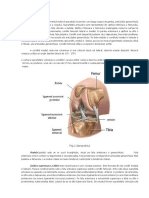
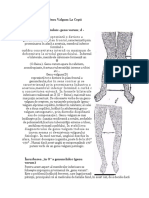
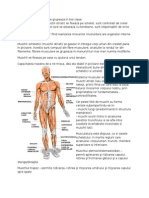
S-ar putea să vă placă și
- Metabolismul HemoglobineiDocument6 paginiMetabolismul HemoglobineiPauhgf MuloÎncă nu există evaluări
- Starile de SocDocument16 paginiStarile de SocAlina Vitali TabarceaÎncă nu există evaluări
- Maladia ScheuermannDocument3 paginiMaladia ScheuermannminichoixÎncă nu există evaluări
- BiomecanicaDocument13 paginiBiomecanicaВлад МортеанÎncă nu există evaluări
- Notiuni de Anatomie Si Fiziologie A Sistemului NervosDocument15 paginiNotiuni de Anatomie Si Fiziologie A Sistemului NervosClaudia-Alexandra BocanetÎncă nu există evaluări
- Curs VI - HidroterapiaDocument30 paginiCurs VI - HidroterapiaMihaela NițulescuÎncă nu există evaluări
- Fractura de Platou Tibial Și Cap PeroneuDocument7 paginiFractura de Platou Tibial Și Cap PeroneuPușcașu CătălinaÎncă nu există evaluări
- Boala ChusingDocument4 paginiBoala ChusingSergiu AndreiÎncă nu există evaluări
- Metabolismul IntermediarDocument3 paginiMetabolismul IntermediarOana KoppandiÎncă nu există evaluări
- Aparatul LocomotorDocument12 paginiAparatul LocomotorCarmen DraghiaÎncă nu există evaluări
- Modificarile Organismului Matern in SarcinaDocument8 paginiModificarile Organismului Matern in Sarcinaserban marianaÎncă nu există evaluări
- Boli Rare in LumeDocument15 paginiBoli Rare in LumeMaria IosefinaÎncă nu există evaluări
- Elemente Privind Fiziopatologia Şi Semiologia Aparatului RespiratorDocument12 paginiElemente Privind Fiziopatologia Şi Semiologia Aparatului RespiratorIrina BordeianuÎncă nu există evaluări
- ANATOMIA APARATULUI DIGESTIV Prima VariantaDocument13 paginiANATOMIA APARATULUI DIGESTIV Prima Variantayoanna216Încă nu există evaluări
- ComaDocument11 paginiComaEstera PascaruÎncă nu există evaluări
- Boli ProfesionaleDocument7 paginiBoli ProfesionaleCat RoseÎncă nu există evaluări
- Emfizemul PulmonarDocument19 paginiEmfizemul PulmonarCelerina AikoÎncă nu există evaluări
- Licenta Gasita Pe Net-Paralizia Cerebrala Lucrare-LicentaDocument70 paginiLicenta Gasita Pe Net-Paralizia Cerebrala Lucrare-LicentaBordeianu RobertÎncă nu există evaluări
- Referat HidroterapieDocument4 paginiReferat HidroterapieSclipcea DaniÎncă nu există evaluări
- Dezvoltarea MembrelorDocument1 paginăDezvoltarea MembrelorIulia IulianaÎncă nu există evaluări
- Tesuturile ConjunctiveDocument17 paginiTesuturile ConjunctiveAlberto1974Încă nu există evaluări
- 18 Steinhardt, Modernism, OSP (Tema)Document3 pagini18 Steinhardt, Modernism, OSP (Tema)Lucian CosneanuÎncă nu există evaluări
- Genu Varum Si Genu Valgum LaDocument3 paginiGenu Varum Si Genu Valgum LaPetre OanaÎncă nu există evaluări
- Avc DorinaDocument13 paginiAvc DorinaDorina PetrovÎncă nu există evaluări
- Paralizia Obstetricala de Plex BrahialDocument14 paginiParalizia Obstetricala de Plex BrahialTatiana DecusearăÎncă nu există evaluări
- GonartrozaDocument2 paginiGonartrozaStefan CincaÎncă nu există evaluări
- Primul Ajutor În Caz de ÎnecDocument6 paginiPrimul Ajutor În Caz de ÎnecAndreea AndrieșÎncă nu există evaluări
- Analizatorul Auditiv VODocument8 paginiAnalizatorul Auditiv VOrefsumÎncă nu există evaluări
- C3 Furuncul. HidrosadenitaDocument22 paginiC3 Furuncul. HidrosadenitaAna-Maria CirnuÎncă nu există evaluări
- AnamnezaDocument6 paginiAnamnezaMarius AlexandrescuÎncă nu există evaluări
- LUXATII ReferatDocument3 paginiLUXATII ReferatAyoub EmiliaÎncă nu există evaluări
- Muschii Corpului Uman Se Grupeaza in Trei ClaseDocument3 paginiMuschii Corpului Uman Se Grupeaza in Trei ClaseGhinea Georgiana ClaudiaÎncă nu există evaluări
- Cv+a PopaDocument3 paginiCv+a PopatuturiÎncă nu există evaluări
- Spondilita AnchilozantaDocument6 paginiSpondilita AnchilozantaSimina MihaiÎncă nu există evaluări
- Portofoliu IgienăDocument29 paginiPortofoliu IgienăMiruna Pana100% (1)
- Ce Este TendinitaDocument5 paginiCe Este TendinitaTurturean ZoiaÎncă nu există evaluări
- Vitamina B1Document5 paginiVitamina B1Christopher BoydÎncă nu există evaluări
- Semiologia IntestinuluiDocument11 paginiSemiologia IntestinuluiMischelle ParvulescuÎncă nu există evaluări
- Faringele Si EsofagulDocument17 paginiFaringele Si EsofagulOana Diana100% (1)
- Boala AddisonDocument40 paginiBoala AddisonMorogai ValeriaÎncă nu există evaluări
- Mijloace Fundamentale - Exercitiu FizicDocument7 paginiMijloace Fundamentale - Exercitiu Fizicbooks.love1995.reading.Încă nu există evaluări
- Margutoiu Alexandru - Sistemul Iritativ VegetativDocument4 paginiMargutoiu Alexandru - Sistemul Iritativ VegetativMargutoiu AlexandruÎncă nu există evaluări
- S03a. FRACTURILE SCAPULEI PDFDocument19 paginiS03a. FRACTURILE SCAPULEI PDFAndrei ScripcaruÎncă nu există evaluări
- 15 Febra TifoidaDocument17 pagini15 Febra TifoidaCtin GeoÎncă nu există evaluări
- Giongiu Valeriu AdrianDocument13 paginiGiongiu Valeriu AdrianiancuÎncă nu există evaluări
- ArticulatiileDocument6 paginiArticulatiileAdriana Dragomir100% (1)
- Proiect Infintare GradinitaDocument42 paginiProiect Infintare GradinitaLuminița CrețuÎncă nu există evaluări
- Semiologia Aparatului DigestivDocument22 paginiSemiologia Aparatului DigestivElena TudoseÎncă nu există evaluări
- Facultatea de Educatie Fizica Si SportDocument4 paginiFacultatea de Educatie Fizica Si SportMitricof BiancaÎncă nu există evaluări
- Traumatisme OsteoDocument10 paginiTraumatisme OsteoGabriela Ximena CaraivanÎncă nu există evaluări
- CURS12-Semiologia Sistemelor Osos Si MuscularDocument107 paginiCURS12-Semiologia Sistemelor Osos Si MuscularCraciunica Dragos-AlexandruÎncă nu există evaluări
- Curs Pediatrie - Pneumonia La Copil 2023Document35 paginiCurs Pediatrie - Pneumonia La Copil 2023nelida luciana egriÎncă nu există evaluări
- Recuperarea in EntorseDocument7 paginiRecuperarea in EntorseFlorin Caprita100% (1)
- TerapiaOcupationala 1Document13 paginiTerapiaOcupationala 1Vica IgnatÎncă nu există evaluări
- Sistemul DigestivDocument3 paginiSistemul DigestivAlexandra HossuÎncă nu există evaluări
- OsteoporozaDocument25 paginiOsteoporozaMadalina CO100% (1)
- Semiologie Budurea VladDocument6 paginiSemiologie Budurea VladVlad BudureaÎncă nu există evaluări
- HemoglobinaDocument8 paginiHemoglobinaxxnetÎncă nu există evaluări
- HemoglobinaDocument4 paginiHemoglobinagelubot100% (3)
- HemoglobinaDocument4 paginiHemoglobinagelubot100% (1)