- 1 72Document61 pagini1 72Misha M AndronachiÎncă nu există evaluări
- Sanatatea Orala 2014 PliantDocument2 paginiSanatatea Orala 2014 PliantClaudia Luiza BălanÎncă nu există evaluări
- Pulpa DentaraDocument35 paginiPulpa DentaraTraista EudochiaÎncă nu există evaluări
- Totalizarea 1 Microbiologie OralDocument65 paginiTotalizarea 1 Microbiologie OralMădălina SoltanÎncă nu există evaluări
- Organitele CelulareDocument5 paginiOrganitele CelulareAlexÎncă nu există evaluări
- 4 2 ParodontiulDocument34 pagini4 2 ParodontiulAdela GabrielaÎncă nu există evaluări
- Profilaxia Prenatală.Document3 paginiProfilaxia Prenatală.Valeria CernegaÎncă nu există evaluări
- Endodontie Clinica II Anul IV Rom - 0Document28 paginiEndodontie Clinica II Anul IV Rom - 0FeliciaÎncă nu există evaluări
- Intrebari Pentru Examenul de La Stomatologie Ortopedica USMFDocument5 paginiIntrebari Pentru Examenul de La Stomatologie Ortopedica USMFDumitru Muntean0% (1)
- Curs 6 Biochimie Enzime 3Document25 paginiCurs 6 Biochimie Enzime 3Black Skayokar100% (1)
- 5) Structura ProteinelorDocument44 pagini5) Structura ProteinelorgolripÎncă nu există evaluări
- Curs Optional ParoDocument69 paginiCurs Optional ParocsmalxÎncă nu există evaluări
- MSS nr.16Document3 paginiMSS nr.16ValeriaOjogÎncă nu există evaluări
- LP 3 - Genetică MedicalăDocument26 paginiLP 3 - Genetică MedicalăDariana RahaianÎncă nu există evaluări
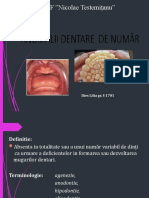
- Anomalii Dentare de NumărDocument43 paginiAnomalii Dentare de NumărLilia LiliaÎncă nu există evaluări
- BIOCHIMIA PROCESULUI DE FORMARE A DIN+óILORDocument22 paginiBIOCHIMIA PROCESULUI DE FORMARE A DIN+óILORteodora100% (1)
- Tema 23Document12 paginiTema 23Ceresău MarinaÎncă nu există evaluări
- Curs 2 MD Peptide, ProteineDocument12 paginiCurs 2 MD Peptide, ProteineDaniela MariaÎncă nu există evaluări
- Curs 13Document3 paginiCurs 13Dunca Ana-MariaÎncă nu există evaluări
- Indicatia Metodica Nr12Document2 paginiIndicatia Metodica Nr12ainura antonÎncă nu există evaluări
- Metabolismul GliceroluluiDocument21 paginiMetabolismul GliceroluluiCalin PopaÎncă nu există evaluări
- Es Dentare-29764Document93 paginiEs Dentare-29764kateÎncă nu există evaluări
- Anomalii Dentare de NumărDocument30 paginiAnomalii Dentare de NumărCamelia PetcuÎncă nu există evaluări
- Cap IIIDocument22 paginiCap IIICezara PanainteÎncă nu există evaluări
- Acizii Nucleici-6084Document157 paginiAcizii Nucleici-6084anabellevanhelsingÎncă nu există evaluări
- Leziuni-ParodontaleDocument14 paginiLeziuni-ParodontaleMl BeÎncă nu există evaluări
- Fiziologie 2Document3 paginiFiziologie 2studentÎncă nu există evaluări
- Micro Prote ZeDocument19 paginiMicro Prote ZeAlexandra MagaleasÎncă nu există evaluări
- Totalizarea 2. Răspuns La Intrebările MetodiceDocument3 paginiTotalizarea 2. Răspuns La Intrebările MetodicestudentÎncă nu există evaluări
- Elaborarea Metodică N 4Document10 paginiElaborarea Metodică N 4Anna BalanÎncă nu există evaluări
- Rolul Tartrului in Producerea Bolii ParodontaleDocument7 paginiRolul Tartrului in Producerea Bolii ParodontaleVintiloiu CorinaÎncă nu există evaluări
- PrezentareDocument16 paginiPrezentaredorinstoÎncă nu există evaluări
- Tartrul DentarDocument7 paginiTartrul DentarAlexandra Robu100% (1)
- Subiecte Biofizica Rezolvate 2012Document18 paginiSubiecte Biofizica Rezolvate 2012gaby90009Încă nu există evaluări
- Tot 1Document22 paginiTot 1Catalina FiodorovÎncă nu există evaluări
- Hipoplazia Smaltului Dentar Si A Cimentului2Document17 paginiHipoplazia Smaltului Dentar Si A Cimentului2Adrian NourÎncă nu există evaluări
- Raspuns Subiecte Examen de Stat Terapie 2018Document171 paginiRaspuns Subiecte Examen de Stat Terapie 2018Cristian BelousÎncă nu există evaluări
- Fiziologia ExcretieiDocument51 paginiFiziologia ExcretieiMedBook100% (2)
- Cromatina SexualăDocument11 paginiCromatina SexualăЮлиан МатковскийÎncă nu există evaluări
- Elaborarea Metodica nr12Document6 paginiElaborarea Metodica nr12Stefan GrigorisÎncă nu există evaluări
- Planul Seminarelor La Bioet Stom 2020-2021-18381Document4 paginiPlanul Seminarelor La Bioet Stom 2020-2021-18381studentÎncă nu există evaluări
- E.M. N 13Document4 paginiE.M. N 13Mihaela CaruntuÎncă nu există evaluări
- Elaborarea Metodica Nr. 1Document5 paginiElaborarea Metodica Nr. 1lozovanudÎncă nu există evaluări
- Teste Ro Examen Stud-1290Document111 paginiTeste Ro Examen Stud-1290Iulian StorojovÎncă nu există evaluări
- Elaborarea Metodica NR 1 OrtoDocument2 paginiElaborarea Metodica NR 1 OrtoValeria CernegaÎncă nu există evaluări
- Elaborarea Metodica nr13Document6 paginiElaborarea Metodica nr13Ana Maria BulatÎncă nu există evaluări
- Tema 22Document6 paginiTema 22Ceresău MarinaÎncă nu există evaluări
- Parodontologie T1-4Document17 paginiParodontologie T1-4Andrei GrumezaÎncă nu există evaluări
- Cazuri Clinice: Consideratii Clinice Privind Restauratiile Protetice Ceramice - Cazuri CliniceDocument8 paginiCazuri Clinice: Consideratii Clinice Privind Restauratiile Protetice Ceramice - Cazuri CliniceFlorina MoraruÎncă nu există evaluări
- Proteze ExDocument93 paginiProteze ExОля СтояноваÎncă nu există evaluări
- Elaborarea Metodica nr13Document8 paginiElaborarea Metodica nr13Михаела ЦурканÎncă nu există evaluări
- Analizatorul VizualDocument4 paginiAnalizatorul VizualCrainic CosminÎncă nu există evaluări
- PAODocument20 paginiPAODaniel CiubatiiÎncă nu există evaluări
- Curs 3 Biochimie - Proteine 3Document33 paginiCurs 3 Biochimie - Proteine 3TGÎncă nu există evaluări
- Curs 1 Biochimie - Proteine 1Document42 paginiCurs 1 Biochimie - Proteine 1Elena CalapodÎncă nu există evaluări
- 2 Structura Proteinelor-67467Document55 pagini2 Structura Proteinelor-67467Anastasia OzarenskiÎncă nu există evaluări
- Curs 1 Biochimie - Proteine 1 PDFDocument42 paginiCurs 1 Biochimie - Proteine 1 PDFCarp TheodorÎncă nu există evaluări
- UntitledDocument37 paginiUntitledBlack SkayokarÎncă nu există evaluări
- Exercitii Caractere Monogenice Normale 20 Noiembrie 2020 LB FinalDocument38 paginiExercitii Caractere Monogenice Normale 20 Noiembrie 2020 LB FinalBlack SkayokarÎncă nu există evaluări
- Exercitii Boli Monogenice 4 Ianuarie 2021 GR 27 de TrimisDocument4 paginiExercitii Boli Monogenice 4 Ianuarie 2021 GR 27 de TrimisBlack SkayokarÎncă nu există evaluări
- Curs BiofizicaDocument72 paginiCurs BiofizicaBlack SkayokarÎncă nu există evaluări
- Curs BF 6 CentrifDocument17 paginiCurs BF 6 CentrifBlack SkayokarÎncă nu există evaluări